
Application of the Yamanaka Transcription Factors Oct4, Sox2, Klf4, and c-Myc from the Laboratory to the Clinic
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
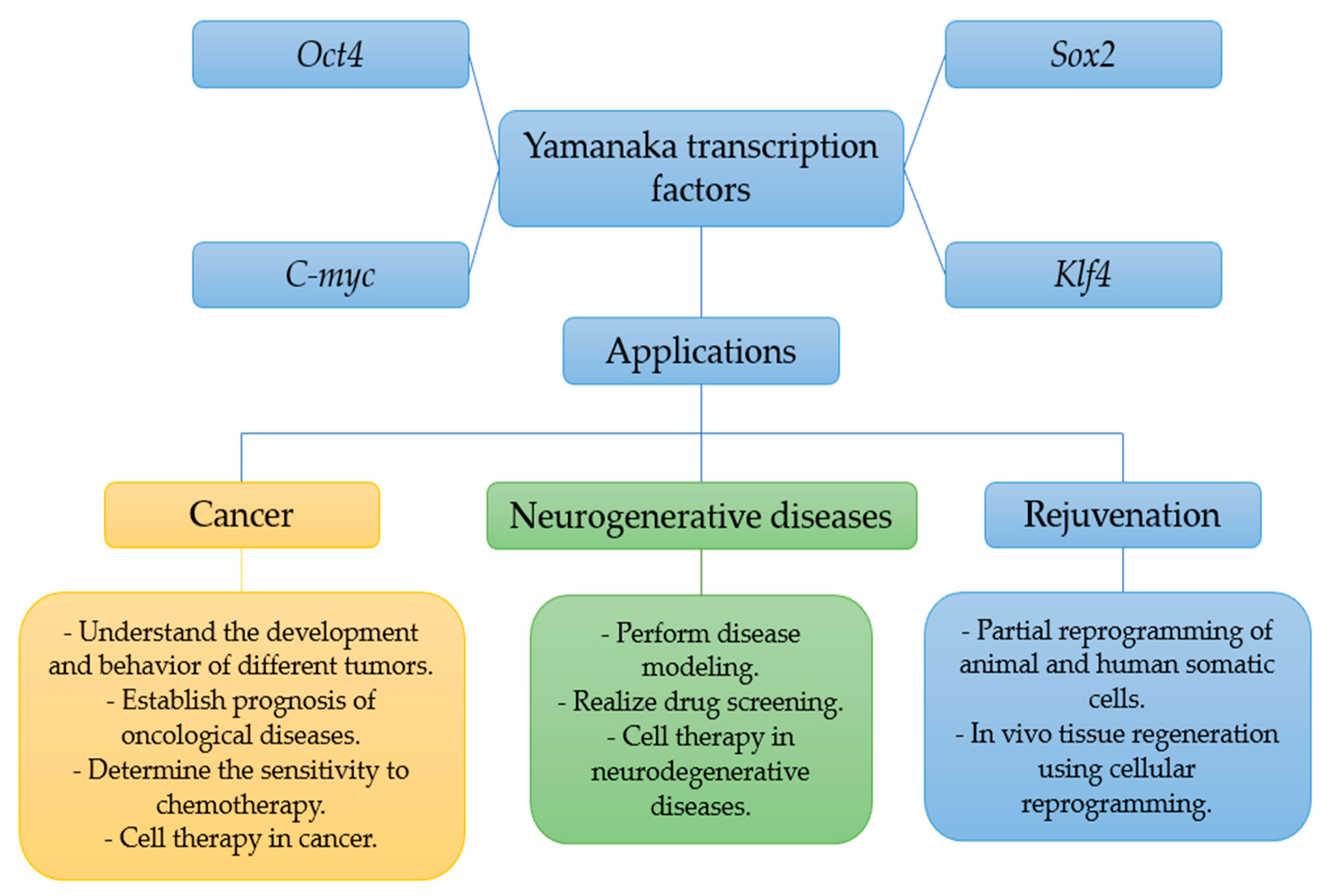
2. Applications of the Yamanaka Transcription Factors in Cancer
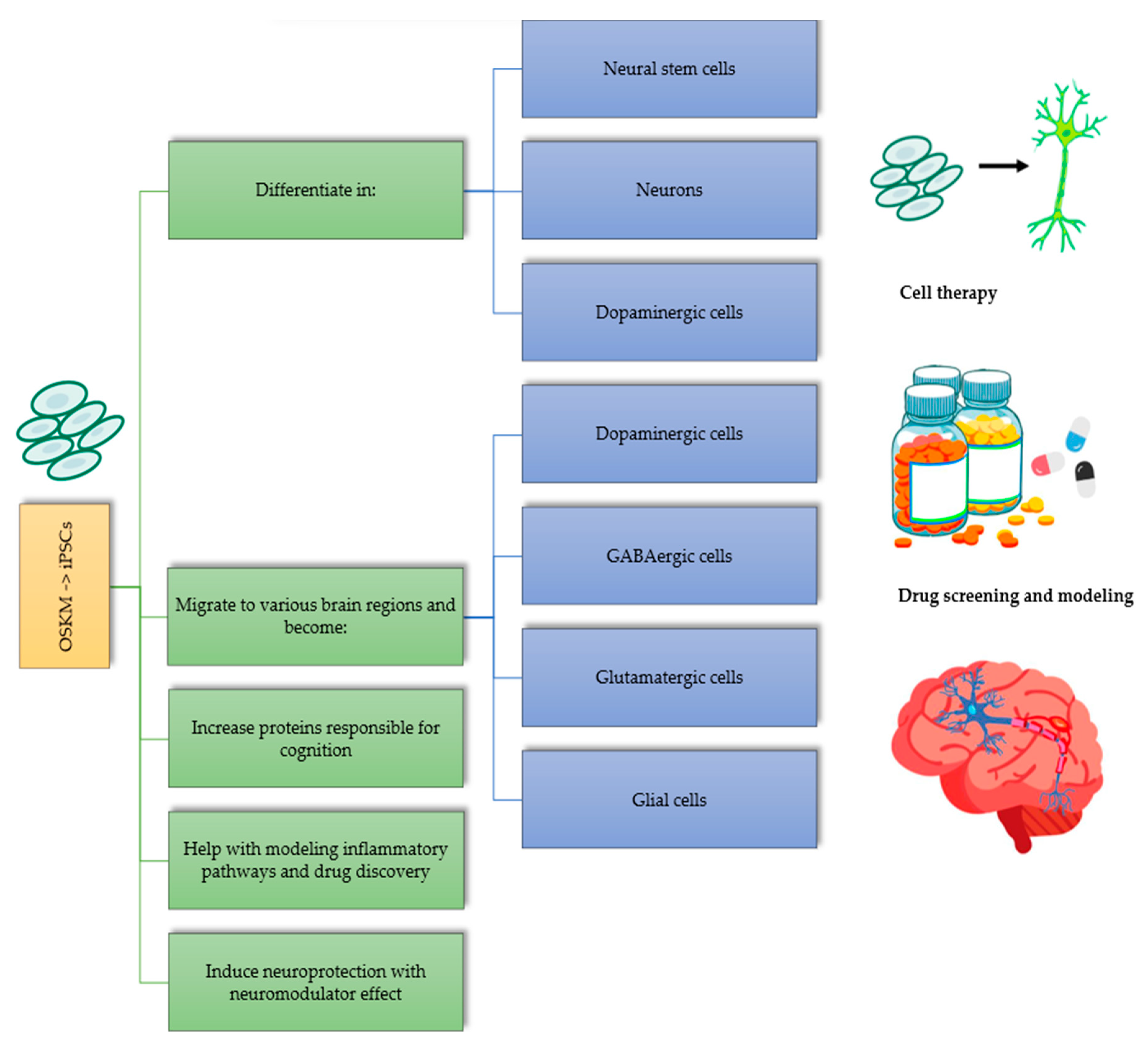
3. Applications of Yamanaka Factors in Neurodegenerative Diseases
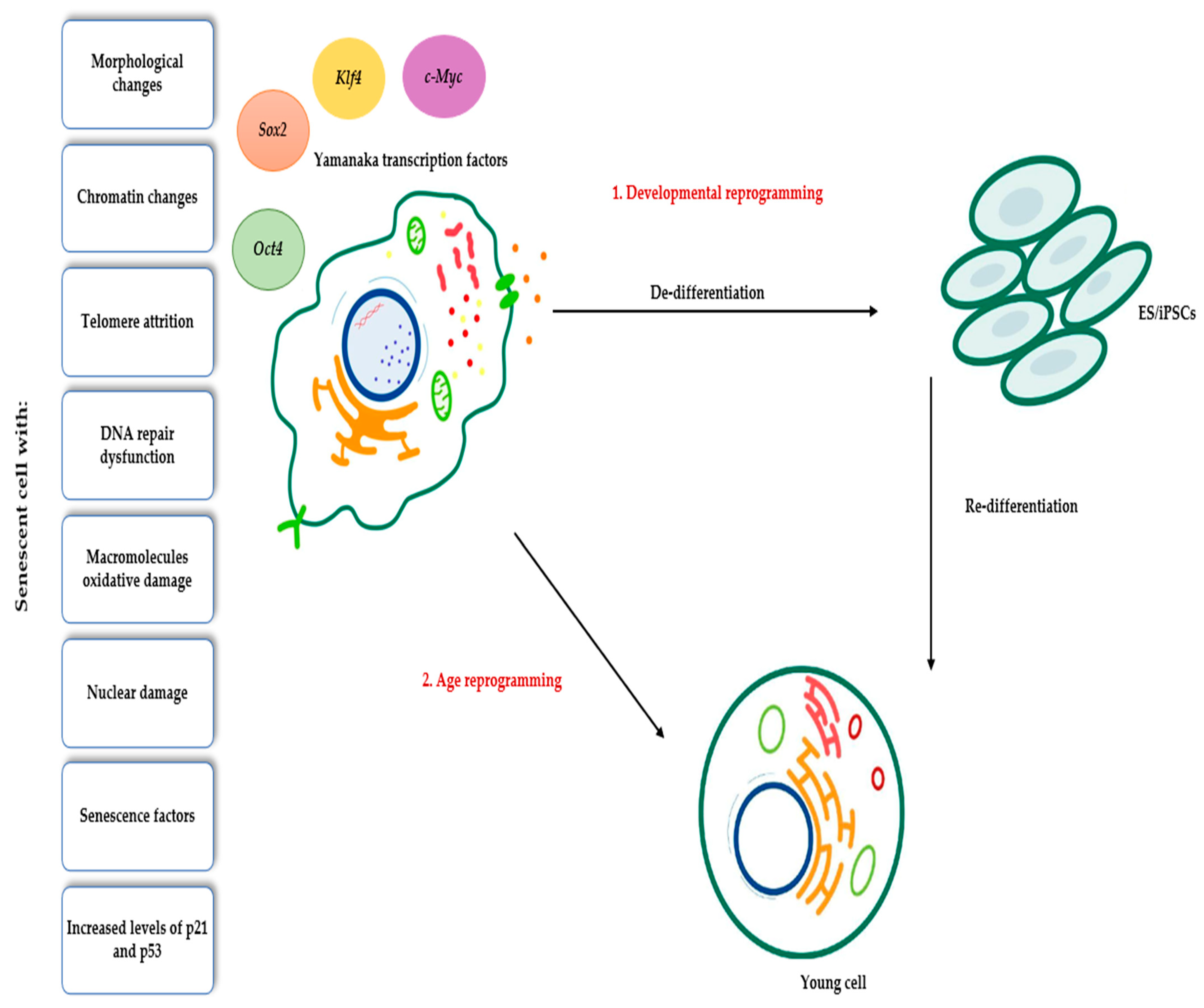
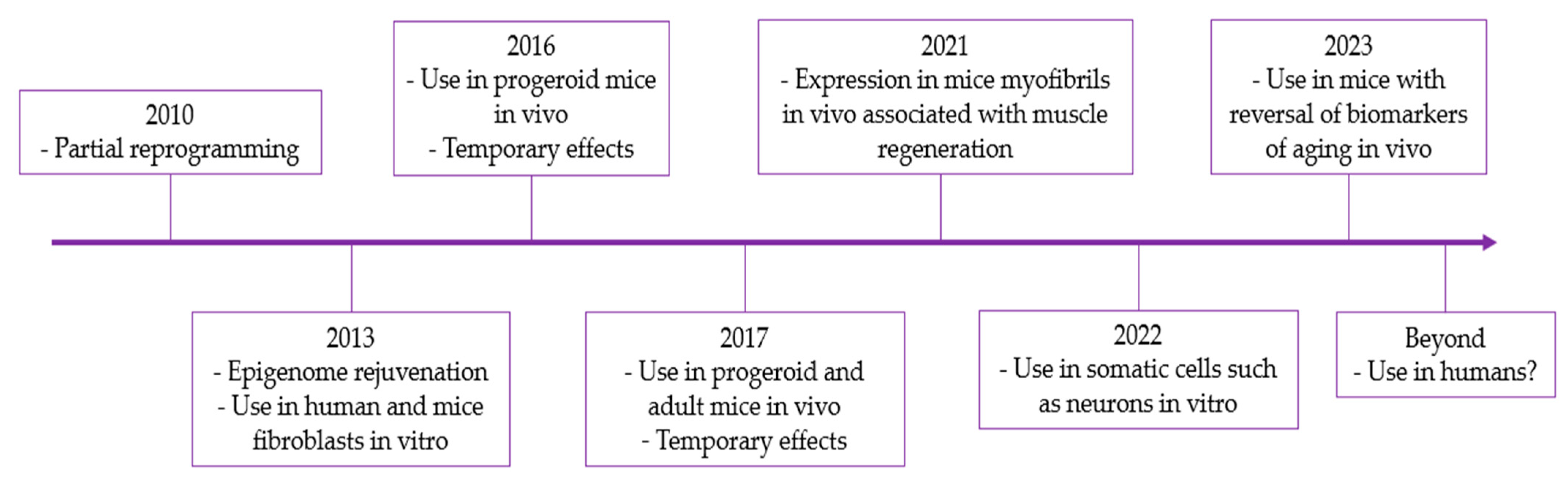
4. Applications of Yamanaka Transcription Factors in Rejuvenation
5. Discussion
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ray, A.; Joshi, J.M.; Sundaravadivelu, P.K.; Raina, K.; Lenka, N.; Kaveeshwar, V.; Thummer, R.P. An overview on promising somatic cell sources utilized for the efficient generation of induced pluripotent stem cells. Stem Cell Rev. Rep. 2021, 17, 1954–1974. [Google Scholar] [CrossRef] [PubMed]
- Stadtfeld, M.; Hochedlinger, K. Induced pluripotency: History, mechanisms, and applications. Genes Dev. 2010, 24, 2239–2263. [Google Scholar] [CrossRef] [PubMed]
- Al Abbar, A.; Ngai, S.C.; Nograles, N.; Alhaji, S.Y.; Abdullah, S. Induced pluripotent stem cells: Reprogramming platforms and applications in cell replacement therapy. Biores. Open Access 2020, 9, 121–136. [Google Scholar] [CrossRef]
- Nichols, J.; Zevnik, B.; Anastassiadis, K.; Niwa, H.; Klewe-Nebenius, D.; Chambers, I.; Schöler, H.; Smith, A. Formation of pluripotent stem cells in the mammalian embryo depends on the POU transcription factor Oct4. Cell 1998, 95, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Niwa, H.; Miyazaki, J.I.; Smith, A.G. Quantitative expression of Oct-3/4 defines differentiation, dedifferentiation or self-renewal of ES cells. Nat. Genet. 2000, 24, 372–376. [Google Scholar] [CrossRef]
- Chaparro, O.; Beltrán, O. Reprogramación nuclear y células pluripotentes inducidas. Rev. Fac. Med. 2009, 17, 252–263. [Google Scholar]
- Ma, Y.; Zhang, X.; Ma, H.; Ren, Y.; Sun, Y.; Wang, Q.; Shi, J. Bioinformatic analysis of the four transcription factors used to induce pluripotent stem cells. Cytotechnology 2014, 66, 967–978. [Google Scholar] [CrossRef]
- Blinka, S.; Rao, S. Nanog Expression in Embryonic Stem Cells—An Ideal Model System to Dissect Enhancer Function. Bioessays 2017, 39, 1700086. [Google Scholar] [CrossRef]
- Welstead, G.G.; Schorderet, P.; Boyer, L.A. The reprogramming language of pluripotency. Curr. Opin. Genet. Dev. 2008, 1, 123–129. [Google Scholar] [CrossRef]
- Huang, K.; Zhang, X.; Shi, J.; Yao, M.; Lin, J.; Li, J.; Liu, H.; Li, H.; Shi, G.; Wang, Z.; et al. Dynamically reorganized chromatin is the key for the reprogramming of somatic cells to pluripotent cells. Sci. Rep. 2015, 7, 17691. [Google Scholar] [CrossRef]
- Madden, S.K.; de Araujo, A.D.; Gerhardt, M.; Fairlie, D.P.; Mason, J.M. Taking the Myc out of cancer: Toward therapeutic strategies to directly inhibit c-Myc. Mol. Cancer 2021, 20, 3. [Google Scholar] [CrossRef] [PubMed]
- Kato, G.J.; Lee, W.M.; Chen, L.L.; Dang, C.V. Max: Functional domains and interaction with c-Myc. Genes Dev. 1992, 6, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Kher, J.D.; Roy, S.P.; Pallavi, K.J.; Gohil, C.J. Review on Induced Pluripotent Stem Cell and its Potential Clinical Applications. J. Anal. Pharm. Res. 2016, 3, 00080. [Google Scholar] [CrossRef]
- Knoepfler, P.S. Why myc? An unexpected ingredient in the stem cell cocktail. Cell Stem Cell. 2008, 2, 18–21. [Google Scholar] [CrossRef] [PubMed]
- Tafani, M.; Perrone, G.A.; Pucci, B.; Russo, A.; Bizzarri, M.; Mechanick, J. Reprogramming Cancer Cells in Endocrine-Related Tumors: Open Issues. Curr. Med. Chem. 2014, 21, 1146–1151. [Google Scholar] [CrossRef] [PubMed]
- Kunisato, A.; Wakatsuki, M.; Kodama, Y.; Shinba, H.; Ishida, I.; Nagao, K. Generation of induced pluripotent stem cells by efficient reprogramming of adult bone marrow cells. Stem Cells Dev. 2010, 19, 229–238. [Google Scholar] [CrossRef]
- Yang, X.; Shao, F.; Guo, D.; Wang, W.; Wang, J.; Zhu, R.; Gao, Y.; He, J.; Lu, Z. WNT/β-catenin-suppressed FTO expression increases mA of c-Myc mRNA to promote tumor cell glycolysis and tumorigenesis. Cell Death Dis. 2021, 12, 462. [Google Scholar] [CrossRef]
- Zhang, J.; Lv, X.; Wei, B.; Gong, X.; Chen, L. CHD4 mediates SOX2 transcription through TRPS1 in luminal breast cancer. Cell. Signal. 2022, 100, 110464. [Google Scholar] [CrossRef]
- Zhang, X.; Cruz, F.; Terry, M.; Remotti, F.; Matushansky, I. Terminal differentiation and loss of tumorigenicity of human cancers via pluripotency based reprogramming. Oncogene 2013, 32, 2249–2260. [Google Scholar] [CrossRef]
- Ai, D.; Yang, X.; Li, X.; Liu, N.; Xie, X.; Liu, Y.; Song, W. Generation of an induced pluripotent stem cell line SDPHi002-A from a patient with lung cancer. Stem Cell Res. 2023, 69, 103096. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, X.; Hu, R.; Pan, G.; Lin, X. SOX2 regulates paclitaxel resistance of A549 non-small cell lung cancer cells via promoting transcription of ClC-3. Oncol. Rep. 2022, 48, 181. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Wang, W.; Chen, B.; Chen, C.; Li, S.; Lu, X. Human induced pluripotent stem cell-derived neurons improve motor asymmetry in a 6-hydroxydopamine-induced rat model of Parkinson’s disease. Cytotherapy 2015, 17, 665–679. [Google Scholar] [CrossRef]
- Luo, Y.; Fan, Y.; Zhou, B.; Xu, Z.; Chen, Y.; Sun, X. Generation of induced pluripotent stem cells from skin fibroblasts of a patient with olivopontocerebellar atrophy. Tohoku J. Exp. Med. 2012, 226, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.W.; Kuang, F.; Wei, L.C.; Ding, Y.X.; Yung, K.K.; Chan, Y.S. Potential application of induced pluripotent stem cells in cell replacement therapy for Parkinson’s disease. CNS Neurol. Disord. Drug Targets 2011, 10, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Du, J.; Fan, J.; Li, W.; Guo, W.; Feng, H.; Lin, J. Generation and Characterization of Induced Pluripotent Stem Cells from Mononuclear Cells in Schizophrenic Patients. Cell J. 2019, 21, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhang, A.; Li, M.; Wang, X.; Wang, X.; Guan, Y.; An, J.; Han, D.; Zhang, Y.A.; Chen, Z. Induced neural stem cells from Macaca fascicularis show potential of dopaminergic neuron specification and efficacy in a mouse Parkinson’s disease model. Acta Histochem. 2022, 124, 151927. [Google Scholar] [CrossRef]
- Feldman, E.L.; Goutman, S.A.; Petri, S.; Mazzini, L.; Savelieff, M.G.; Shaw, P.J.; Sobue, G. Amyotrophic lateral sclerosis. Lancet 2022, 15, 1363–1380. [Google Scholar] [CrossRef] [PubMed]
- Dimos, J.T.; Rodolfa, K.T.; Niakan, K.K.; Weisenthal, L.M.; Mitsumoto, H.; Chung, W.; Croft, G.F.; Saphier, G.; Leibel, R.; Goland, R.; et al. Induced pluripotent stem cells generated from patients with ALS can be differentiated into motor neurons. Science 2008, 321, 1218–1221. [Google Scholar] [CrossRef]
- Ahmadian-Moghadam, H.; Sadat-Shirazi, M.S.; Zarrindast, M.R. Therapeutic potential of stem cells for treatment of neurodegenerative diseases. Biotechnol. Lett. 2020, 42, 1073–1101. [Google Scholar] [CrossRef]
- Du, H.; Huo, Z.; Chen, Y.; Zhao, Z.; Meng, F.; Wang, X.; Liu, S.; Zhang, H.; Zhou, F.; Liu, J.; et al. Induced Pluripotent Stem Cells and Their Applications in Amyotrophic Lateral Sclerosis. Cells 2023, 12, 971. [Google Scholar] [CrossRef]
- Ferraiuolo, L.; Maragakis, N.J. Mini-Review: Induced pluripotent stem cells and the search for new cell-specific ALS therapeutic targets. Neurosci. Lett. 2021, 755, 135911. [Google Scholar] [CrossRef] [PubMed]
- Lamas, N.J.; Roybon, L. Harnessing the Potential of Human Pluripotent Stem Cell-Derived Motor Neurons for Drug Discovery in Amyotrophic Lateral Sclerosis: From the Clinic to the Laboratory and Back to the Patient. Front. Drug Discov. 2021, 1, 773424. [Google Scholar] [CrossRef]
- Lee, S.; Huang, E.J. Modeling ALS and FTD with iPSC-derived neurons. Brain Res. 2017, 1656, 88–97. [Google Scholar] [CrossRef]
- Sivandzade, F.; Cucullo, L. Regenerative Stem Cell Therapy for Neurodegenerative Diseases: An Overview. Int. J. Mol. Sci. 2021, 22, 2153. [Google Scholar] [CrossRef]
- Xu, J.; Fang, S.; Deng, S.; Li, H.; Lin, X.; Huang, Y.; Chung, S.; Shu, Y.; Shao, Z. Generation of neural organoids for spinal-cord regeneration via the direct reprogramming of human astrocytes. Nat. Biomed. Eng. 2023, 7, 253–269. [Google Scholar] [CrossRef]
- Singh, P.B.; Zhakupova, A. Age reprogramming: Cell rejuvenation by partial reprogramming. Development 2022, 149, dev200755. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.B.; Zacouto, F. Nuclear reprogramming and epigenetic rejuvenation. J. Biosci. 2010, 35, 315–319. [Google Scholar] [CrossRef]
- Manukyan, M.; Singh, P.B. Epigenetic rejuvenation. Genes Cells 2012, 17, 337–343. [Google Scholar] [CrossRef]
- Manukyan, M.; Singh, P.B. Epigenome rejuvenation: HP1β mobility as a measure of pluripotent and senescent chromatin ground states. Sci. Rep. 2014, 4, 4789. [Google Scholar] [CrossRef]
- Hamsanathan, S.; Gurkar, A.U. Lipids as Regulators of Cellular Senescence. Front. Physiol. 2022, 13, 796850. [Google Scholar] [CrossRef]
- Zhang, W.; Qu, J.; Liu, G.H.; Belmonte, J.C.I. The ageing epigenome and its rejuvenation. Nat. Rev. Mol. Cell. Biol. 2020, 21, 137–150. [Google Scholar] [CrossRef]
- De Lima Camillo, L.P.; Quinlan, R.B. A ride through the epigenetic landscape: Aging reversal by reprogramming. Geroscience 2021, 43, 463–485. [Google Scholar] [CrossRef] [PubMed]
- Ocampo, A.; Reddy, P.; Martinez-Redondo, P.; Platero-Luengo, A.; Hatanaka, F.; Hishida, T.; Li, M.; Lam, D.; Kurita, M.; Beyret, E.; et al. In Vivo Amelioration of Age-Associated Hallmarks by Partial Reprogramming. Cell 2016, 167, 1719–1733. [Google Scholar] [CrossRef] [PubMed]
- Horvath, S.; Raj, K. DNA methylation-based biomarkers and the epigenetic clock theory of ageing. Nat. Rev. Genet. 2018, 19, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Olova, N.; Simpson, D.J.; Marioni, R.E.; Chandra, T. Partial reprogramming induces a steady decline in epigenetic age before loss of somatic identity. Aging Cell 2018, 18, e12877. [Google Scholar] [CrossRef]
- Mendelsohn, A.R.; Larrick, J.W.; Lei, J.L. Rejuvenation by Partial Reprogramming of the Epigenome. Rejuvenation Res. 2017, 20, 146–150. [Google Scholar] [CrossRef]
- Yang, S.G.; Wang, X.W.; Qian, C.; Zhou, F.Q. Reprogramming neurons for regeneration: The fountain of youth. Prog. Neurobiol. 2022, 214, 102284. [Google Scholar] [CrossRef]
- Wang, C.; Rabadan Ros, R.; Martinez-Redondo, P.; Ma, Z.; Shi, L.; Xue, Y.; Guillen-Guillen, I.; Huang, L.; Hishida, T.; Liao, H.K.; et al. In vivo partial reprogramming of myofibers promotes muscle regeneration by remodeling the stem cell niche. Nat. Commun. 2021, 12, 3094. [Google Scholar] [CrossRef]
- Macip, C.C.; Hasan, R.; Hoznek, V.; Kim, J.; IV, L.E.M.; Sethna, S.; Davidsohn, N. Gene Therapy Mediated Partial Reprogramming Extends Lifespan and Reverses Age- Related Changes in Aged Mice. bioRxiv 2023, 1–22. [Google Scholar] [CrossRef]
- Pessina, P.; Di Stefano, B. Early Life Reprogramming-Based Treatment Promotes Longevity. Cell. Reprogram. 2023, 25, 9–10. [Google Scholar] [CrossRef]
- Simpson, D.J.; Olova, N.N.; Chandra, T. Cellular reprogramming and epigenetic rejuvenation. Clin. Epigen. 2021, 13, 170. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef]
- Nakagawa, M.; Koyanag, M.; Tanabe, K.; Takahashi, K.; Ichisaka, T.; Aoi, T.; Okita, K.; Mochiduki, Y.; Takizawa, N.; Yamanaka, S. Generation of induced pluripotent stem cells without Myc from mouse and human fibroblasts. Nat. Biotechnol. 2008, 26, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Freed, C.R. Adenoviral gene delivery can reprogram human fibroblasts to induced pluripotent stem cells. Stem Cells 2009, 27, 2667–2674. [Google Scholar] [CrossRef] [PubMed]
- Haase, A.; Olmer, R.; Schwanke, K.; Wunderlich, S.; Merkert, S.; Hess, C.; Zweigerdt, R.; Gruh, I.; Meyer, J.; Wagner, S.; et al. Generation of induced pluripotent stem cells from human cord blood. Cell Stem Cell 2009, 5, 434–441. [Google Scholar] [CrossRef]
- Kim, D.; Kim, C.H.; Moon, J.I.; Chung, Y.G.; Chang, M.Y.; Han, B.S.; Ko, S.; Yang, E.; Cha, K.Y.; Lanza, R.; et al. Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins. Cell Stem Cell 2009, 4, 472–476. [Google Scholar] [CrossRef]
- Fusaki, N.; Ban, H.; Nishiyama, A.; Saeki, K.; Hasegawa, M. Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai virus, an RNA virus that does not integrate into the host genome. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2009, 85, 348–362. [Google Scholar] [CrossRef]
- Barrero, M.J.; Boué, S.; Izpisúa Belmonte, J.C. Epigenetic mechanisms that regulate cell identity. Cell Stem Cell 2010, 7, 565–570. [Google Scholar] [CrossRef]
- Nicetto, D.; Zaret, K.S. Role of H3K9me3 heterochromatin in cell identity establishment and maintenance. Curr. Opin. Genet. Dev. 2019, 55, 1–10. [Google Scholar] [CrossRef]
- Hawkins, R.D.; Hon, G.C.; Lee, L.K.; Ngo, Q.; Lister, R.; Pelizzola, M.; Edsall, L.E.; Kuan, S.; Luu, Y.; Klugman, S.; et al. Distinct epigenomic landscapes of pluripotent and lineage-committed human cells. Cell Stem Cell 2010, 6, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Sánchez Alvarado, A.; Tsonis, P.A. Bridging the regeneration gap: Genetic insights from diverse animal models. Nat. Rev. Genet. 2006, 7, 873–884. [Google Scholar] [CrossRef]
- Wang, W.; Hu, C.K.; Zeng, A.; Alegre, D.; Hu, D.; Gotting, K.; Ortega Granillo, A.; Wang, Y.; Robb, S.; Schnittker, R.; et al. Changes in regeneration-responsive enhancers shape regenerative capacities in vertebrates. Science 2020, 369, eaaz3090. [Google Scholar] [CrossRef]
- Guan, J.; Wang, G.; Wang, J.; Zhang, Z.; Fu, Y.; Cheng, L.; Meng, G.; Lyu, Y.; Zhu, J.; Li, Y.; et al. Chemical reprogramming of human somatic cells to pluripotent stem cells. Nature 2022, 605, 325–331. [Google Scholar] [CrossRef] [PubMed]
- De Magalhães, J.P.; Ocampo, A. Cellular reprogramming and the rise of rejuvenation biotech. Trends Biotechnol. 2022, 40, 639–642. [Google Scholar] [CrossRef] [PubMed]
- Damdimopoulou, P.; Rodin, S.; Stenfelt, S.; Antonsson, L.; Tryggvason, K.; Hovatta, O. Human embryonic stem cells. Best Pract. Res. Clin. Obstet. Gynaecol. 2016, 31, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Bai, Q.; Desprat, R.; Klein, B.; Lemaître, J.M.; De Vos, J. Embryonic stem cells or induced pluripotent stem cells? A DNA integrity perspective. Curr. Gene Ther. 2013, 13, 93–98. [Google Scholar] [CrossRef]
- Yamanaka, S. Pluripotent Stem Cell-Based Cell Therapy-Promise and Challenges. Cell Stem Cell 2020, 27, 523–531. [Google Scholar] [CrossRef]
- Ohnuki, M.; Takahashi, K. Present and future challenges of induced pluripotent stem cells. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140367. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguirre, M.; Escobar, M.; Forero Amézquita, S.; Cubillos, D.; Rincón, C.; Vanegas, P.; Tarazona, M.P.; Atuesta Escobar, S.; Blanco, J.C.; Celis, L.G. Application of the Yamanaka Transcription Factors Oct4, Sox2, Klf4, and c-Myc from the Laboratory to the Clinic. Genes 2023, 14, 1697. https://doi.org/10.3390/genes14091697
Aguirre M, Escobar M, Forero Amézquita S, Cubillos D, Rincón C, Vanegas P, Tarazona MP, Atuesta Escobar S, Blanco JC, Celis LG. Application of the Yamanaka Transcription Factors Oct4, Sox2, Klf4, and c-Myc from the Laboratory to the Clinic. Genes. 2023; 14(9):1697. https://doi.org/10.3390/genes14091697
Chicago/Turabian StyleAguirre, Marisol, Manuela Escobar, Sebastián Forero Amézquita, David Cubillos, Camilo Rincón, Paula Vanegas, María Paula Tarazona, Sofía Atuesta Escobar, Juan Camilo Blanco, and Luis Gustavo Celis. 2023. "Application of the Yamanaka Transcription Factors Oct4, Sox2, Klf4, and c-Myc from the Laboratory to the Clinic" Genes 14, no. 9: 1697. https://doi.org/10.3390/genes14091697
APA StyleAguirre, M., Escobar, M., Forero Amézquita, S., Cubillos, D., Rincón, C., Vanegas, P., Tarazona, M. P., Atuesta Escobar, S., Blanco, J. C., & Celis, L. G. (2023). Application of the Yamanaka Transcription Factors Oct4, Sox2, Klf4, and c-Myc from the Laboratory to the Clinic. Genes, 14(9), 1697. https://doi.org/10.3390/genes14091697