
Phylogenetic Analysis of Russian Native Sheep Breeds Based on mtDNA Sequences

 ,
,  ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Bioinformatics Data Processing
3. Results
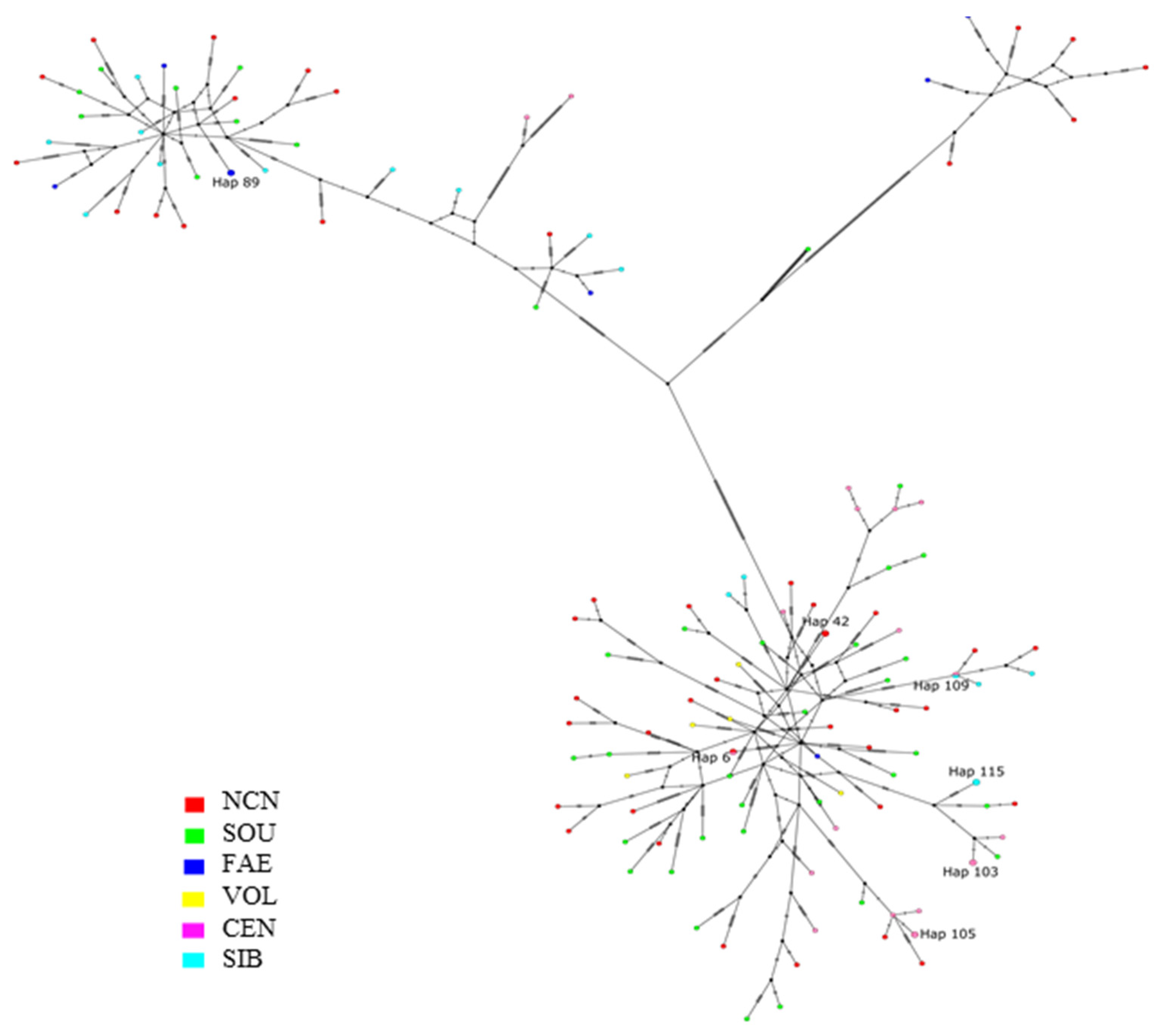
3.1. Genetic Diversity and Haplotype Network
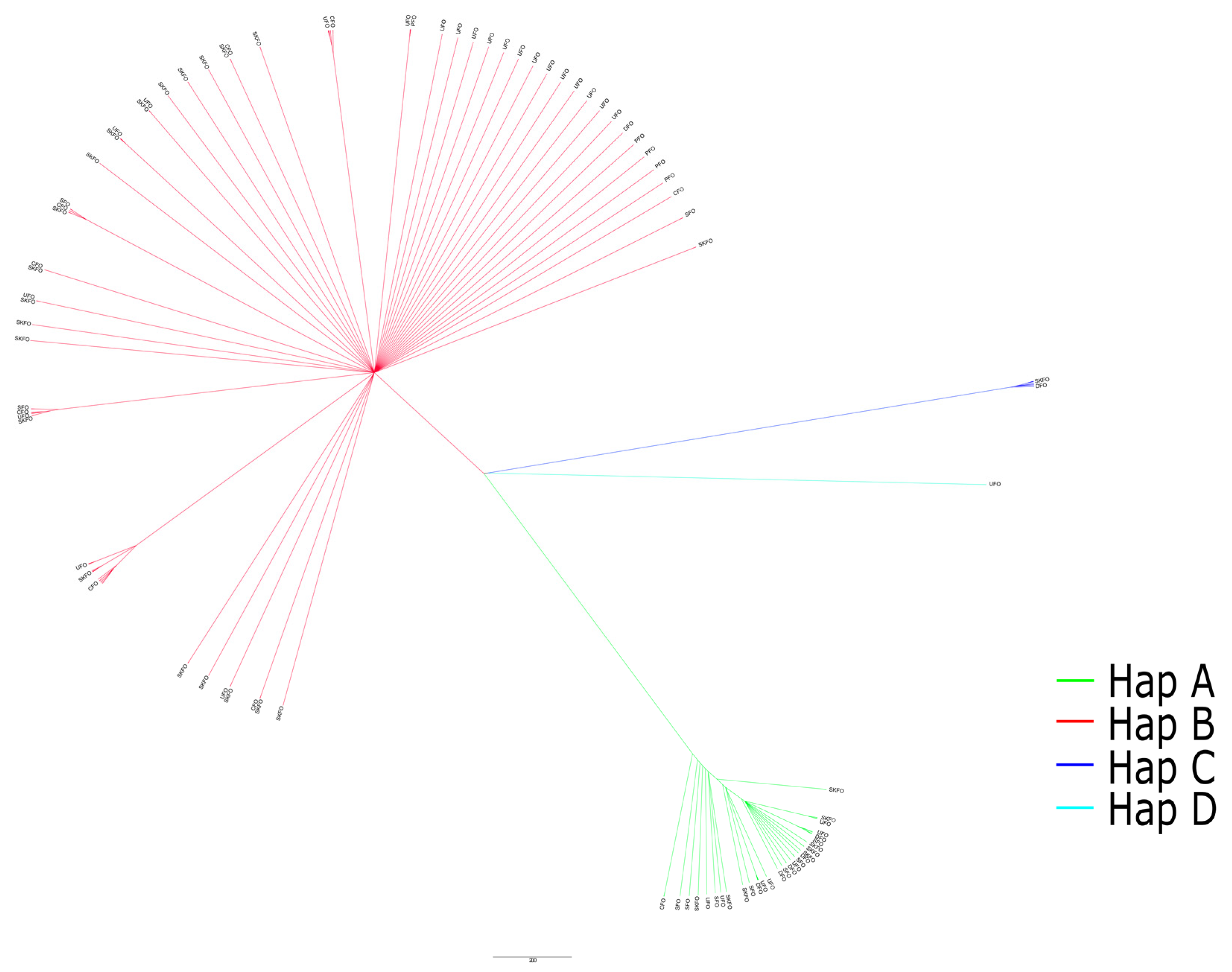
3.2. Haplogroup Assignment
3.3. Genetic Differentiation between Russian Sheep from Diverse Geographical Regions
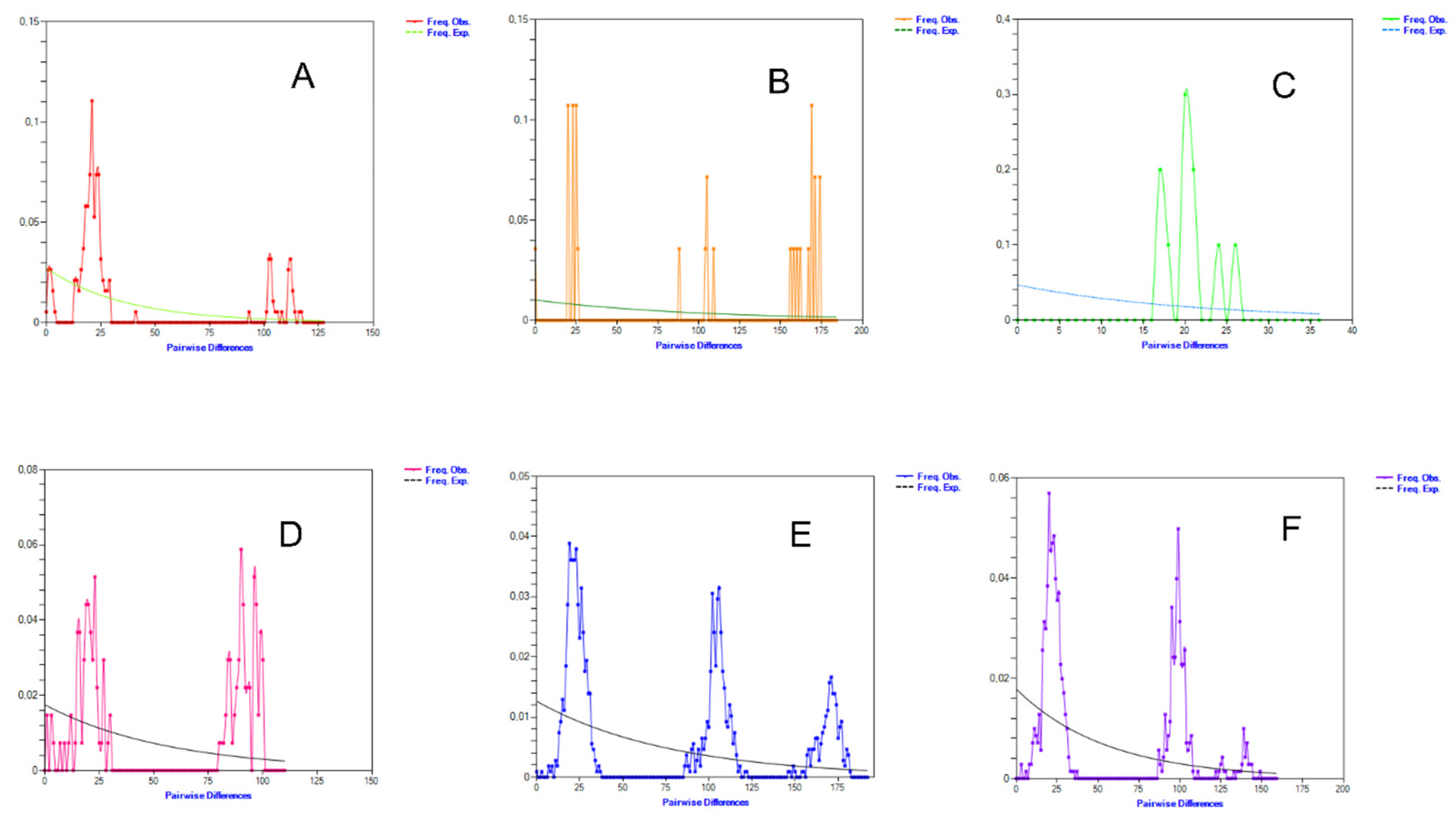
3.4. Population Dynamics and Mismatch Distribution of Russian Sheep from Different Regions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chessa, B.; Pereira, F.; Arnaud, F.; Amorim, A.; Goyache, F.; Mainland, I.; Kao, R.R.; Pemberton, J.M.; Beraldi, D.; Stear, M.J.; et al. Revealing the history of sheep domestication using retrovirus integrations. Science 2009, 324, 532–536. [Google Scholar] [CrossRef] [PubMed]
- Luigi-Sierra, M.G.; Mármol-Sánchez, E.; Amills, M. Comparing the diversity of the casein genes in the Asian mouflon and domestic sheep. Anim. Genet. 2020, 51, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Zeder, M.A. Domestication and early agriculture in the Mediterranean Basin: Origins, diffusion, and impact. Proc. Natl. Acad. Sci. USA 2008, 105, 11597–11604. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). Available online: https://www.fao.org/livestock-systems/global-distributions/sheep/en/. (accessed on 21 August 2023).
- Chen, Z.H.; Xu, Y.X.; Xie, X.L.; Wang, D.F.; Aguilar-Gómez, D.; Liu, G.J.; Li, X.; Esmailizadeh, A.; Rezaei, V.; Kantanen, J.; et al. Whole-genome sequence analysis unveils different origins of European and Asiatic mouflon and domestication-related genes in sheep. Commun. Biol. 2021, 4, 1307. [Google Scholar] [CrossRef] [PubMed]
- Meadows, J.R.; Cemal, I.; Karaca, O.; Gootwine, E.; Kijas, J.W. Five ovine mitochondrial lineages identified from sheep breeds of the near East. Genetics 2007, 175, 1371–1379. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, C.; Ermini, L.; Rizzi, E.; Corti, G.; Luciani, S.; Marota, I.; De Bellis, G.; Rollo, F. Phylogenetic position of a copper age sheep (Ovis aries) mitochondrial DNA. PLoS ONE 2012, 7, e33792. [Google Scholar] [CrossRef]
- Peters, J.; von den Driesch, A.; Helmer, D. The upper Euphrates-Tigris Basin: Cradle of agro-pastoralism? In The First Steps of Animal Domestication. New Archaeological Approaches; Vigne, J.D., Peters, J., Helmer, D., Eds.; Oxbow Books: Oxford, UK, 2005; pp. 96–124. [Google Scholar]
- Molaee, V.; Bazzucchi, M.; De Mia, G.M.; Otarod, V.; Abdollahi, D.; Rosati, S.; Lühken, G. Phylogenetic analysis of small ruminant lentiviruses in Germany and Iran suggests their expansion with domestic sheep. Sci. Rep. 2020, 10, 2243. [Google Scholar] [CrossRef]
- Larsson, M.N.A.; Miranda, P.M.; Pan, L.; Başak Vural, K.; Kaptan, D.; Rodrigues Soares, A.E.; Kivikero, H.; Kantanen, J.; Somel, M.; Özer, F.; et al. Ancient sheep genomes reveal four Millennia of North European short-tailed sheep in the Baltic Sea region. bioRxiv 2023, 544912. [Google Scholar] [CrossRef]
- Zeder, M.A. Domestication as a model system for the extended evolutionary synthesis. Interface Focus 2017, 7, 20160133. [Google Scholar] [CrossRef]
- Tapio, M.; Marzanov, N.; Ozerov, M.; Cinkulov, M.; Gonzarenko, G.; Kiselyova, T.; Murawski, M.; Viinalass, H.; Kantanen, J. Sheep mitochondrial DNA variation in European, Caucasian, and Central Asian areas. Mol. Biol. Evol. 2006, 23, 1776–1783. [Google Scholar] [CrossRef]
- Niemi, M.; Bläuer, A.; Iso-Touru, T.; Nyström, V.; Harjula, J.; Taavitsainen, J.P.; Storå, J.; Lidén, K.; Kantanen, J. Mitochondrial DNA and Y-chromosomal diversity in ancient populations of domestic sheep (Ovis aries) in Finland: Comparison with contemporary sheep breeds. Genet. Sel. Evol. 2013, 45, 2. [Google Scholar] [CrossRef] [PubMed]
- Lv, F.H.; Peng, W.F.; Yang, J.; Zhao, Y.X.; Li, W.R.; Liu, M.J.; Ma, Y.H.; Zhao, Q.J.; Yang, G.L.; Wang, F.; et al. Mitogenomic meta-analysis identifies two phases of migration in the history of Eastern Eurasian sheep. Mol. Biol. Evol. 2015, 32, 2515–2533. [Google Scholar] [CrossRef] [PubMed]
- Ganbold, O.; Lee, S.H.; Paek, W.K.; Munkhbayar, M.; Seo, D.; Manjula, P.; Khujuu, T.; Purevee, E.; Lee, J.H. Mitochondrial DNA variation and phylogeography of native Mongolian goats. Asian Australas J. Anim. Sci. 2020, 33, 902–912. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Kumar, S., Jr.; Kolte, A.P.; Kumar, S. Extensive variation and sub-structuring in lineage A mtDNA in Indian sheep: Genetic evidence for domestication of sheep in India. PLoS ONE 2013, 8, e77858. [Google Scholar] [CrossRef]
- Ministry of Agriculture of the Russian Federation. Open Data. Available online: http://opendata.mcx.ru/opendata. (accessed on 17 April 2023).
- Zhong, Y.; Tang, Z.; Huang, L.; Wang, D.; Lu, Z. Genetic diversity of Procambarus clarkii populations based on mitochondrial DNA and microsatellite markers in different areas of Guangxi, China. Mitochondrial DNA Part A 2020, 31, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Scherf, B.D. World Watch List for Domestic Animal Diversity, 3rd ed.; FAO: Rome, Italy, 2000; pp. 7–13. [Google Scholar]
- Ramljak, J.; Bunevski, G.; Bytyqi, H.; Marković, B.; Brka, M.; Ivanković, A.; Kume, K.; Stojanović, S.; Nikolov, V.; Simčič, M.; et al. Conservation of a domestic metapopulation structured into related and partly admixed strains. Mol. Ecol. 2018, 27, 1633–1650. [Google Scholar] [CrossRef]
- Mishra, A.K.; Ahlawat, S.; Sharma, R.; Arora, R.; Singh, S.; Jain, A. Assessment of genetic diversity of the fat-tailed Dumba sheep of India by mitochondrial and microsatellite markers. Anim. Biotechnol. 2023; advance online publication. [Google Scholar]
- Salim, B.; Alasmari, S.; Mohamed, N.S.; Ahmed, M.A.; Nakao, R.; Hanotte, O. Genetic variation and demographic history of Sudan desert sheep reveal two diversified lineages. BMC Genom. 2023, 24, 118. [Google Scholar] [CrossRef]
- Al-Araimi, N.A.; Al-Atiyat, R.M.; Gaafar, O.M.; Vasconcelos, R.; Luzuriaga-Neira, A.; Eisa, M.O.; Amir, N.; Benaissa, M.H.; Alfaris, A.A.; Aljumaah, R.S.; et al. Maternal genetic diversity and phylogeography of native Arabian goats. Livest. Sci. 2017, 206, 88–94. [Google Scholar] [CrossRef]
- Ibrahim, A.; Baliarti, E.; Budisatria, I.G.S.; Artama, W.T.; Widayanti, R.; Maharani, D.; Tavares, L.; Margawati, E.T. Genetic diversity and relationship among Indonesian local sheep breeds on Java Island based on mitochondrial cytochrome b gene sequences. J. Genet. Eng. Biotechnol. 2023, 21, 34. [Google Scholar] [CrossRef]
- Dymova, M.A.; Zadorozhny, A.V.; Mishukova, O.V.; Khrapov, E.A.; Druzhkova, A.S.; Trifonov, V.A.; Kichigin, I.G.; Tishkin, A.A.; Grushin, S.P.; Filipenko, M.L. Mitochondrial DNA analysis of ancient sheep from Altai. Anim. Genet. 2017, 48, 615–618. [Google Scholar] [CrossRef]
- Kamalakkannan, R.; Kumar, S.; Bhavana, K.; Prabhu, V.R.; Machado, C.B.; Singha, H.S.; Sureshgopi, D.; Vijay, V.; Nagarajan, M. Evidence for independent domestication of sheep mtDNA lineage A in India and introduction of lineage B through Arabian sea route. Sci. Rep. 2021, 11, 19733. [Google Scholar] [CrossRef] [PubMed]
- Mukhametzharova, I.; Islamov, Y.; Shauyenov, S.; Ibrayev, D.; Atavliyeva, S.; Tarlykov, P. Genetic characterization of Kazakh native sheep breeds using mitochondrial DNA. J. Biol. Sci. 2018, 18, 341–348. [Google Scholar] [CrossRef]
- Wood, N.J.; Phua, S.H. Variation in the control region sequence of the sheep mitochondrial genome. Anim. Genet. 1996, 27, 25–33. [Google Scholar] [CrossRef]
- Hiendleder, S.; Mainz, K.; Plante, Y.; Lewalski, H. Analysis of mitochondrial DNA indicates that domestic sheep are derived from two different ancestral maternal sources: No evidence for contributions from urial and argali sheep. J. Hered. 1998, 89, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Du, L.X.; Ma, Y.H.; Guan, W.J.; Li, H.B.; Zhao, Q.J.; Li, X.; Rao, S.Q. A novel maternal lineage revealed in sheep (Ovis aries). Anim. Genet. 2005, 36, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Pedrosa, S.; Uzun, M.; Arranz, J.J.; Gutiérrez-Gil, B.; San Primitivo, F.; Bayón, Y. Evidence of three maternal lineages in Near Eastern sheep supporting multiple domestication events. Proc. Biol. Sci. 2005, 272, 2211–2217. [Google Scholar] [CrossRef] [PubMed]
- Pereira, F.; Davis, S.J.; Pereira, L.; McEvoy, B.; Bradley, D.G.; Amorim, A. Genetic signatures of a Mediterranean influence in Iberian Peninsula sheep husbandry. Mol. Biol. Evol. 2006, 23, 1420–1426. [Google Scholar] [CrossRef] [PubMed]
- Sanna, D.; Barbato, M.; Hadjisterkotis, E.; Cossu, P.; Decandia, L.; Trova, S.; Pirastru, M.; Leoni, G.G.; Naitana, S.; Francalacci, P.; et al. The first mitogenome of the Cyprus Mouflon (Ovis gmelini ophion): New insights into the phylogeny of the genus Ovis. PLoS ONE 2015, 10, e0144257. [Google Scholar] [CrossRef]
- Demirci, S.; Koban Baştanlar, E.; Dağtaş, N.D.; Pişkin, E.; Engin, A.; Ozer, F.; Yüncü, E.; Doğan, S.A.; Togan, I. Mitochondrial DNA diversity of modern, ancient and wild sheep (Ovis gmelinii anatolica) from Turkey: New insights on the evolutionary history of sheep. PLoS ONE 2013, 8, e81952. [Google Scholar] [CrossRef]
- Mariotti, M.; Valentini, A.; Marsan, P.A.; Pariset, L. Mitochondrial DNA of seven Italian sheep breeds shows faint signatures of domestication and suggests recent breed formation. Mitochondrial DNA 2013, 24, 577–583. [Google Scholar] [CrossRef]
- Gáspárdy, A.; Berger, B.; Zabavnik-Piano, J.; Kovács, E.; Annus, K.; Zenke, P.; Sáfár, L.; Maróti-Agóts, Á. Comparison of mtDNA control region among descendant breeds of the extinct Zaupel sheep revealed haplogroup C and D in Central Europe. Vet. Med. Sci. 2021, 7, 2330–2338. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ding, X.; Zeng, Y.; Yue, Y.; Guo, X.; Guo, T.; Chu, M.; Wang, F.; Han, J.; Feng, R.; et al. Genetic diversity and phylogenetic evolution of Tibetan sheep based on mtDNA D-Loop sequences. PLoS ONE 2016, 11, e0159308. [Google Scholar]
- Rafia, P.; Tarang, A. Sequence variations of mitochondrial DNA displacement-loop in Iranian indigenous sheep breeds. J. Appl. Anim. Sci. 2016, 6, 363–368. [Google Scholar]
- Rezaei, H.R.; Naderi, S.; Chintauan-Marquier, I.C.; Jordan, S.; Taberlet, P.; Virk, A.T.; Naghash, H.R.; Rioux, D.; Kaboli, M.; Luikart, G.; et al. Evolution and taxonomy of the wild species of the genus Ovis (Mammalia, Artiodactyla, Bovidae). Mol. Phylogenet. Evol. 2010, 54, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Barbato, M.; Masseti, M.; Pirastru, M.; Columbano, N.; Scali, M.; Vignani, R.; Mereu, P. Islands as time capsules for genetic diversity conservation: The case of the Giglio Island mouflon. Diversity 2022, 14, 609. [Google Scholar] [CrossRef]
- Liu, J.; Lu, Z.; Yuan, C.; Wang, F.; Yang, B. Phylogeography and phylogenetic evolution in Tibetan sheep based on mt-CyB sequences. Animals 2020, 10, 1177. [Google Scholar] [CrossRef]
- Kalinowski, S.T.; Wagner, A.P.; Taper, M.L. ML-Relate: A computer program for maximum likelihood estimation of relatedness and relationship. Mol. Ecol. Notes. 2006, 6, 576–579. [Google Scholar] [CrossRef]
- Basic Local Alignment Search Tool (BLAST). Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 10 January 2023).
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Peng, M.S.; Fan, L.; Shi, N.N.; Ning, T.; Yao, Y.G.; Murphy, R.W.; Wang, W.; Zhang, Y.P. DomeTree: A canonical toolkit for mitochondrial DNA analyses in domesticated animals. Mol. Ecol. Resour. 2015, 15, 1238–1242. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Molecular Evolution, Phylogenetics and Epidemiology. Available online: http://tree.bio.ed.ac.uk/soft-ware/figtree. (accessed on 1 April 2023).
- Leigh, J.W.; Bryant, D. Popart: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Wright, S. Evolution in Mendelian populations. Genetics 1931, 16, 97–159. [Google Scholar] [CrossRef]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Resende, A.; Gonçalves, J.; Muigai, A.W.; Pereira, F. Mitochondrial DNA variation of domestic sheep (Ovis aries) in Kenya. Anim. Genet. 2016, 47, 377–381. [Google Scholar] [CrossRef]
- Sharma, R.; Ahlawat, S.; Sharma, H.; Sharma, P.; Panchal, P.; Arora, R.; Tantia, M.S. Microsatellite and mitochondrial DNA analyses unveil the genetic structure of native sheep breeds from three major agro-ecological regions of India. Sci. Rep. 2020, 10, 20422. [Google Scholar] [CrossRef]
- Taylor, W.T.T.; Pruvost, M.; Posth, C.; Rendu, W.; Krajcarz, M.T.; Abdykanova, A.; Brancaleoni, G.; Spengler, R.; Hermes, T.; Schiavinato, S.; et al. Evidence for early dispersal of domestic sheep into Central Asia. Nat. Hum. Behav. 2021, 5, 1169–1179. [Google Scholar] [CrossRef]
- Machová, K.; Málková, A.; Vostrý, L. Sheep post-domestication expansion in the context of mitochondrial and Y chromosome haplogroups and haplotypes. Genes 2022, 13, 613. [Google Scholar] [CrossRef] [PubMed]
- Wanjala, G.; Bagi, Z.; Kusza, S. Meta-analysis of mitochondrial DNA control region diversity to shed light on phylogenetic relationship and demographic history of African sheep (Ovis aries) Breeds. Biology 2021, 10, 762. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Tang, Z.; Yu, H.; Han, L.; Ren, X.; Zhao, X.; Zhu, H.; Zhou, H. Early history of Chinese domestic sheep indicated by ancient DNA analysis of Bronze Age Individuals. J. Archaeol. Sci. 2011, 38, 896–902. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Geographical Region | n * | Group Composition (Breed) |
|---|---|---|
| Far Eastern (FAE) | 8 | Buubei (n = 5) |
| Baikal fine-fleeced (n = 3) | ||
| Volga (VOL) | 5 | Tsigai (n = 5) |
| North Caucasian (NCN) | 47 | Dagestan Mountain (n = 4) |
| Karachaev (n = 10) | ||
| Lezgin (n = 7) | ||
| Manych Merino (n = 5) | ||
| North Caucasian (n = 5) | ||
| Soviet Merino (n = 3) | ||
| Stavropol (n = 5) | ||
| Tushin (n = 8) | ||
| Siberian (SIB) | 17 | Altai Mountain (n = 5) |
| Buryat (n = 2) | ||
| Kulundin (n = 5) | ||
| Mongol (n = 1) | ||
| Tuva short-fat-tailed (n = 4) | ||
| Southern (SOU) | 38 | Volgograd (n = 8) |
| Groznenk (n = 7) | ||
| Kalmyk fat-rumped (n = 5) | ||
| Karakul (n = 6) | ||
| Salsk (n = 5) | ||
| Edilbai (n = 7) | ||
| Central (CEN) | 20 | Kuibyshev (n = 3) |
| Kuchugur (n = 3) | ||
| Romanov (n = 8) | ||
| Russian Longhaired (n = 6) |
| Primer | Nucleotide Sequences | Length of Amplification Product, b.p. |
|---|---|---|
| mtDNA_For1 | 5′-AGTACGGCGTAAAGCGTGTT-3′ | 2987 |
| mtDNA_Rev1 | 5′-AATGGTGCTCGGTTTGTTTC-3′ | |
| mtDNA_For2 | 5′-GAAAAGGCCCAAACGTTGTA-3′ | 3294 |
| mtDNA_Rev2 | 5′-GATATTATGGCTCATACTATTCCTATATA-3′ | |
| mtDNA_For3 | 5′-TCCTATATCAACACCTATTCTGATTCTT-3′ | 3525 |
| mtDNA_Rev3 | 5′-GGAAGTCAGAATGCGATGGT-3′ | |
| mtDNA_For4 | 5′-ACACCAAACCCACGCTTATC-3′ | 3183 |
| mtDNA_Rev4 | 5′-AAAATTGATTGCTGCGATGGGT-3′ | |
| mtDNA_For5 | 5′-TGAACGAGTTCACAGCCGAA-3′ | 2810 |
| mtDNA_Rev5 | 5′-ATTGTAAGTGGTGGGGTTGG-3′ | |
| mtDNA_For6 | 5′-AGCAATTCCCATAGCCTCCT-3′ | 3517 |
| mtDNA_Rev6 | 5′-GGCTGTTGCGGTAGTACTCT-3′ |
| Group 1 | n 2 | S 3 | K 4 | H 5 | Hd 6 (±SEM) 7 | π 8 (±SEM) | Tajima’s D 9 | Fu’s Fs 10 |
|---|---|---|---|---|---|---|---|---|
| FAE | 8 | 246 | 97,893 | 7 | 0.964 ±0.077 | 0.00589 ±0.00136 | 0.17425 ns13 | 3.582 ns13 |
| VOL | 5 | 49 | 20,400 | 5 | 1.000 ±0.126 | 0.00123 ±0.00017 | −1.00076 ns13 | 0.544 ns13 |
| NCN | 47 | 468 | 78,247 | 46 | 0.999 ±0.005 | 0.00471 ±0.00052 | −0.97712 ns13 | −8.987 *11 |
| SIB | 17 | 166 | 56,191 | 17 | 1.000 ±0.020 | 0.00339 ±0.00026 | 0.58964 ns13 | −2.097 **12 |
| SOU | 38 | 414 | 55,092 | 38 | 1.000 ±0.006 | 0.00332 ±0.00041 | −1.67111 ns13 | −10.572 *11 |
| CEN | 20 | 214 | 35,926 | 19 | 0.995 ±0.018 | 0.00216 ±0.00059 | −1.68045 ns13 | −2.62 **12 |
| Overall | 135 | 804 | 62,615 | 128 | 0.9992 ±0.010 | 0.00378 ±0.00027 | −1.91342 *11 | −31.631 *11 |
| Groups | FAE 1 | VOL 2 | SIB 3 | NCN 4 | SOU 5 | CEN 6 |
|---|---|---|---|---|---|---|
| FAE 1 | 0 | |||||
| VOL 2 | 0.39243 | 0 | ||||
| SIB 3 | 0.09707 | 0.29906 | 0 | |||
| NCN 4 | 0.16099 | 0.04873 | 0.05781 | 0 | ||
| SOU 5 | 0.29847 | 0.03248 | 0.10804 | 0.00937 | 0 | |
| CEN 6 | 0.43960 | 0.00818 | 0.28091 | 0.07288 | 0.04058 | 0 |
| Source of Variation | Freedom Degrees d.f. | Sum of Squares, SS | Variance Components, VC | Percentage of Variation V % |
|---|---|---|---|---|
| Among populations | 2 | 249.322 | 10.20234 | 22.13 |
| Within populations | 27 | 969.312 | 35.90044 | 77.87 |
| Total | 29 | 1218.633 | 46.10277 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koshkina, O.; Deniskova, T.; Dotsev, A.; Kunz, E.; Selionova, M.; Medugorac, I.; Zinovieva, N. Phylogenetic Analysis of Russian Native Sheep Breeds Based on mtDNA Sequences. Genes 2023, 14, 1701. https://doi.org/10.3390/genes14091701
Koshkina O, Deniskova T, Dotsev A, Kunz E, Selionova M, Medugorac I, Zinovieva N. Phylogenetic Analysis of Russian Native Sheep Breeds Based on mtDNA Sequences. Genes. 2023; 14(9):1701. https://doi.org/10.3390/genes14091701
Chicago/Turabian StyleKoshkina, Olga, Tatiana Deniskova, Arsen Dotsev, Elisabeth Kunz, Marina Selionova, Ivica Medugorac, and Natalia Zinovieva. 2023. "Phylogenetic Analysis of Russian Native Sheep Breeds Based on mtDNA Sequences" Genes 14, no. 9: 1701. https://doi.org/10.3390/genes14091701
APA StyleKoshkina, O., Deniskova, T., Dotsev, A., Kunz, E., Selionova, M., Medugorac, I., & Zinovieva, N. (2023). Phylogenetic Analysis of Russian Native Sheep Breeds Based on mtDNA Sequences. Genes, 14(9), 1701. https://doi.org/10.3390/genes14091701