
Rust (Uromyces viciae-fabae Pers. de-Bary) of Pea (Pisum sativum L.): Present Status and Future Resistance Breeding Opportunities

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
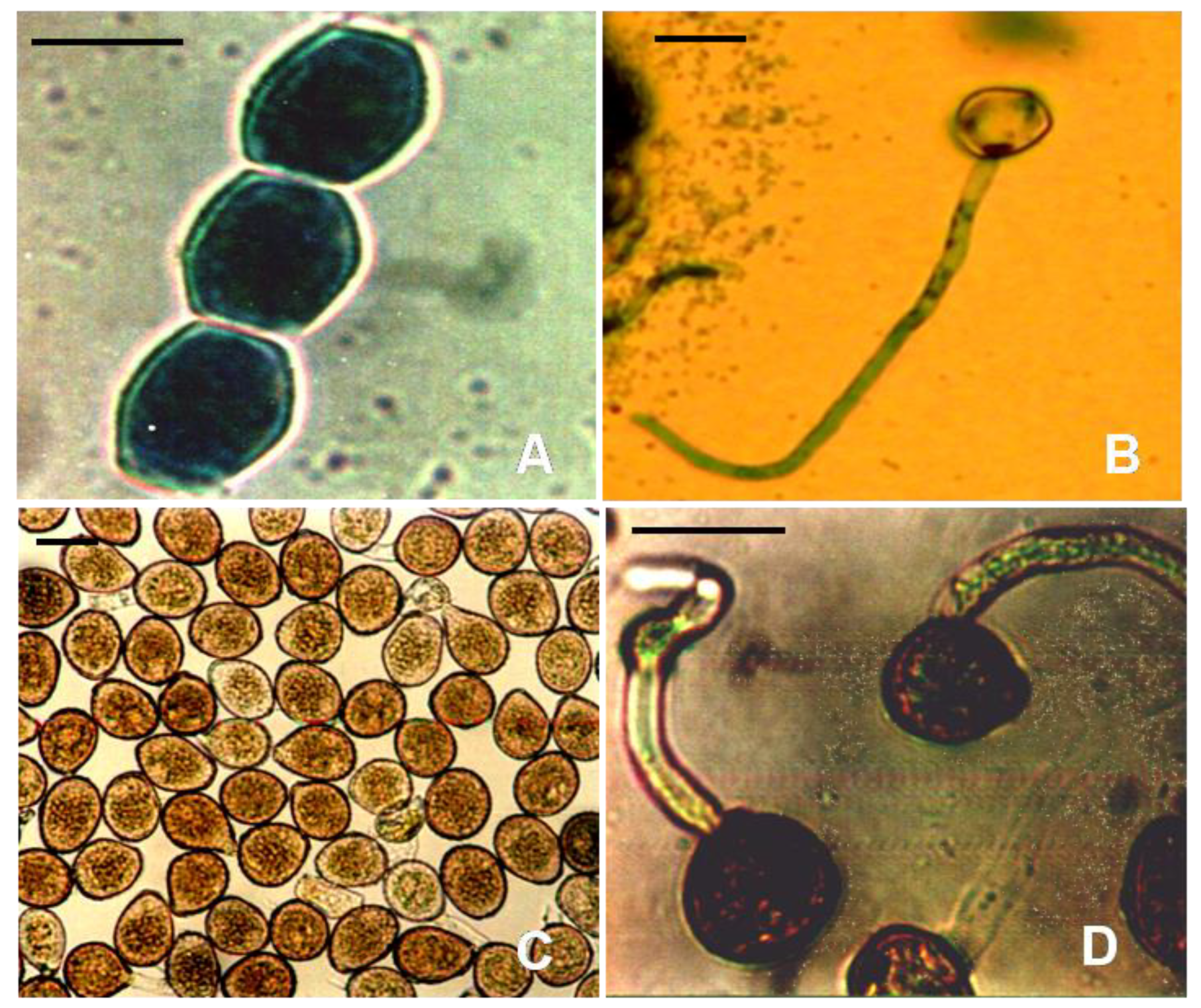
2. Nomenclature, Distribution and Host Range of Uromyces fabae
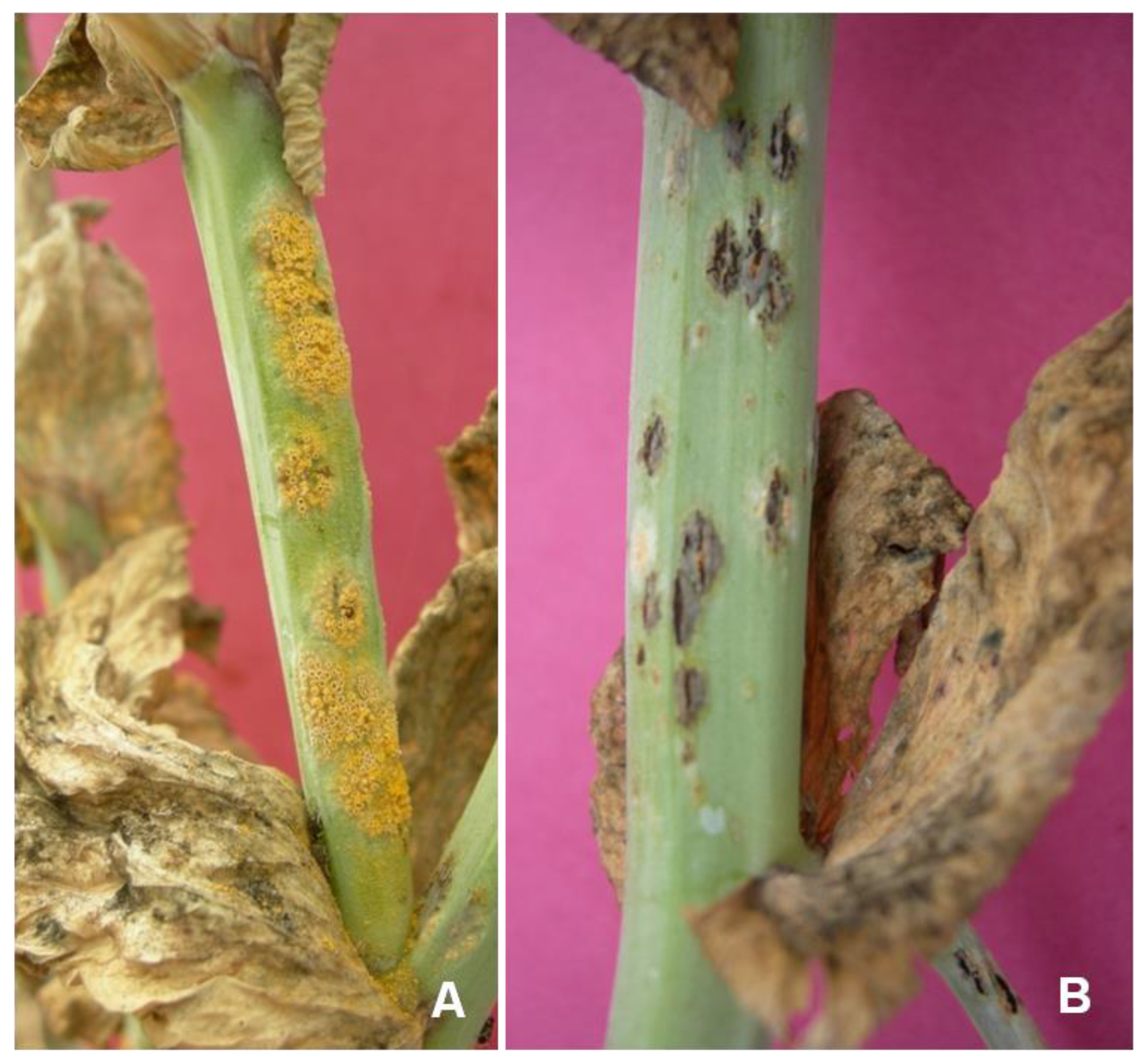
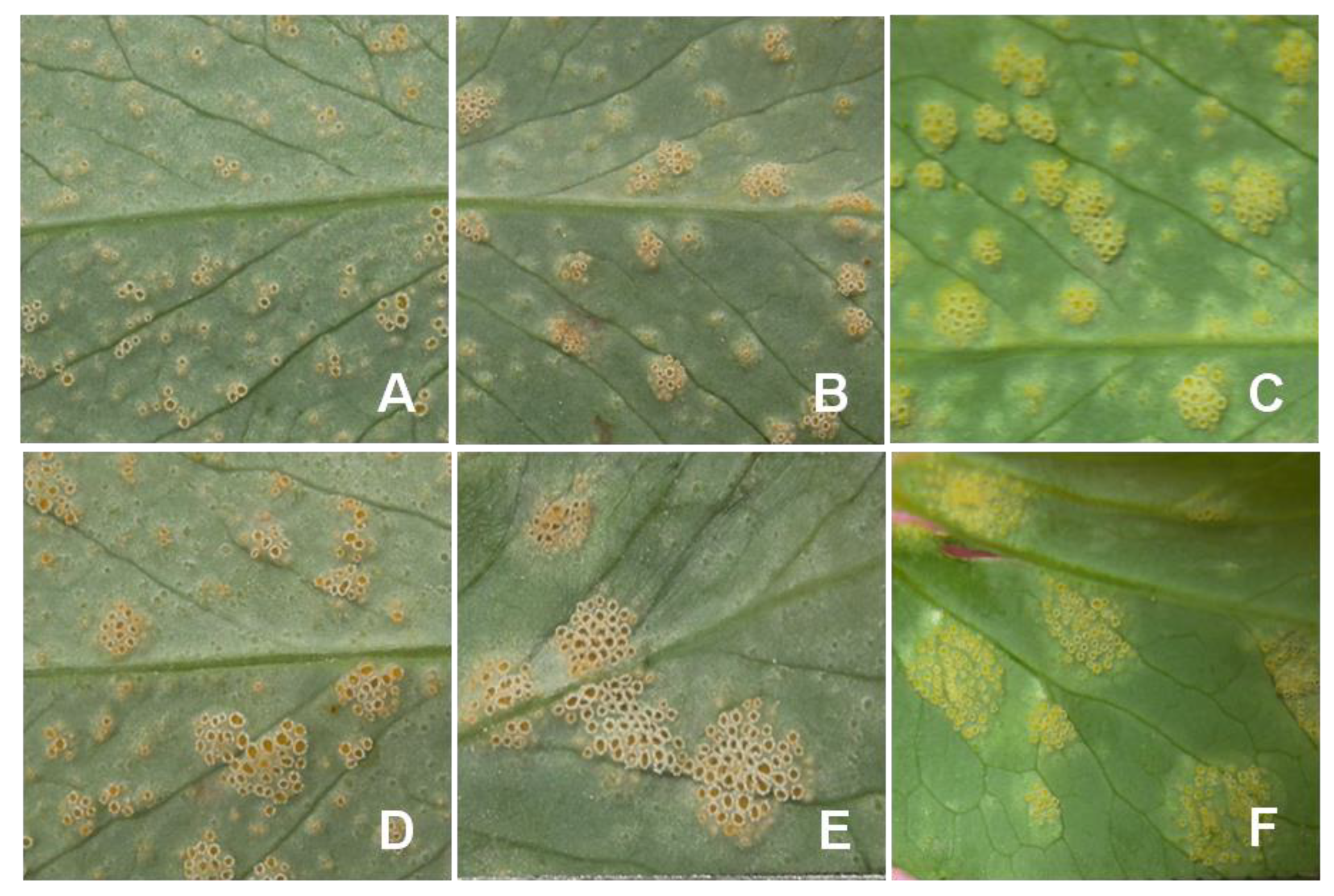
3. Symptoms of Pea Rust
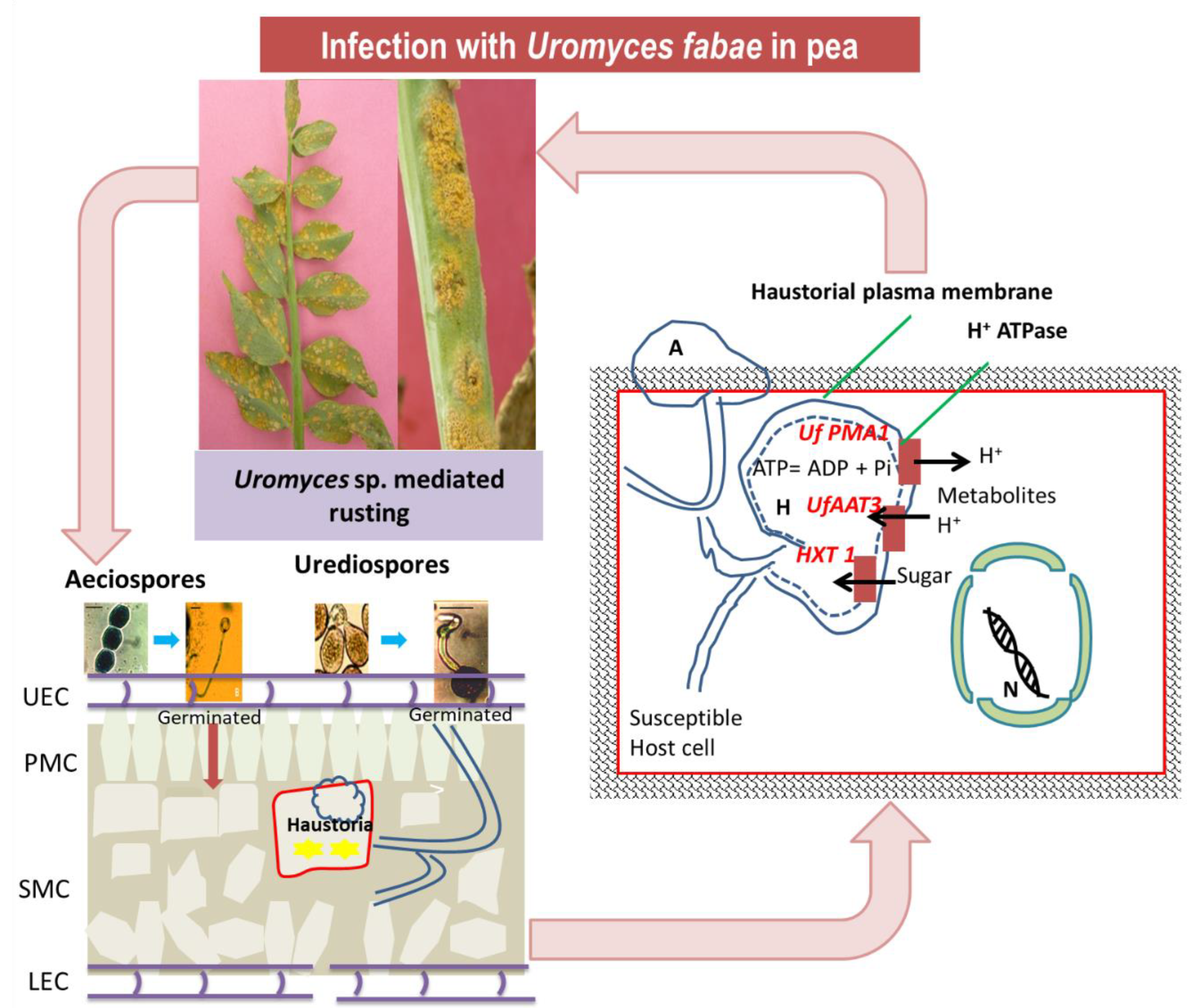
4. Host-Pathogen Interaction

5. Genetics of Rust Resistance in Pea
6. Slow Rusting
6.1. Components of Slow Rusting
6.2. Histopathological Indicators of Slow Rusting
6.2.1. Number and Size of Haustoria
6.2.2. Early Abortive Colonies
6.2.3. Enhanced Lignifications under Infected Conditions
6.3. Biochemicals Associated with Slow Rusting
6.4. Interrelationship among Slow Rusting Components
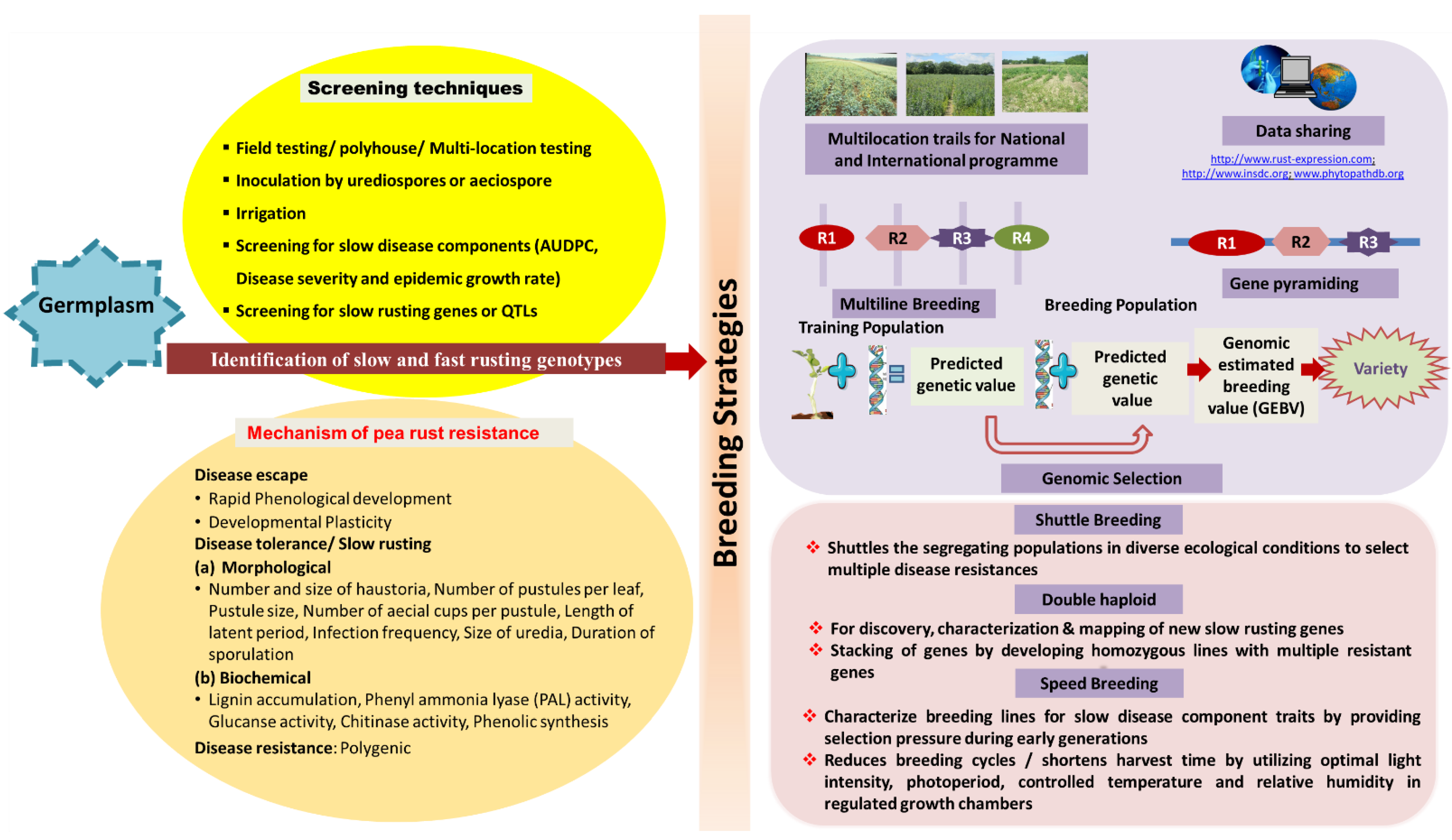
7. Pea Rust Screening
8. Molecular Mapping and Marker-Assisted Selection (MAS)
9. Conclusions and the Future Prospects
- In relation to host specialization, U. viciae-fabae is circumglobal on Lathyrus, Pisum, and Vicia. So, more research is required to achieve the ultimate classification of the U. viciae-fabae complex.
- Identification of physiological races based on a standard set of pea rust differentials is required.
- Survival and recurrence of pea rust pathogen will need to be ascertained by studying the effect of temperature, soil depth, over-summering, and migration on the survivability of urediospore of U. viciae-fabae.
- More work is required on host-specificity and pathogenic variability at the molecular level in U. viciae-fabae to elucidate the differential pathogenicity of isolates.
- Sexual reproduction of this autoecious fungus should be more precisely studied to conclude the possible effects of the matting system on the lack of association between molecular polymorphisms and virulence.
- Hypersensitivity is not reported, and a completely effective source of resistance has not been found. Accumulation of more slow rusting components should be completed in different resistance genotypes to achieve a high level of durable rust resistance.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Graham, P.H.; Vance, C.P. Legumes: Importance and constraints to greater use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, A.G.; Warkentin, T.D. Reactions of field pea varieties to three isolates of Uromyces fabae. Can. J. Plant Sci. 2002, 82, 253–255. [Google Scholar] [CrossRef]
- Vijayalakshmi, S.; Yadav, K.; Kushwaha, C.; Sarode, S.B.; Srivastava, C.P.; Chand, R.; Singh, B.D. Identification of RAPD markers linked to the rust (Uromyces fabae) resistance gene in pea (Pisum sativum). Euphytica 2005, 144, 265–274. [Google Scholar] [CrossRef]
- Kushwaha, C.; Chand, R.; Srivastava, C.P. Role of aeciospores in outbreak of pea (Pisum sativum L.) rust (Uromyces fabae). Eur. J. Plant Pathol. 2006, 115, 323–330. [Google Scholar] [CrossRef]
- Barilli, E.; Sillero, J.C.; Moral, A.; Rubiales, D. Characterization of resistance response of pea (Pisum spp.) against rust (Uromyces pisi). Plant Breed. 2009, 128, 665–670. [Google Scholar] [CrossRef]
- Emeran, A.A.; Sillero, J.C.; Niks, R.E.; Rubiales, D. Infection structures of host-specialized isolates of Uromyces viciae-fabae and of others Uromyces infecting leguminous crops. Plant Dis. 2005, 89, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, A.L.; Singh, V.K. Performance of pea varieties/ lines against powdery mildew and rust. Indian J. Pulses Res. 1994, 7, 92–93. [Google Scholar]
- EPPO Standards Pea. EPPO Bulletin. Available online: https://www.eppo.int/RESOURCES/eppo_standards/pp2_gpp (accessed on 20 July 2022).
- Cummins, G.B. Rust Fungi on Legumes and Composites in North America; The University of Arizona Press: Tucson, AZ, USA, 1987. [Google Scholar]
- Pal, A.B.; Brahmappa, B.; Sohi, H.S.; Rawal, R.D. 1979. Studies on inheritance of resistance to rust (Uromyces fabae Pers. de Bary) on pea. SABRAO J. 1979, 11, 101–103. [Google Scholar]
- Katiyar, R.P.; Ram, R.S. Genetics of rust resistance in pea. Ind. J. Genet. Plant Breed. 1987, 47, 46–48. [Google Scholar]
- Tyagi, M.K.; Srivastava, C.P. Inheritance of powdery mildew and rust resistance in pea. Ann. Biol. 1999, 15, 13–16. [Google Scholar]
- Singh, R.; Ram, H. Inheritance of days to flowering and rust resistance in peas. Res. Crops 2001, 2, 414–418. [Google Scholar]
- Singh, A.K.; Rai, R.; Srivastava, C.P.; Singh, B.D.; Kushwaha, C.; Chand, R. A Quantitative analysis of rust (Uromyces fabae Pers. de-Bary) resistance in pea (Pisum sativum L.) using RILs. Indian J. Agric. Sci. 2012, 82, 190–192. [Google Scholar]
- Chand, R.; Srivastava, C.P.; Singh, B.D.; Sarode, S.B. Identification and characterization of slow rusting components in pea (Pisum sativum L.). Genet. Resour. Crop Evol. 2006, 53, 219–224. [Google Scholar] [CrossRef]
- Kushwaha, C.; Chand, R.; Singh, A.K.; Rai, R.; Srivastava, C.P.; Singh, B.D.; Mohapatra, C. Lignification and early abortive colonies as indicators of partial resistance to rust in pea. Trop. Plant Pathol. 2016, 41, 91–97. [Google Scholar] [CrossRef]
- Link, T.; Seibel, C.; Voegele, R.T. Early insights into the genome sequence of Uromyces fabae. Front. Plant Sci. 2014, 5, 587–591. [Google Scholar] [CrossRef] [Green Version]
- Kreplak, J.; Madoui, M.-A.; Cápal, P.; Novák, P.; Labadie, K.; Aubert, G.; Bayer, P.E.; Gali, K.K.; Syme, R.A.; Main, D.; et al. A reference genome for pea provides insight into legume genome evolution. Nat. Genet. 2019, 51, 1411–1422. [Google Scholar] [CrossRef]
- de Bary, A. Morphologie und Physiologie-Der Plize Flechten und Myxomyceter. 1862. Available online: https://www.biodiversitylibrary.org/item/211464#page/7/mode/1up (accessed on 16 December 2022).
- Arthur, J.C.; Cummins, G.B. Manual of Rusts in United States and Canada; Hafner Publishing Company: New York, NY, USA, 1962. [Google Scholar]
- Gäumann, E.A. Comparative Morphology of Fungi; Biotech Books: New Delhi, India, 1998. [Google Scholar]
- Conner, R.L.; Bernier, C.C. Host range of Uromyces viciae-fabae. Phytopathology 1982, 72, 687–689. [Google Scholar] [CrossRef]
- Emeran, A.A.; Rubiales, D. Effects of eleven fungicides on faba bean rust (Uromyces viciae-fabae) control. In Proceedings of the 4th European Conference on Grain Legumes: Towards the Sustainable Production of Healthy Food, Feed and Novel Products, Cracow, Poland, 8–12 July 2001; Volume 241. [Google Scholar]
- Rubiales, D.; Sillero, J.C. Uromyces viciae-fabae haustorium formation in susceptible and resistant faba bean lines. Eur. J. Plant Pathol. 2003, 109, 71–73. [Google Scholar] [CrossRef]
- Uppal, B.M. International Bulletin on Crop Protection. Int. Inst. Agric. 1933, 7, M103–M746. [Google Scholar]
- Prasada, R.; Verma, U.N. Studies on lentil rust, Uromyces fabae (Pers) de Bary in India. Ind. Phytopathol. 1948, 1, 142–146. [Google Scholar]
- Kapooria, R.G.; Sinha, S. Studies on host range of Uromyces fabae (Persoon) de Bary. Ind. Phytopathol. 1966, 19, 224–230. [Google Scholar]
- Kapooria, R.G.; Sinha, S. Further studies on the host spectrum of Uromyces fabae (Persoon) de Bary in India. Ind. Phytopathol. 1971, 24, 170–171. [Google Scholar]
- Baruah, H.K. Text Book of Plant Pathology; Oxford and IBH Publications: New Delhi, India, 1980. [Google Scholar]
- Barilli, E.; Satovic, Z.; Rubiales, D.; Torres, A.M. Mapping of quantitative trait loci controlling partial resistance against rust incited by Uromyces pisi (Pers.) Wint. in a Pisum fulvum L. intraspecific cross. Euphytica 2010, 175, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Sillero, J.C.; Fondevilla, S.; Davidson, J.; Vaz Patto, M.C.; Warkentin, T.D.; Thomas, J.; Rubiales, D. Screening techniques and sources of resistance to rusts and mildews in grain legumes. Euphytica 2006, 147, 255–272. [Google Scholar] [CrossRef]
- Voegele, R.T. Uromyces fabae: Development, metabolism, and interactions with its host Vicia faba. FEMS Microbiol. Let. 2006, 259, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Voegele, R.T.; Christine, S.; Matthias, H.; Kurt, M. The role of haustoria in sugar supply during infection of broad bean by the rust fungus Uromyces fabae. PNAS 2001, 98, 8133–8138. [Google Scholar] [CrossRef] [Green Version]
- Rubiales, D.; Rojas-Molina, M.M.; Sillero, J.C. Identification of pre- and posthaustorial resistance to rust (Uromyces viciae-fabae) in lentil (Lens culinaris) germplasm. Plant Breed. 2013, 123, 676–680. [Google Scholar] [CrossRef]
- Martins, D.; Araújo, S.D.S.; Rubiales, D.; Vaz Patto, M.C. Legume crops and biotrophic pathogen interactions: A continuous cross-talk of a multilayered array of defense mechanisms. Plants 2020, 9, 1460. [Google Scholar] [CrossRef]
- Kemen, E.; Kemen, A.C.; Rafiqi, M.; Hempel, U.; Mendgen, K.; Hahn, M.; Voegele, R.T. Identification of a protein from rust fungi transferred from haustoria into infected plant cells. MPMI 2005, 18, 1130–1139. [Google Scholar] [CrossRef] [Green Version]
- Struck, C.; Siebels, C.; Rommel, O.; Wernitz, M.; Hahn, M. The plasma membrane H(+)-ATPase from the biotrophic rust fungus Uromyces fabae: Molecular characterization of the gene (PMA1) and functional expression of the enzyme in yeast. MPMI 1998, 11, 458–465. [Google Scholar] [CrossRef]
- Szabo, L.J.; Bushnell, W.R. Hidden robbers: The role of fungal haustoria in parasitism of plants. PNAS 2001, 98, 7654–7655. [Google Scholar] [CrossRef] [Green Version]
- Hahn, M.; Neef, U.; Struck, C.; Göttfert, M.; Mendgen, K. Molecular plant-microbe interactions: A putative amino acid transporter is specifically expressed in haustoria of the rust fungus Uromyces fabae. MPMI 1997, 10, 438–445. [Google Scholar] [CrossRef] [Green Version]
- Struck, C.; Mueller, E.; Martin, H.; Lohaus, G. The Uromyces fabae UfAAT3 gene encodes a general amino acid permease that prefers uptake of in planta scarce amino acids. Mol. Plant Pathol. 2004, 5, 183–189. [Google Scholar] [CrossRef]
- Niks, R.E.; Rubiales, D. Potentially durable resistance mechanisms in plant to specialized fungal pathogens. Euphytica 2002, 124, 201–216. [Google Scholar] [CrossRef]
- Singh, R.M.; Srivastava, C.P. Evaluation classification and usefulness of pea germplasm lines for quantitative characters. Leg. Res. 1985, 8, 68–73. [Google Scholar]
- Kumar, T.B.A.; Rangaswamy, K.T.; Ravi, K. Assessment of tall field pea genotypes for slow rusting resistance. Leg. Res. 1994, 17, 79–82. [Google Scholar]
- Rai, R.; Singh, A.K.; Singh, B.D.; Joshi, A.K.; Chand, R.; Srivastava, C.P. Molecular mapping for resistance to pea rust caused by Uromyces fabae (Pers.) de-Bary. Theor. Appl. Genet. 2011, 123, 803–813. [Google Scholar] [CrossRef]
- Rai, R.; Singh, A.K.; Chand, R.; Srivastava, C.P.; Joshi, A.K.; Singh, B.D. Genomic regions controlling components of resistance for pea rust caused by Uromyces fabae (Pers.) de-Bary. J. Plant Biochem. Biotech. 2017, 25, 133–141. [Google Scholar] [CrossRef]
- Sillero, J.C.; Rubiales, D. Histological characterization of resistance to Uromyces viciae-fabae in faba bean. Phytopathology 2002, 92, 294–299. [Google Scholar] [CrossRef] [Green Version]
- Caldwell, R.M. Breeding for general and/or specific plant disease resistance. In Proceedings of the 3rd International Wheat Genetic Symposium, Canberra, Australia, 5–9 August 1968; pp. 263–272. [Google Scholar]
- Parlevliet, J.E. Components of resistance that reduce the rate of epidemic development. Annu. Rev. Phytopathol. 1979, 17, 203–222. [Google Scholar] [CrossRef]
- Rubiales, D.; Niks, R.E. Characterization of Lr34, a major gene conferring nonhypersensitive resistance to wheat leaf rust. Plant Dis. 1995, 79, 1208–1212. [Google Scholar] [CrossRef]
- Moldenhauer, J.B.M.; Moerschbacher, W.; van der Westhuizen, A.J. Histological investigation of stripe rust (Puccinia striiformis f.sp. tritici) development in resistant and susceptible wheat cultivars. Plant Pathol. 2006, 55, 469–474. [Google Scholar]
- Habtu, A.; Zadoks, J.C. Components of partial resistance in Phaseolus beans against Ethiopian isolates of bean rust. Euphytica 1995, 85, 95–102. [Google Scholar] [CrossRef]
- Conner, R.L.; Bernier, C.C. Slow rusting resistance in Vicia fabae. Can. J. Plant Pathol. 1982, 4, 263–265. [Google Scholar] [CrossRef]
- Marshall, D.R. The advantages and hazards of genetic homogeneity. In The Genetic Basis of Epidemics in Agriculture; Day, P.R., Ed.; New York Academy of Sciences: New York, NY, USA, 1977; pp. 1–20. [Google Scholar]
- Wilcoxson, R.D.; Skovmand, B.; Atif, A.H. Evaluation of wheat cultivars for ability to retard development of stem rust. Ann. Appl. Biol. 1975, 80, 275–281. [Google Scholar] [CrossRef]
- Green, G.J. Virulence changes in Puccinia graminis f. sp. tritici in Canada. Can. J. Bot. 1975, 53, 1377–1386. [Google Scholar] [CrossRef]
- Pederson, W.L.; Leath, S. Pyramiding major genes for resistance to maintain residual effects. Ann. Rev. Phytopathol. 1988, 26, 369–378. [Google Scholar] [CrossRef]
- Gupta, R.P. Evaluation of pea germplasm for their reaction to powdery mildew and rust. Ind. J. Pulses Res. 1990, 3, 186–188. [Google Scholar]
- Chand, R.; Srivastava, C.P.; Kushwaha, C. Screening technique for pea (Pisum sativum L.) genotypes against rust disease (Uromyces fabae Pers de Bary). Ind. J. Agric. Sci. 2004, 74, 166–167. [Google Scholar]
- Wilcoxson, R.D. Genetics of slow rusting in cereals. Phytopathology 1981, 71, 989–993. [Google Scholar] [CrossRef]
- Ohm, H.W.; Shaner, G.E. Three components of slow leaf rusting at different growth stages in wheat. Phytopathology 1976, 66, 1356–1360. [Google Scholar] [CrossRef]
- Kushwaha, C.; Chand, R.; Srivastava, C.P.; Singh, A.K.; Rai, R.; Singh, B.D. Usefulness of number of aecial cups per pustule for selection for slow rusting in pea (Pisum sativum L.) against Uromyces fabae (Pers. de Bary). Ind. J. Agric. Sci. 2010, 80, 933–936. [Google Scholar]
- Matern, U.; Kneusel, R.E. Phenolic compounds in plant disease resistance. Phytoparasitica 1988, 16, 153–170. [Google Scholar] [CrossRef]
- Prats, E.; Rubiales, D.; Jorrín, J. Acibenzolar-Smethyl-induced resistance to sunflower rust (Puccinia helianthi) is associated with an enhancement of coumarins on foliar surface. Physio. Mol. Plant Pathol. 2002, 60, 155–162. [Google Scholar] [CrossRef]
- Hückelhoven, C. Cell wall-associated mechanisms of disease resistance and susceptibility. Ann. Rev. Phytopathol. 2007, 45, 101–127. [Google Scholar] [CrossRef]
- Iriti, M.; Faoro, F. Benzothiadiazole (BTH) induces cell-death independent resistance in Phaseolus vulgaris against Uromyces appendiculatus. J. Phytopathol. 2003, 151, 171–180. [Google Scholar] [CrossRef]
- Kushwaha, C.; Chand, R.; Singh, A.K.; Kumar, M.; Srivastava, C.P. Differential Induction of β-1, 3-Glucanase Gene in Expression of Partial Resistance to Rust (Uromyces fabae (Pers.) de-Bary) in Pea (Pisum sativum L.). Russ. J. Plant Physiol. 2018, 65, 697–701. [Google Scholar] [CrossRef]
- Jeger, M.J.; Viljanen-Rollinson, S.L.H. The use of area under disease progress curve (AUDPC) to assess quantitative disease resistance in crop cultivars. Theor. Appl. Genet. 2001, 102, 32–40. [Google Scholar] [CrossRef]
- Singh, V.K.; Srivastava, C.P.; Chand, R.; Singh, S.P.; Singh, A.K.; Singh, K.K. Variability studies for yield and its contributing characters and AUDPC in pea (Pisum sativum L.). Veg. Sci. 2008, 35, 38–40. [Google Scholar]
- Srivastava, C.P.; Lal, S.; Kushwaha, C.; Chand, R.; Singh, A.K.; Singh, V.K.; Singh, B.D. Genetic variability in the segregating generations and the breeding strategies for selection to improve yield and resistance to rust in pea. J. Food Leg. 2009, 22, 158–161. [Google Scholar]
- Kushwaha, C.; Srivastava, C.P.; Chand, R.; Singh, B.D. Identification and evaluation of critical time for assessment of slow rusting in pea against Uromyces fabae. Field Crop Res. 2007, 103, 1–4. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, V.K.; Chand, R.; Kushwaha, C.; Srivastava, C.P. Evaluation of slow rusting components in pea (Pisum sativum L.). J. Plant Pathol. 2015, 97, 87–92. [Google Scholar]
- Xu, X.; Bai, G.; Carver, B.F.; Shaner, G.E.; Hunger, R.M. Molecular characterization of slow leaf-rusting resistance in wheat. Crop Sci. 2005, 45, 758–765. [Google Scholar] [CrossRef] [Green Version]
- Das, A.; Parihar, A.K.; Saxena, D.; Singh, D.; Singha, K.D.; Kushwaha, K.P.S.; Chand, R.; Bal, R.S.; Chandra, S.; Gupta, S. Deciphering Genotype-by- Environment Interaction for Targeting Test Environments and Rust Resistant Genotypes in Field Pea (Pisum sativum L.). Front. Plant Sci. 2019, 10, 825. [Google Scholar] [CrossRef]
- Pal, A.B.; Brahmappa, B.; Rawal, R.D.; Ullasa, B.A. Field resistance of pea germ plasm to powdery mildew (Erysiphe polygoni) and rust (Uromyces fabae). Plant Dis. 1980, 64, 1085–1086. [Google Scholar] [CrossRef]
- Singh, S.J.; Sokhi, S.S. Evaluation of pea cultivars to Uromyces vicieae fabae. Ind. Phytopathol. 1984, 21, 85–87. [Google Scholar]
- Singh, R.N. A zero to nine point rating scale for evaluation of pea genotypes for resistance to powdery mildew. Narendra Deva J. Agric. Res. 1991, 6, 339–340. [Google Scholar]
- Shaner, G.; Finney, R.E. The effect of nitrogen fertilization on the expression of slow-mildewing resistance in Knox wheat. Phytopathology 1977, 67, 1051–1056. [Google Scholar] [CrossRef] [Green Version]
- Jeger, M.J. Analysis of disease progress as a basis for evaluating disease management practices. Ann. Rev. Phytopathol. 2004, 42, 61–82. [Google Scholar] [CrossRef]
- Yadav, A.S.; Singh, A.K.; Chand, R.; Vaish, S.S. Genetic characterization and population structure of pea (Pisum sativum L.) by molecular markers against rust (Uromyces viciae-fabae) in newly developed genotypes. Sustainability 2022, 14, 15082. [Google Scholar] [CrossRef]
- Young, N.D. QTL mapping and quantitative disease resistance in plants. Ann. Rev. Phytopathol. 1996, 34, 479–501. [Google Scholar] [CrossRef]
- Loridon, K.; McPhee, K.; Morin, J.; Dubreuil, P.; Pilet-Nayel, M.-L.; Aubert, G.; Rameau, C.; Baranger, A.; Coyne, C.; Lejeune-Hènaut, I.; et al. Microsatellite marker polymorphism and mapping in pea (Pisum sativum L.). Theor. Appl. Genet. 2005, 111, 1022–1031. [Google Scholar] [CrossRef]
- Avila, C.M.; Sillero, J.C.; Rubiales, D.; Moreno, M.T.; Torres, A.M. Identification of RAPD markers linked to the Uvf-1 gene conferring hypersensitive resistance against rust (Uromyces viciae-fabae) in Vicia faba L. Theor. Appl. Genet. 2003, 107, 353–358. [Google Scholar] [CrossRef]
- Saha, G.C.; Sarker, A.; Chen, W.; Vandemark, G.J.; Muehlbauer, F.J. Identification of markers associated with genes for rust resistance in Lens culinaris Medik. Euphytica 2010, 175, 261–265. [Google Scholar] [CrossRef]
- Mekonnen, F.; Mekbib, F.; Kumar, S.; Ahmed, S.; Chahoe, R.K. Identification of molecular markers associated with rust (Uromyces vicia-fabae Pers.) resistance genes in lentil (Lens culinaris subsp. culinaris). Can. J. Crop Prot. 2014, 2, 27–36. [Google Scholar]
- Upadhyay, V.; Kushwaha, K.P.S.; Pandey, P. Molecular screening of pea germplasm for rust disease resistance using SSR Marker. J. Pure Appl. Microbiol. 2017, 11, 343–348. [Google Scholar] [CrossRef]
- Ijaz, U.; Sudheesh, S.; Kaur, S.; Sadeque, A.; Bariana, H.; Bansal, U.; Adhikari, K. Mapping of two new rust resistance genes Uvf-2 and Uvf-3 in faba bean. Agronomy 2021, 11, 1370. [Google Scholar] [CrossRef]
- Singh, J.; Sirari, A.; Singh, H.; Kumar, A.; Jaidka, M.; Mandahal, K.S.; Kumar, S.; Singh, S. Identifying and validating SSR markers linked with rust resistance in lentil (Lens culinaris). Plant Breed. 2021, 140, 477–485. [Google Scholar] [CrossRef]
- Rubiales, D.; Castillejo, M.A.; Madrid, E.; Barilli, E.; Rispail, N. Legume breeding for rust resistance: Lessons to learn from the model Medicago truncatula. Euphytica 2011, 180, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Hurtado-Gonzales, O.P.; Valentini, G.; Gilio, T.A.; Martins, A.M.; Song, Q.; Pastor-Corrales, M.A. Fine mapping of Ur-3, a historically important rust resistance locus in common bean. G3 Genes Genomes Genet. 2017, 7, 557–569. [Google Scholar] [CrossRef] [Green Version]
- Acevedo, M.; Steadman, J.R.; Rosas, J.C.; Venegas, J. Characterization of virulence diversity of the bean rust pathogen Uromyces appendiculatus in wild bean populations as a tool for effective resistance gene deployment. Annu. Rep.-Bean Improv. Coop. 2005, 48, 132. [Google Scholar]
- Miklas, P.N.; Pastor-Corrales, M.A.; Jung, G.; Coyne, D.P.; Kelly, J.D.; McClean, P.E.; Gepts, P. Comprehensive linkage map of bean rust resistance genes. Annu. Rep.-Bean Improv. Coop. 2002, 45, 125–129. [Google Scholar]
- Park, S.O.; Coyne, D.P.; Steadman, J.R.; Skroch, P.W. Mapping of the Ur-7 gene for specific resistance to rust in common bean. Crop Sci. 2003, 43, 1470–1476. [Google Scholar] [CrossRef]
- Wu, X.; Wang, B.; Xin, Y.; Wang, Y.; Tian, S.; Wang, J.; Li, G. Unravelling the Genetic Architecture of Rust Resistance in the Common Bean (Phaseolus vulgaris L.) by Combining QTL-Seq and GWAS Analysis. Plants 2022, 11, 953. [Google Scholar] [CrossRef]
- Alleyne, A.T.; Steadman, J.R.; Eskridge, K.M. Monitoring changing virulence patterns of Uromyces appendiculatus in the resistant pinto bean cultivar Olathe by rep-PCR. Eur. J. Plant Pathol. 2008, 122, 315–319. [Google Scholar] [CrossRef]
- Jung, G.; Coyne, D.P.; Bokosi, J.; Steadman, J.R.; Nienhuis, J. Mapping genes for specific and adult plant resistance to rust and abaxial leaf pubescence and their genetic relationships using randomly amplified polymorphic DNA (RAPD) markers in common bean. J. Am. Soc. Horti. Sci. 1998, 123, 859–863. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Character | Uromyces pisi | U. viciae-fabae/U. fabae |
| Occurrence | Temperate regions e.g., Europe, Australia, Canada | Tropical and sub-tropical regions, e.g., India, China |
| Conducive weather | Comparatively cooler and less humid | Warm humid |
| Fungus | Heteroecious macrocyclic fungus completes life-cycle on Cypress spurge (Euphorbia cyparissias L.) | Autoecious macrocyclic |
| Infecting stage | Uredial | Uredial and acidial |
| Repeating spore | Urediospores | Aeciospores/Urediospores |
| Infection structures | Substomatal vesicles (SSVs) of U. pisi are oblong to oval, with both ends curved, and form one primary infection hyphae (PIH) each. | SSVs of U. viciae-fabae are variable in shape ranging from fusiform to cylindrical tubes, oval to globoid croissant-like, sausage-like, or triangular, and generally form two PIH. |
| ITS markers | ITS1 region has a unique 90 bp deletion region | No such deletion region found |
| Genetics of resistance | Polygenic | Single major gene to polygenic |
| Markers | Marker | Distance from the Resistance Gene | Host Crop | Parents | Reference |
| OPD13736, OPL181032 & OPI20900 | RAPD | - | Faba bean | 2N52 (resistant) & VF-176 (susceptible) | [82] |
| SC10-82360 & SCRI-711000 | RAPD | 10.8 cM and 24.5 cM from the Ruf gene | Pea | HUVP 1 (HUVP 1 × FC 1) | [3] |
| F7XEM4a | SRAP | 7.9 cM | Lentil | ILL-4605 (resistant) & ILL-5888 (susceptible) | [83] |
| AD146 & AA416 | SSR | 7.3 cM | Pea | HUVP 1 (susceptible) & FC 1 (resistant) | [44] |
| GLLC106 | SSR | 10cM | Lentil | FLIP-2004-7L (resistant) × L-9–12 (susceptible) | [84] |
| A446-AA505 and AD146-AA416 | SSR | 10.8 cM | Pea | HUVP 1 (susceptible) and FC 1 (resistant) | [45] |
| AA446, AA505, AD146 & AA416 | SSR | - | Pea | Pant P 244, Pant P 42 | [85] |
| KASP_Vf_0703 & KASP_C250539 | KASP | 4.9 cM & 2.9 cM from Uvf-2 | Faba bean | Doza#12034 × Ac1655 (resistant) × Fiord (susceptible) | [86] |
| KASP_Ac×F165 & KASP_vf_1090 | 2.5 cM & 10.1 cM from Uvf-3 | ||||
| LcSSR440 & LcSSR606 | SSR | 8.3 and 8.1cM | Lentil | FLIP-2004-7L (resistant) × L-9–12 (susceptible) | [87] |
| Resistant Gene/Locus Chromosome | Linkage Group/Chromosome | Donor Genotype | Corresponding Pathotype | References |
| Uvf-1 gene | - | 2N52 | Race 1 | [88] |
| Uf2 | Chromosome 3 | #12034 (Doza) Ac1655 | pathotype 24–40 | [86] |
| Ur-3+ | - | Mex 235 | - | [89] |
| Ur-11 | B11 | PI 181996 | - | [90] |
| Ur-3 | B11 Chromosome 5 | Aurora and NEP-2 | Race 44, 63 | [89,91] |
| Ur-4 | LG6 (or B6) | Early Gallatin (EG) | Race 63 | [92] |
| Ur-5 | LG4 (or B4) | GN BelNeb-RR-1 | Race 59, 63 | [92,93] |
| Ur-6 | B11 | Golden Gate Wax and Olathe | Races 49, 67, and 108 | [91,94] |
| Ur-7 | LG 11 | GN 1140 | Race59 | [92] |
| Ur-9, | LG1 | PC 50 | A88TI-20a & D82C1-1 | [91,95] |
| Ur-12 | LG 4b | PC 50 | A88TI-4b | [91,95] |
| Qruf2 and Qruf3 | LGI & LGVI | FC 1 | - | [45] |
| One major (Qruf) and one minor (Qruf1) QTL | LGVII | FC 1 | - | [45] |
| Qruf and/or Qruf1 | LGVII | Pant P 42 | - | [85] |
| Uvf-2, Uvf-3 | chromosomes III and V | Doza#12034 & Ac1655 | pathotype 24–40 | [86] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, A.K.; Kushwaha, C.; Shikha, K.; Chand, R.; Mishra, G.P.; Dikshit, H.K.; Devi, J.; Aski, M.S.; Kumar, S.; Gupta, S.; et al. Rust (Uromyces viciae-fabae Pers. de-Bary) of Pea (Pisum sativum L.): Present Status and Future Resistance Breeding Opportunities. Genes 2023, 14, 374. https://doi.org/10.3390/genes14020374
Singh AK, Kushwaha C, Shikha K, Chand R, Mishra GP, Dikshit HK, Devi J, Aski MS, Kumar S, Gupta S, et al. Rust (Uromyces viciae-fabae Pers. de-Bary) of Pea (Pisum sativum L.): Present Status and Future Resistance Breeding Opportunities. Genes. 2023; 14(2):374. https://doi.org/10.3390/genes14020374
Chicago/Turabian StyleSingh, Anil Kumar, Chanda Kushwaha, Kumari Shikha, Ramesh Chand, Gyan P. Mishra, Harsh Kumar Dikshit, Jyoti Devi, Muraleedhar S. Aski, Shiv Kumar, Sanjeev Gupta, and et al. 2023. "Rust (Uromyces viciae-fabae Pers. de-Bary) of Pea (Pisum sativum L.): Present Status and Future Resistance Breeding Opportunities" Genes 14, no. 2: 374. https://doi.org/10.3390/genes14020374
APA StyleSingh, A. K., Kushwaha, C., Shikha, K., Chand, R., Mishra, G. P., Dikshit, H. K., Devi, J., Aski, M. S., Kumar, S., Gupta, S., & Nair, R. M. (2023). Rust (Uromyces viciae-fabae Pers. de-Bary) of Pea (Pisum sativum L.): Present Status and Future Resistance Breeding Opportunities. Genes, 14(2), 374. https://doi.org/10.3390/genes14020374