
Brain Targeted AAV1-GALC Gene Therapy Reduces Psychosine and Extends Lifespan in a Mouse Model of Krabbe Disease

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Care and Neonatal AAV1-GALC Particle Injection
2.2. Construction and Production of AAV1-GALC Viral Particles
2.3. Determination of Psychosine Levels
2.4. Survivability and Body Weight Analysis
2.5. Immunohistochemistry
2.6. Luxol-Fast Blue (LFB)/Periodic Acid Schiff (PAS) Staining
2.7. Liver GALC Analysis
2.8. Statistical Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kohlschutter, A. Lysosomal leukodystrophies: Krabbe disease and metachromatic leukodystrophy. Handb. Clin. Neurol. 2013, 113, 1611–1618. [Google Scholar] [PubMed]
- Suzuki, K.; Suzuki, Y. Globoid cell leucodystrophy (Krabbe’s disease): Deficiency of galactocerebroside beta-galactosidase. Proc. Natl. Acad. Sci. USA 1970, 66, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Nagara, H.; Kobayashi, T.; Goto, I. The twitcher mouse: Accumulation of galactosylsphingosine and pathology of the sciatic nerve. Brain Res. 1988, 454, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Nagara, H.; Ogawa, H.; Sato, Y.; Kobayashi, T.; Suzuki, K. The twitcher mouse: Degeneration of oligodendrocytes in vitro. Dev. Brain Res. 1986, 26, 79–84. [Google Scholar] [CrossRef]
- Seitelberger, F. Demyelination and leukodystrophy at an early age. Bol. Estud. Med. Biol. 1981, 31, 373–382. [Google Scholar]
- Escolar, M.L.; Poe, M.D.; Provenzale, J.M.; Richards, K.C.; Allison, J.; Wood, S.; Wenger, D.A.; Pietryga, D.; Wall, D.; Champagne, M.; et al. Transplantation of umbilical-cord blood in babies with infantile Krabbe’s disease. N. Engl. J. Med. 2005, 352, 2069–2081. [Google Scholar] [CrossRef] [Green Version]
- Yoon, I.C.; Bascou, N.A.; Poe, M.D.; Szabolcs, P.; Escolar, M.L. Long-term neurodevelopmental outcomes of hematopoietic stem cell transplantation for late-infantile Krabbe disease. Blood 2021, 137, 1719–1730. [Google Scholar] [CrossRef]
- Mitsutake, A.; Matsukawa, T.; Iwata, A.; Ishiura, H.; Mitsui, J.; Mori, H.; Toya, T.; Honda, A.; Kurokawa, M.; Sakai, N.; et al. Favorable outcome of hematopoietic stem cell transplantation in late-onset Krabbe disease. Brain Dev. 2023, 45, 408–412. [Google Scholar] [CrossRef]
- Kobayashi, T.; Yamanaka, T.; Jacobs, J.M.; Teixeira, F.; Suzuki, K. The twitcher mouse: An enzymatically authentic model of human globoid cell leukodystrophy (Krabbe disease). Brain Res. 1980, 202, 479–483. [Google Scholar] [CrossRef]
- Suzuki, K. The twitcher mouse: A model for Krabbe disease and for experimental therapies. Brain Pathol. 1995, 5, 249–258. [Google Scholar] [CrossRef]
- Lee, W.C.; Tsoi, Y.K.; Dickey, C.A.; DeLucia, M.W.; Dickson, D.W.; Eckman, C.B. Suppression of galactosylceramidase (GALC) expression in the twitcher mouse model of globoid cell leukodystrophy (GLD) is caused by nonsense-mediated mRNA decay (NMD). Neurobiol. Dis. 2006, 23, 273–280. [Google Scholar] [CrossRef]
- Mitsuo, K.; Kobayashi, T.; Shinnoh, N.; Goto, I. Biosynthesis of galactosylsphingosine (Psychosine) in the twitcher mouse. Neurochem. Res. 1989, 14, 899–903. [Google Scholar] [CrossRef]
- Ichioka, T.; Kishimoto, Y.; Brennan, S.; Santos, G.W.; Yeager, A.M. Hematopoietic cell transplantation in murine globoid cell leukodystrophy (the twitcher mouse): Effects on levels of galactosylceramidase, psychosine, and galactocerebrosides. Proc. Natl. Acad. Sci. USA 1987, 84, 4259–4263. [Google Scholar] [CrossRef]
- Luzi, P.; Rafi, M.A.; Wenger, D.A. Characterization of the large deletion in the GALC gene found in patients with Krabbe disease. Hum. Mol. Genet. 1995, 4, 2335–2338. [Google Scholar] [CrossRef]
- Lee, W.C.; Courtenay, A.; Troendle, F.J.; Stallings-Mann, M.L.; Dickey, C.A.; Delucia, M.W.; Dickson, D.W.; Eckman, C.B. Enzyme replacement therapy results in substantial improvements in early clinical phenotype in a mouse model of globoid cell leukodystrophy. FASEB J. 2005, 19, 1549–1551. [Google Scholar] [CrossRef]
- Olmstead, C.E. Neurological and neurobehavioral development of the mutant ‘twitcher’ mouse. Behav. Brain Res. 1987, 25, 143–153. [Google Scholar] [CrossRef]
- Kobayashi, S.; Katayama, M.; Bourque, E.; Suzuki, K.; Suzuki, K. The twitcher mouse: Positive immunohistochemical staining of globoid cells with monoclonal antibody against Mac-1 antigen. Dev. Brain Res. 1985, 20, 49–54. [Google Scholar] [CrossRef]
- Snook, E.R.; Fisher-Perkins, J.M.; Sansing, H.A.; Lee, K.M.; Alvarez, X.; MacLean, A.G.; Peterson, K.E.; Lackner, A.A.; Bunnell, B.A. Innate Immune Activation in the Pathogenesis of a Murine Model of Globoid Cell Leukodystrophy. Am. J. Pathol. 2013, 184, 382–396. [Google Scholar] [CrossRef] [Green Version]
- Taniike, M.; Suzuki, K. Spacio-temporal progression of demyelination in twitcher mouse: With clinico-pathological correlation. Acta Neuropathol. 1994, 88, 228–236. [Google Scholar] [CrossRef]
- Wilson, I.; Vitelli, C.; Yu, G.K.; Pacheco, G.; Vincelette, J.; Bunting, S.; Sisó, S. Quantitative Assessment of Neuroinflammation, Myelinogenesis, Demyelination, and Nerve Fiber Regeneration in Immunostained Sciatic Nerves From Twitcher Mice with a Tissue Image Analysis Platform. Toxicol. Pathol. 2021, 49, 950–962. [Google Scholar] [CrossRef]
- Sutter, P.A.; Ménoret, A.; Jellison, E.R.; Nicaise, A.M.; Bradbury, A.M.; Vella, A.T.; Bongarzone, E.R.; Crocker, S.J. CD8+ T cell depletion prevents neuropathology in a mouse model of globoid cell leukodystrophy. J. Exp. Med. 2023, 220, e20221862. [Google Scholar] [CrossRef] [PubMed]
- Ellison, S.; Parker, H.; Bigger, B. Advances in therapies for neurological lysosomal storage disorders. J. Inherit. Metab. Dis. 2023. [Google Scholar] [CrossRef] [PubMed]
- Kido, J.; Sugawara, K.; Nakamura, K. Gene therapy for lysosomal storage diseases: Current clinical trial prospects. Front. Genet. 2023, 14, 1064924. [Google Scholar] [CrossRef] [PubMed]
- The Guide for the Care and Use of Laboratory Animals. ILAR J. 2016, 57, NP. [CrossRef] [Green Version]
- Passini, M.A.; Watson, D.J.; Vite, C.H.; Landsburg, D.J.; Feigenbaum, A.L.; Wolfe, J.H. Intraventricular brain injection of adeno-associated virus type 1 (AAV1) in neonatal mice results in complementary patterns of neuronal transduction to AAV2 and total long-term correction of storage lesions in the brains of beta-glucuronidase-deficient mice. J. Virol. 2003, 77, 7034–7040. [Google Scholar]
- Chakrabarty, P.; Rosario, A.; Cruz, P.; Siemienski, Z.; Ceballos-Diaz, C.; Crosby, K.; Jansen, K.; Borchelt, D.R.; Kim, J.-Y.; Jankowsky, J.L.; et al. Capsid Serotype and Timing of Injection Determines AAV Transduction in the Neonatal Mice Brain. PLoS ONE 2013, 8, e67680. [Google Scholar] [CrossRef]
- Klein, R.L.; Hamby, M.E.; Gongb, Y.; Hirko, A.C.; Wangc, S.; Hughes, J.A.; King, M.A.; Meyer, E.M. Dose and Promoter Effects of Adeno-Associated Viral Vector for Green Fluorescent Protein Expression in the Rat Brain. Exp. Neurol. 2002, 176, 66–74. [Google Scholar] [CrossRef]
- Sheehan, D.; CaH, B.B. Theory and Practice of Histotechnology, 2nd ed.; The CV Mosby Company: St. Louis, MO, USA, 1980. [Google Scholar]
- Zhu, H.; Lopez-Rosas, A.; Qiu, X.; Van Breemen, R.B.; Bongarzone, E.R. Detection of the Neurotoxin Psychosine in Samples of Peripheral Blood: Application in Diagnostics and Follow-up of Krabbe Disease. Arch. Pathol. Lab. Med. 2012, 136, 709–710. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Grover, W.D. Krabbe’s leukocystrophy (globoid cell leukodystrophy). An ultrastructural study. Am. J. Obstet. Gynecol. 1970, 106, 385–396. [Google Scholar]
- Jesionek-Kupnicka, D.; Majchrowska, A.; Krawczyk, J.; Wendorff, J.; Barcikowska, M.; Lukaszek, S.; Liberski, P.P. Krabbe disease: An ultrastructural study of globoid cells and reactive astrocytes at the brain and optic nerves. Folia Neuropathol. 1997, 35, 155–162. [Google Scholar]
- Iacono, D.; Koga, S.; Peng, H.; Manavalan, A.; Daiker, J.; Castanedes-Casey, M.; Martin, N.B.; Herdt, A.R.; Gelb, M.H.; Dickson, D.W.; et al. Galactosylceramidase deficiency and pathological abnormalities in cerebral white matter of Krabbe disease. Neurobiol. Dis. 2022, 174, 105862. [Google Scholar] [CrossRef]
- Mingozzi, F.; Meulenberg, J.J.; Hui, D.J.; Basner-Tschakarjan, E.; Hasbrouck, N.C.; Edmonson, S.A.; Hutnick, N.A.; Betts, M.R.; Kastelein, J.J.; Stroes, E.S.; et al. AAV-1–mediated gene transfer to skeletal muscle in humans results in dose-dependent activation of capsid-specific T cells. Blood 2009, 114, 2077–2086. [Google Scholar] [CrossRef]
- Brantly, M.L.; Chulay, J.D.; Wang, L.; Mueller, C.; Humphries, M.; Spencer, L.T.; Rouhani, F.; Conlon, T.J.; Calcedo, R.; Betts, M.R.; et al. Sustained transgene expression despite T lymphocyte responses in a clinical trial of rAAV1-AAT gene therapy. Proc. Natl. Acad. Sci. USA 2009, 106, 16363–16368. [Google Scholar] [CrossRef]
- Mendell, J.R.; Rodino-Klapac, L.R.; Rosales-Quintero, X.; Kota, J.; Coley, B.D.; Galloway, G.; Craenen, J.M.; Lewis, S.; Malik, V.; Shilling, C.; et al. Limb-girdle muscular dystrophy type 2D gene therapy restores α-sarcoglycan and associated proteins. Ann. Neurol. 2009, 66, 290–297. [Google Scholar] [CrossRef]
- Hadaczek, P.; Stanek, L.; Ciesielska, A.; Sudhakar, V.; Samaranch, L.; Pivirotto, P.; Bringas, J.; O’Riordan, C.; Mastis, B.; San Sebastian, W.; et al. Widespread AAV1- and AAV2-mediated transgene expression in the nonhuman primate brain: Implications for Huntington’s disease. Mol. Ther. Methods Clin. Dev. 2016, 3, 16037. [Google Scholar] [CrossRef]
- Watakabe, A.; Ohtsuka, M.; Kinoshita, M.; Takaji, M.; Isa, K.; Mizukami, H.; Ozawa, K.; Isa, T.; Yamamori, T. Comparative analyses of adeno-associated viral vector serotypes 1, 2, 5, 8 and 9 in marmoset, mouse and macaque cerebral cortex. Neurosci. Res. 2014, 93, 144–157. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, Y.; Hirai, Y.; Miyake, K.; Shimada, T. Targeted gene transfer into ependymal cells through intraventricular injection of AAV1 vector and long-term enzyme replacement via the CSF. Sci. Rep. 2014, 4, 5506. [Google Scholar] [CrossRef] [Green Version]
- Aschauer, D.F.; Kreuz, S.; Rumpel, S. Analysis of Transduction Efficiency, Tropism and Axonal Transport of AAV Serotypes 1, 2, 5, 6, 8 and 9 in the Mouse Brain. PLoS ONE 2013, 8, e76310. [Google Scholar] [CrossRef] [Green Version]
- Ridder, R.; Geisse, S.; Kleuser, B.; Kawalleck, P.; Gram, H. A COS-cell-based system for rapid production and quantification of scFv::IgC kappa antibody fragments. Gene 1995, 166, 273–276. [Google Scholar] [CrossRef]
- Rafi, M.A.; Rao, H.Z.; Passini, M.A.; Curtis, M.; Vanier, M.T.; Zaka, M.; Luzi, P.; Wolfe, J.H.; Wenger, D.A. AAV-Mediated expression of galactocerebrosidase in brain results in attenuated symptoms and extended life span in murine models of globoid cell leukodystrophy. Mol. Ther. 2005, 11, 734–744. [Google Scholar] [CrossRef]
- Hordeaux, J.; Jeffrey, B.A.; Jian, J.; Choudhury, G.R.; Michalson, K.; Mitchell, T.W.; Buza, E.L.; Chichester, J.; Dyer, C.; Bagel, J.; et al. Efficacy and Safety of a Krabbe Disease Gene Therapy. Hum. Gene Ther. 2022, 33, 499–517. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Fantz, C.R.; Levy, B.; Rafi, M.A.; Vogler, C.; Wenger, D.A.; Sands, M.S. AAV2/5 vector expressing galactocerebrosidase ameliorates CNS disease in the murine model of globoid-cell leukodystrophy more efficiently than AAV2. Mol. Ther. 2005, 12, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.-S.; Hsiao, C.-D.; Liau, I.; Lin, S.-P.; Chiang, M.-F.; Chuang, C.-K.; Wang, T.-J.; Wu, T.-Y.; Jian, Y.-R.; Huang, S.-F.; et al. CNS-targeted AAV5 gene transfer results in global dispersal of vector and prevention of morphological and function deterioration in CNS of globoid cell leukodystrophy mouse model. Mol. Genet. Metab. 2011, 103, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Rafi, M.A.; Rao, H.Z.; Luzi, P.; Curtis, M.T.; Wenger, D.A. Extended Normal Life After AAVrh10-mediated Gene Therapy in the Mouse Model of Krabbe Disease. Mol. Ther. 2012, 20, 2031–2042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafi, M.A.; Luzi, P.; Wenger, D.A. Can early treatment of twitcher mice with high dose AAVrh10-GALC eliminate the need for BMT? Bioimpacts 2021, 11, 135–146. [Google Scholar] [CrossRef]
- Karumuthil-Melethil, S.; Marshall, M.S.; Heindel, C.; Jakubauskas, B.; Bongarzone, E.R.; Gray, S.J. Intrathecal administration of AAV/GALC vectors in 10-11-day-old twitcher mice improves survival and is enhanced by bone marrow transplant. J. Neurosci. Res. 2016, 94, 1138–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafi, M.A.; Fugaro, J.; Amini, S.; Luzi, P.; de Gala, G.; Victoria, T.; Dubell, C.; Shahinfar, M.; Wenger, D.A. Retroviral Vector-Mediated Transfer of the Galactocerebrosidase (GALC) cDNA Leads to Overexpression and Transfer of GALC Activity to Neighboring Cells. Biochem. Mol. Med. 1996, 58, 142–150. [Google Scholar] [CrossRef]
- Spratley, S.J.; Hill, C.H.; Viuff, A.H.; Edgar, J.R.; Skjødt, K.; Deane, J.E. Molecular Mechanisms of Disease Pathogenesis Differ in Krabbe Disease Variants. Traffic 2016, 17, 908–922. [Google Scholar] [CrossRef]
- Hill, C.H.; Cook, G.M.; Spratley, S.J.; Fawke, S.; Graham, S.C.; Deane, J.E. The mechanism of glycosphingolipid degradation revealed by a GALC-SapA complex structure. Nat. Commun. 2018, 9, 151. [Google Scholar] [CrossRef] [Green Version]
- Potter, G.B.; Santos, M.; Davisson, M.T.; Rowitch, D.H.; Marks, D.L.; Bongarzone, E.R.; Petryniak, M.A. Missense mutation in mouse GALC mimics human gene defect and offers new insights into Krabbe disease. Hum. Mol. Genet. 2013, 22, 3397–3414. [Google Scholar] [CrossRef] [Green Version]
- Rebiai, R.; Rue, E.; Zaldua, S.; Nguyen, D.; Scesa, G.; Jastrzebski, M.; Foster, R.; Wang, B.; Jiang, X.; Tai, L.; et al. CRISPR-Cas9 Knock-In of T513M and G41S Mutations in the Murine beta-Galactosyl-Ceramidase Gene Re-capitulates Early-Onset and Adult-Onset Forms of Krabbe Disease. Front. Mol. Neurosci. 2022, 15, 896314. [Google Scholar] [CrossRef]
- Lin, D.-S.; Hsiao, C.-D.; Lee, A.Y.-L.; Ho, C.-S.; Liu, H.-L.; Wang, T.-J.; Jian, Y.-R.; Hsu, J.-C.; Huang, Z.-D.; Lee, T.-H.; et al. Mitigation of cerebellar neuropathy in globoid cell leukodystrophy mice by AAV-mediated gene therapy. Gene 2015, 571, 81–90. [Google Scholar] [CrossRef]
- Feltri, M.L.; Weinstock, N.I.; Favret, J.; Dhimal, N.; Wrabetz, L.; Shin, D. Mechanisms of demyelination and neurodegeneration in globoid cell leukodystrophy. Glia 2021, 69, 2309–2331. [Google Scholar] [CrossRef]
- Nasir, G.; Chopra, R.; Elwood, F.; Ahmed, S.S. Krabbe Disease: Prospects of Finding a Cure Using AAV Gene Therapy. Front. Med. 2021, 8, 760236. [Google Scholar] [CrossRef]
- Heller, G.; Bradbury, A.M.; Sands, M.S.; Bongarzone, E.R. Preclinical studies in Krabbe disease: A model for the investigation of novel combination therapies for lysosomal storage diseases. Mol. Ther. 2023, 31, 7–23. [Google Scholar] [CrossRef]





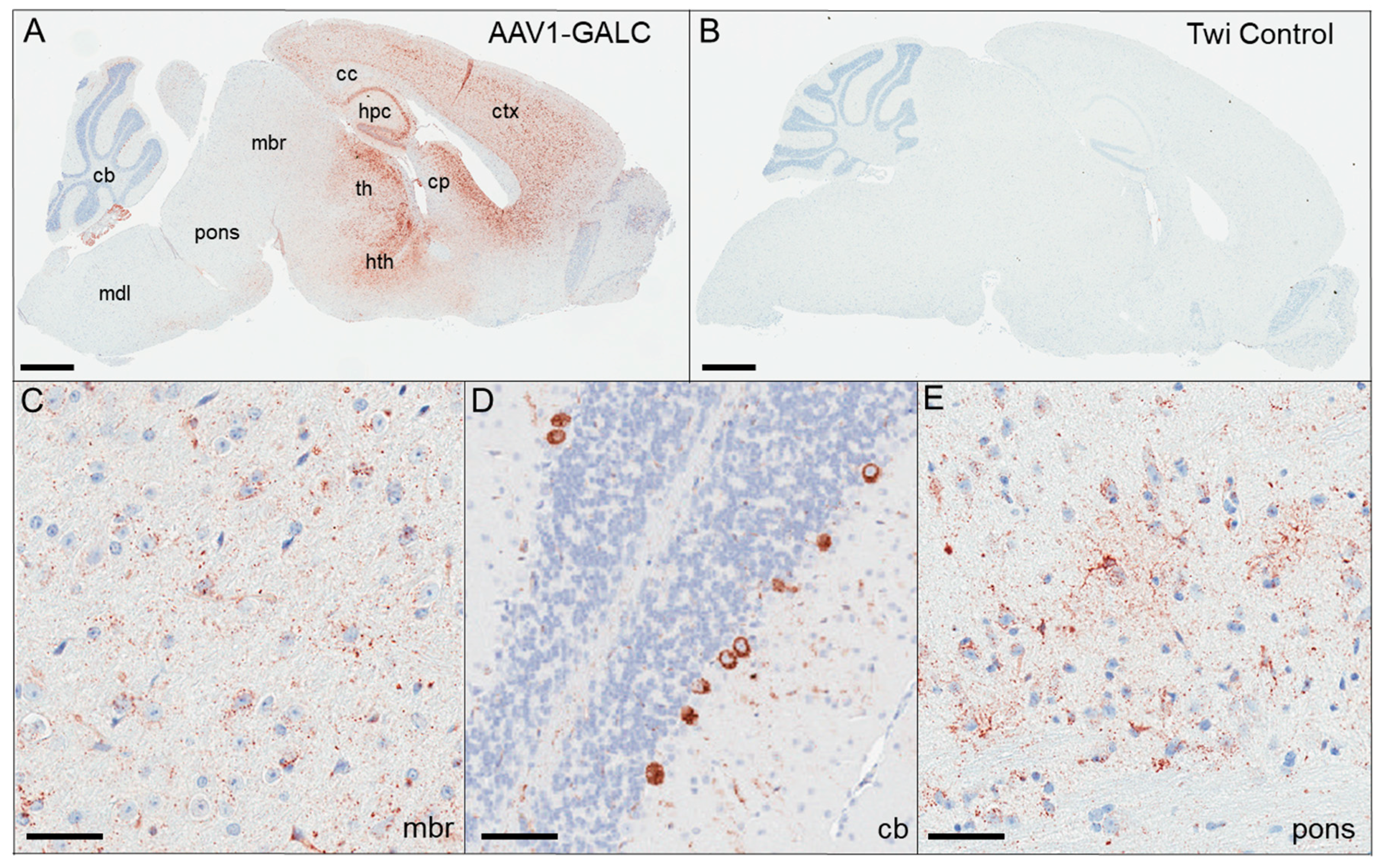{kind=link}
{kind=link}
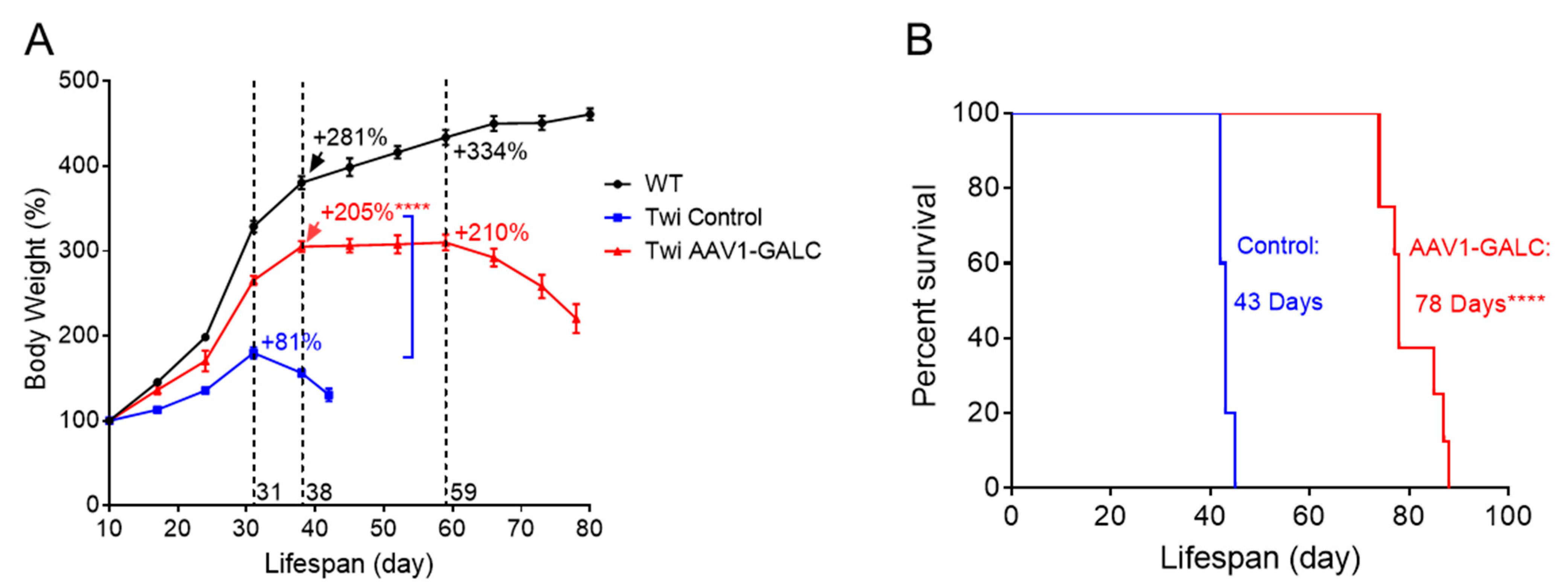{kind=link}
{kind=link}
| ID | Genotype | Exp Group | Treatment Age (PND) | Lifespan (PND) Median (%) | Psychosine (ng/mg) Mean (%) | Max BW (% PND10) | |
|---|---|---|---|---|---|---|---|
| Anterior | Posterior | ||||||
| C01 | Twi | Control | 45 | 6.2 | 13.8 | 179 | |
| C02 | Twi | Control | 42 | 6.0 | 11.5 | 204 | |
| C03 | Twi | Control | 42 | 5.6 | 9.6 | 188 | |
| C04 | Twi | Control | 43 | 4.7 | 11.5 | 165 | |
| C05 | Twi | Control | 43 | 6.3 | 12.3 | 167 | |
| 43 (100%) | 5.8 (100%) | 11.8 (100%) | 181 | ||||
| A01 | Twi | AAV1-GALC | 0 | 78 | 0.9 | 1.6 | 331 |
| A02 | Twi | AAV1-GALC | 0 | 88 | 0.8 | 0.9 | 366 |
| A03 | Twi | AAV1-GALC | 0 | 77 | 0.5 | 2.4 | 304 |
| A04 | Twi | AAV1-GALC | 0 | 87 | 0.6 | 2.0 | 322 |
| A05 | Twi | AAV1-GALC | 3 | 74 | 0.7 | 4.4 | 319 |
| A06 | Twi | AAV1-GALC | 3 | 74 | 0.5 | 4.0 | 306 |
| A07 | Twi | AAV1-GALC | 1 | 78 | 0.7 | 5.5 | 288 |
| A08 | Twi | AAV1-GALC | 1 | 85 | 0.5 | 1.0 | 298 |
| 78 (181%) | 0.6 (11%) | 2.7 (23%) | 317 | ||||
| Controls for psychosine analysis | |||||||
| W01 | WT | Control | 41 | 0.8 | 0.1 | ||
| W02 | WT | Control | 41 | 0.4 | 0.0 | ||
| W03 | WT | Control | 41 | 0.4 | 0.6 | ||
| 0.5 (9%) | 0.3 (2%) | ||||||
| Controls for BW analysis | |||||||
| W04 | WT | Control | 100 | 475 | |||
| W05 | WT | Control | 100 | 495 | |||
| W06 | WT | Control | 100 | 521 | |||
| W07 | WT | Control | 100 | 500 | |||
| 498 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herdt, A.R.; Peng, H.; Dickson, D.W.; Golde, T.E.; Eckman, E.A.; Lee, C.W. Brain Targeted AAV1-GALC Gene Therapy Reduces Psychosine and Extends Lifespan in a Mouse Model of Krabbe Disease. Genes 2023, 14, 1517. https://doi.org/10.3390/genes14081517
Herdt AR, Peng H, Dickson DW, Golde TE, Eckman EA, Lee CW. Brain Targeted AAV1-GALC Gene Therapy Reduces Psychosine and Extends Lifespan in a Mouse Model of Krabbe Disease. Genes. 2023; 14(8):1517. https://doi.org/10.3390/genes14081517
Chicago/Turabian StyleHerdt, Aimee R., Hui Peng, Dennis W. Dickson, Todd E. Golde, Elizabeth A. Eckman, and Chris W. Lee. 2023. "Brain Targeted AAV1-GALC Gene Therapy Reduces Psychosine and Extends Lifespan in a Mouse Model of Krabbe Disease" Genes 14, no. 8: 1517. https://doi.org/10.3390/genes14081517
APA StyleHerdt, A. R., Peng, H., Dickson, D. W., Golde, T. E., Eckman, E. A., & Lee, C. W. (2023). Brain Targeted AAV1-GALC Gene Therapy Reduces Psychosine and Extends Lifespan in a Mouse Model of Krabbe Disease. Genes, 14(8), 1517. https://doi.org/10.3390/genes14081517