
Characterization and Comparison of the Two Mitochondrial Genomes in the Genus Rana

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Assembly and Annotation
2.3. Bioinformatic Analysis
2.4. Phylogenetic Analyses
3. Results and Discussion
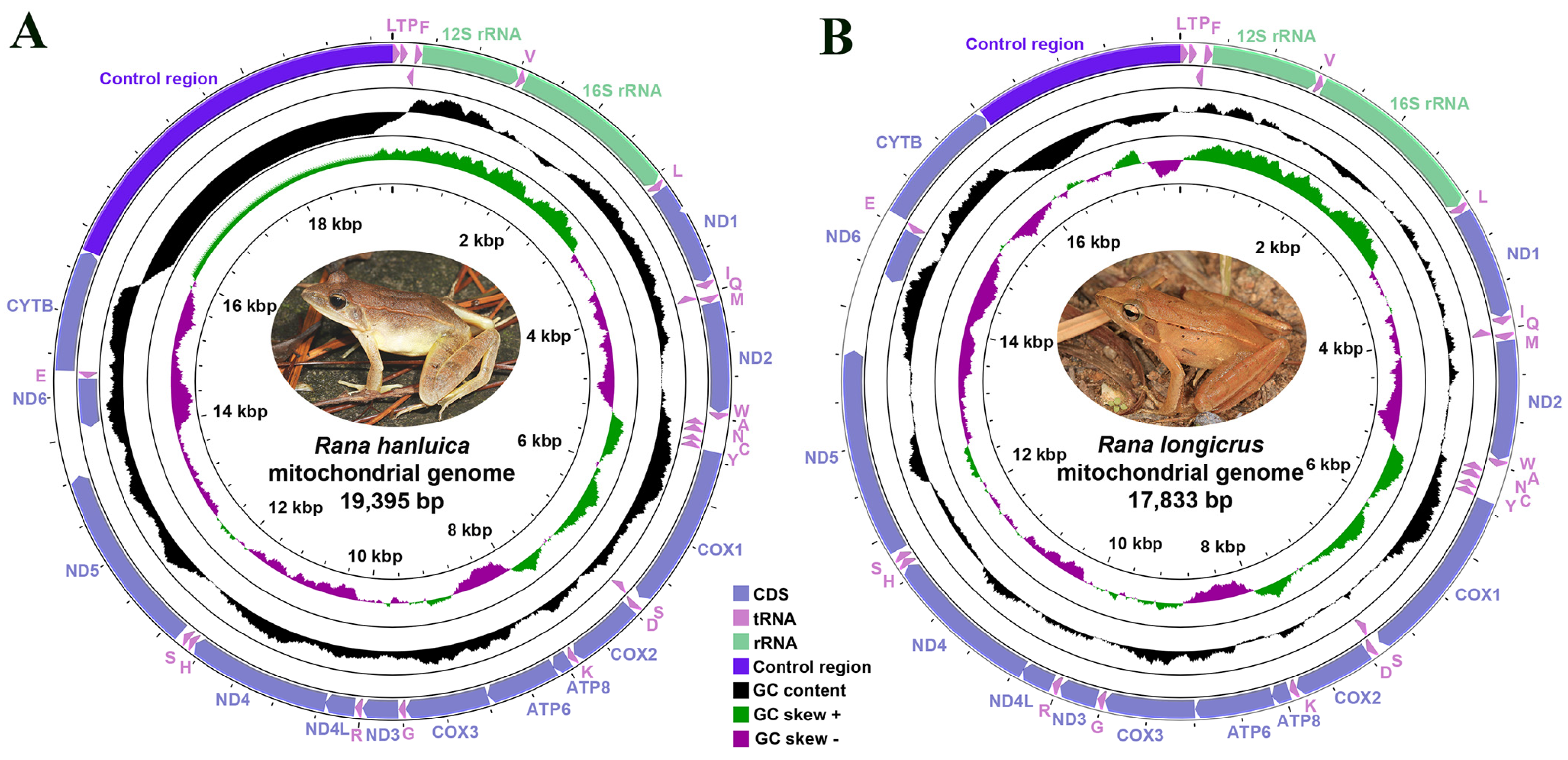
3.1. General Features of Mitogenomes of Two Rana Species
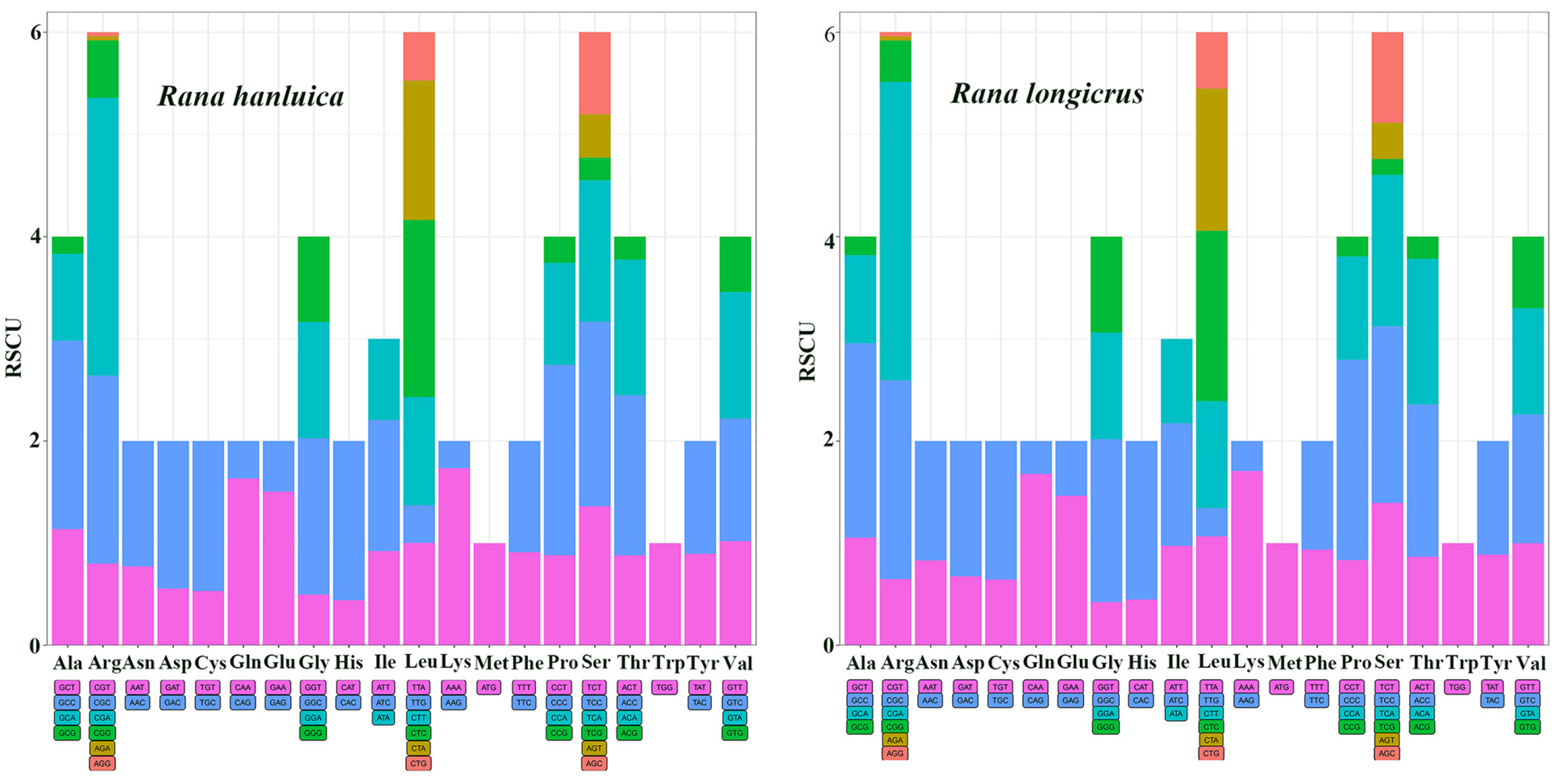
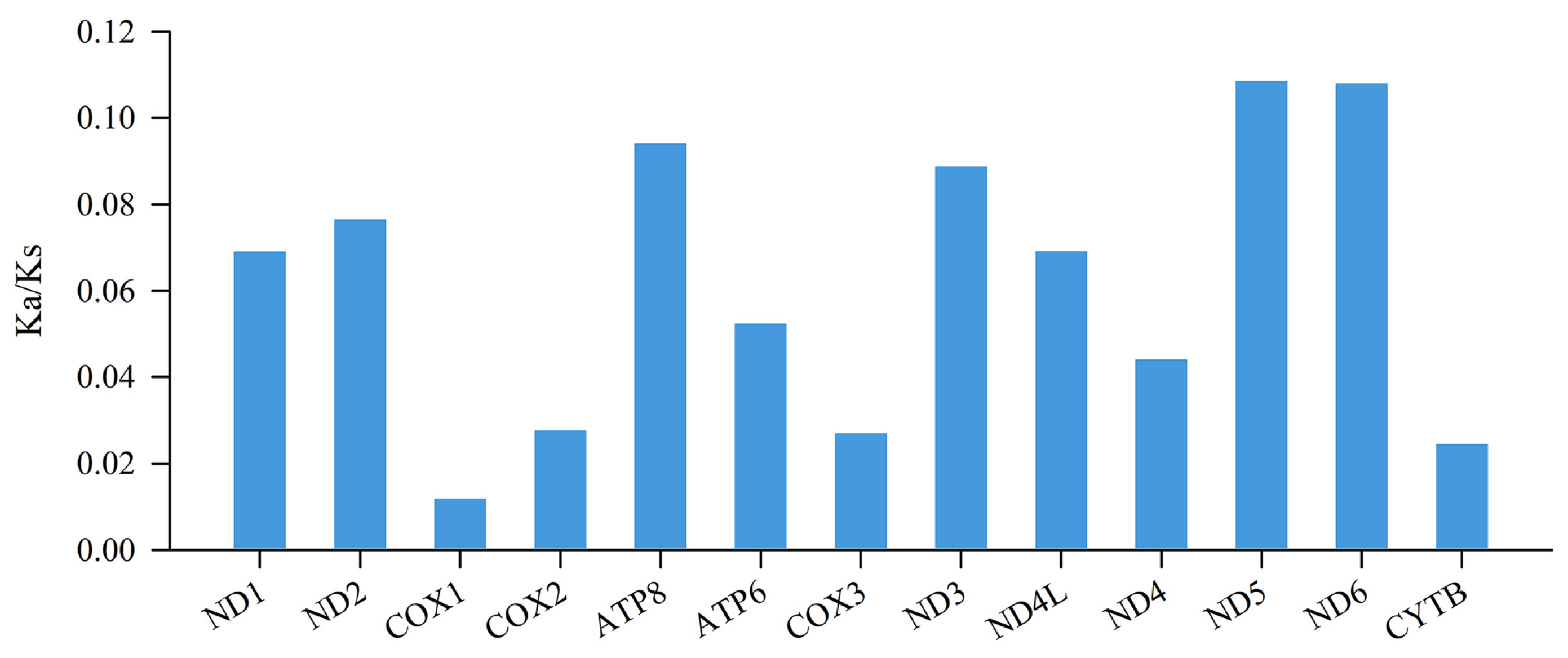
3.2. Protein-Coding Genes and Codon Usage
3.3. Transfer RNA and Ribosomal RNA Genes
3.4. Noncoding Regions
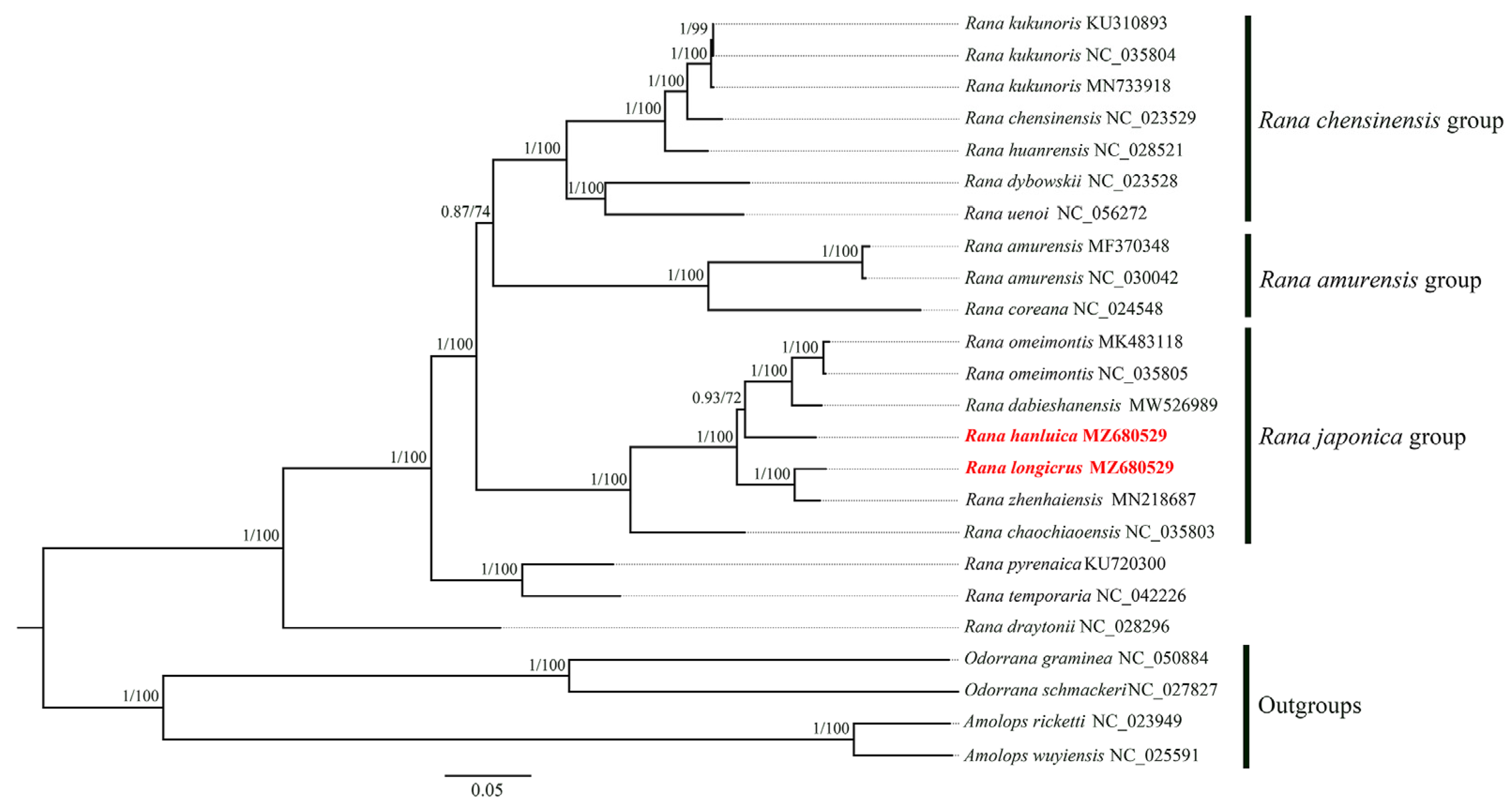
3.5. Phylogenetic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cai, Y.; Shen, S.; Lu, L.; Storey, K.B.; Yu, D.; Zhang, J. The complete mitochondrial genome of Pyxicephalus adspersus: High gene rearrangement and phylogenetics of one of the world’s largest frogs. PeerJ 2019, 7, e7532. [Google Scholar] [CrossRef]
- Li, F.; Lv, Y.; Wen, Z.; Bian, C.; Zhang, X.; Guo, S.; Shi, Q.; Li, D. The complete mitochondrial genome of the intertidal spider (Desis jiaxiangi) provides novel insights into the adaptive evolution of the mitogenome and the evolution of spiders. BMC Ecol. Evol. 2021, 21, 72. [Google Scholar] [CrossRef]
- Zhang, J.; Luu, B.E.; Yu, D.; Zhang, L.; Al-Attar, R.; Storey, K.B. The complete mitochondrial genome of Dryophytes versicolor: Phylogenetic relationship among Hylidae and mitochondrial protein-coding gene expression in response to freezing and anoxia. Int. J. Biol. Macromol. 2019, 132, 461–469. [Google Scholar] [CrossRef]
- Wang, Z.; Shi, X.; Tao, Y.; Wu, Q.; Bai, Y.; Guo, H.; Tang, D. The complete mitochondrial genome of Parasesarma pictum (Brachyura: Grapsoidea: Sesarmidae) and comparison with other Brachyuran crabs. Genomics 2019, 111, 799–807. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, Z.; Tang, D.; Xu, X.; Tao, Y.; Ji, C.; Wang, Z. Characterization and comparison of the mitochondrial genomes from two Alpheidae species and insights into the phylogeny of Caridea. Genomics 2020, 112, 65–70. [Google Scholar] [CrossRef]
- Chen, G.; Wang, B.; Liu, J.; Xie, F.; Jiang, J. Complete mitochondrial genome of Nanorana pleskei (Amphibia: Anura: Dicroglossidae) and evolutionary characteristics. Curr. Zool. 2011, 57, 785–805. [Google Scholar] [CrossRef]
- Xin, Z.; Liu, Y.; Zhu, X.; Wang, Y.; Zhang, H.; Zhang, D.; Zhou, C.; Tang, B.; Liu, Q. Mitochondrial genomes of two Bombycoidea insects and implications for their phylogeny. Sci. Rep. 2017, 7, 6544. [Google Scholar] [CrossRef] [PubMed]
- AmphibiaWeb. Available online: https://amphibiaweb.org (accessed on 20 July 2023).
- Yan, F.; Jiang, K.; Chen, H.; Fang, P.; Jin, J.; Li, Y.; Wang, S.; Murphy, R.W.; Che, J.; Zhang, Y. Matrilineal history of the Rana longicrus species group (Rana, Ranidae, Anura) and the description of a new species from Hunan, southern China. Asian Herpetol. Res. 2011, 2, 61–71. [Google Scholar]
- Boulenger, G.A. A monograph of the South Asian, Papuan, Melanesian, and Australian frogs of the genus Rana. Rec. Ind. Mus. 1920, 20, 1–126. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, S.; Zhu, H.; Li, P.; Yang, B.; Ma, J. Phylogeny and biogeography of South Chinese brown frogs (Ranidae, Anura). PLoS ONE 2017, 12, e175113. [Google Scholar] [CrossRef]
- Yuan, Z.; Zhou, W.; Chen, X.; Poyarkov, N.A.; Chen, H.; Jang-Liaw, N.; Chou, W.; Matzke, N.J.; Iizuka, K.; Min, M.; et al. Spatiotemporal diversification of the true frogs (genus Rana): A historical framework for a widely studied group of model organisms. Syst. Biol. 2016, 65, 824–842. [Google Scholar] [CrossRef] [PubMed]
- Fei, L.; Hu, S.Q.; Ye, C.Y.; Huang, Y.Z. Amphibia Vol. 3. Anura Ranidae. Fauna Sinica; Science Press: Beijing, China, 2009; pp. 1–22. [Google Scholar]
- Che, J.; Pang, J.; Zhao, E.; Matsui, M.; Zhang, Y. Phylogenetic relationships of the Chinese brown frogs (genus Rana) inferred from partial mitochondrial 12S and 16S rRNA gene sequences. Zool. Sci. 2007, 24, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Fei, L.; Ye, C.Y. On taxonomic status and relationships of Rana japonica group. China (Amphibia: Anura: Radidae). Cultum. Herpetol. Sin. 2000, 8, 74–80. [Google Scholar]
- Yang, B.; Zhou, Y.; Min, M.; Matsui, M.; Dong, B.; Li, P.; Fong, J.J. Diversity and phylogeography of Northeast Asian brown frogs allied to Rana dybowskii (Anura, Ranidae). Mol. Phylogenet. Evol. 2017, 112, 148–157. [Google Scholar] [CrossRef]
- Wan, H.; Lyu, Z.; Qi, S.; LIU, L.; Wang, Y. A new species of the Rana japonica group (Anura, Ranidae, Rana) from China, with a taxonomic proposal for the R. johnsi group. Zookeys 2020, 942, 141–158. [Google Scholar] [CrossRef]
- Zhu, S.; Zhang, X.; Zhang, S.; Zhang, R.; Liu, S.; Xiong, R. Molecular identification of a Rana hanluica population distributed in Youyang County, Chongqing City in China. Open J. Nat. Sci. 2018, 6, 1–8. [Google Scholar] [CrossRef]
- Pope, C.H.; Boring, A.M. A survey of Chinese Amphibia. Peking Nat. Hist. Bull. 1940, 15, 13–86. [Google Scholar]
- Wu, Y.; Yan, F.; Stuart, B.L.; Prendini, E.; Suwannapoom, C.; Dahn, H.A.; Zhang, B.; Cai, H.; Xu, Y.; Jiang, K.; et al. A combined approach of mitochondrial DNA and anchored nuclear phylogenomics sheds light on unrecognized diversity, phylogeny, and historical biogeography of the torrent frogs, genus Amolops (Anura: Ranidae). Mol. Phylogenet. Evol. 2020, 148, 106789. [Google Scholar] [CrossRef]
- Jin, J.; Yu, W.; Yang, J.; Song, Y.; DePamphilis, C.W.; Yi, T.; Li, D. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Meng, G.; Li, Y.; Yang, C.; Liu, S. MitoZ: A toolkit for animal mitochondrial genome assembly, annotation and visualization. Nucleic Acids Res. 2019, 47, e63. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Yang, Z. Computational Molecular Evolution; Oxford University Press: Oxford, UK; New York, NY, USA, 2006. [Google Scholar]
- Dong, B.; Zhou, Y.; Yang, B. The complete mitochondrial genome of the Rana huanrensis (Anura: Ranidae). Mitochondrial DNA A 2016, 27, 4551–4552. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yin, W.; Xia, R.; Lei, G.; Fu, C. Complete mitochondrial genome of a brown frog, Rana kunyuensis (Anura: Ranidae). Mitochondrial DNA A 2016, 27, 34–35. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lei, G.; Fu, C. Complete mitochondrial genomes of two brown frogs, Rana dybowskii and Rana cf. chensinensis (Anura: Ranidae). Mitochondrial DNA A 2016, 27, 155–156. [Google Scholar] [CrossRef] [PubMed]
- Peso-Fernández, M.; Ponti De La Iglesia, R.; Ponz Segrelles, G.; Gonzalez Martinez, R.; Arcones Segovia, A.; Vieites, D.R. The complete mitochondrial genome of the Endangered European brown frog Rana pyrenaica through RNAseq. Mitochondrial DNA B 2016, 1, 394–396. [Google Scholar] [CrossRef]
- Liu, P.; Wang, H.; Zhao, W. Sequencing and analysis of the complete mitochondrial genome of Rana amurensis (Anura: Ranidae). Mitochondrial DNA B 2017, 2, 424–425. [Google Scholar] [CrossRef]
- Chen, J. The complete mitochondrial genome of common terrestrial frog (Rana temporaria). Mitochondrial DNA B 2018, 3, 978–979. [Google Scholar] [CrossRef]
- Jiang, L.; Zhang, M.; Deng, L.; Xu, Z.; Shi, H.; Jia, X.; Lai, Z.; Ruan, Q.; Chen, W. Characteristics of the mitochondrial genome of Rana omeimontis and related species in Ranidae: Gene rearrangements and phylogenetic relationships. Ecol. Evol. 2020, 10, 12817–12837. [Google Scholar] [CrossRef]
- Wang, J.; Li, Z.; Gao, H.; Liu, Z.; Teng, L. The complete mitochondrial genome of the Rana kukunoris (Anura: Ranidae) from Inner Mongolia, China. Mitochondrial DNA B 2020, 5, 586–587. [Google Scholar] [CrossRef] [PubMed]
- Suk, H.Y.; Jeon, J.Y.; Kim, D.; Cha, S.; Min, M. The complete mitochondrial genome information of Rana uenoi (Amphibia, Anura, Ranidae) and the phylogenetic implication. Mitochondrial DNA B 2021, 6, 689–690. [Google Scholar] [CrossRef]
- Huang, M.; Zhao, Q.; Wu, Y. Characterization of the complete mitochondrial genome of Zhenhai brown frog Rana zhenhaiensis (Anura: Ranidae). Mitochondrial DNA B 2019, 4, 3204–3205. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Duan, R.; Kong, X.; Wang, H.; Zhu, H. The complete mitochondrial genome of Amolops wuyiensis (Anura: Ranidae). Mitochondrial DNA A 2016, 27, 1705–1706. [Google Scholar]
- Li, Y.; Wu, X.; Zhang, H.; Yan, P.; Xue, H.; Wu, X. The complete mitochondrial genome of Amolops ricketti (Amphidia, Anura, Ranidae). Mitochondrial DNA A 2016, 27, 242–243. [Google Scholar] [CrossRef]
- Jin, X.; Li, W.; Hu, S.; Li, W.; Yang, J. The complete mitochondrial genome of large odorous frog, Odorrana graminea (Amphibia: Ranidae) and phylogenetic analysis. Mitochondrial DNA B 2020, 5, 3139–3140. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Pereira, S.L. Mitochondrial genome organization and vertebrate phylogenetics. Genet. Mol. Biol. 2000, 23, 745–752. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Wu, X.; Xue, H.; Yan, P.; Wu, X. A novel mitogenomic rearrangement for Odorrana schmackeri (Anura: Ranidae) and phylogeny of Ranidae inferred from thirteen mitochondrial protein-coding genes. Amphib.-Reptil. 2014, 35, 331–343. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, J.; Zhai, X.; Zhu, Y.; Chen, X. Complete mitochondrial genome of the green odorous frog Odorrana margaretae (Anura: Ranidae). Mitochondrial DNA 2015, 26, 487–488. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yu, J.; Liu, J.; Zhou, M.; Li, B.; Ouyang, B. Three new Ranidae mitogenomes and the evolution of mitochondrial gene rearrangements among Ranidae species. Asian Herpetol. Res. 2018, 9, 85–98. [Google Scholar]
- Jeffrey, L.B. Survey and summary animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef]
- Jiang, L.; Zhao, L.; Cheng, D.; Zhu, L.; Zhang, M.; Ruan, Q.; Chen, W. The complete mitochondrial genome sequence of the Sichuan Digging Frog, Kaloula rugifera (Anura: Microhylidae) and its phylogenetic implications. Gene 2017, 626, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Kurisaki, M.; Hashiguchi, Y.; Kumazawa, Y. Variation and evolution of polyadenylation profiles in sauropsid mitochondrial mRNAs as deduced from the high-throughput RNA sequencing. BMC Genom. 2017, 18, 665. [Google Scholar] [CrossRef]
- Mayfield, J.E.; McKenna, J.F. AT rich sequences in vertebrate DNA. Chromosoma 1978, 67, 157–163. [Google Scholar] [CrossRef]
- Cai, Y.; Li, Q.; Zhang, J.; Storey, K.B.; Yu, D. Characterization of the mitochondrial genomes of two toads, Anaxyrus americanus (Anura: Bufonidae) and Bufotes pewzowi (Anura: Bufonidae), with phylogenetic and selection pressure analyses. PeerJ 2020, 8, e8901. [Google Scholar] [CrossRef]
- Shahzadi, I.; Mehmood, F.; Ali, Z.; Malik, M.S.; Waseem, S.; Mirza, B.; Ahmed, I.; Waheed, M.T. Comparative analyses of chloroplast genomes among three Firmiana species: Identification of mutational hotspots and phylogenetic relationship with other species of Malvaceae. Plant Gene 2019, 19, 100199. [Google Scholar]
- Zhang, X.; Liu, P.; Xu, S.; Rizo, E.Z.; Zhang, Q.; Dumont, H.J.; Han, B. Geographic Variation of Phyllodiaptomus tunguidus Mitogenomes: Genetic Differentiation and Phylogeny. Front. Genet. 2021, 12, 711992. [Google Scholar] [CrossRef]
- Chen, L.; Lin, Y.; Xiao, Q.; Lin, Y.; Du, Y.; Lin, C.; Ward-Fear, E.; Hu, C.; Qu, Y.; Li, H. Characterization of the complete mitochondrial genome of the many-lined sun skink (Eutropis multifasciata) and comparison with other Scincomorpha species. Genomics 2020, 113, 2526–2536. [Google Scholar] [CrossRef] [PubMed]
- Kurabayashi, A.; Ueshima, R. Complete sequence of the mitochondrial DNA of the primitive opisthobranch gastropod Pupa strigosa: Systematic implication of the genome organization. Mol. Biol. Evol. 2000, 17, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Wolstenholme, D.R. Animal mitochondrial DNA: Structure and evolution. Int. Rev. Cyto. 1992, 141, 173–216. [Google Scholar]
- Wang, C.; Qian, L.; Zhang, C.; Guo, W.; Pan, T.; Wu, J.; Wang, H.; Zhang, B. A new species of Rana from the Dabie Mountains in eastern China (Anura, Ranidae). Zookeys 2017, 724, 135. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Size (bp) | A + T Content (%) | G + C Content (%) | AT−Skew | GC−Skew | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| RH | RL | RH | RL | RH | RL | RH | RL | RH | RL | |
| Whole genome | 19,395 | 17,833 | 59.1 | 55.6 | 40.9 | 44.4 | −0.045 | −0.007 | −0.314 | −0.328 |
| PCGs | 11,279 | 11,288 | 53.9 | 54 | 46.1 | 46 | −0.088 | −0.082 | −0.322 | −0.321 |
| rRNA genes | 2509 | 2509 | 55.4 | 55.9 | 44.6 | 44.1 | 0.157 | 0.162 | −0.15 | −0.148 |
| tRNA genes | 1530 | 1530 | 57.6 | 56.9 | 42.4 | 43.1 | 0.015 | 0.024 | 0.045 | 0.032 |
| CR | 3602 | 1930 | 78.8 | 64.2 | 21.2 | 35.8 | −0.134 | −0.031 | −0.251 | −0.294 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-M.; Zhang, C.-Y.; Luo, S.-T.; Ding, G.-H.; Qiao, F. Characterization and Comparison of the Two Mitochondrial Genomes in the Genus Rana. Genes 2023, 14, 1786. https://doi.org/10.3390/genes14091786
Wang Y-M, Zhang C-Y, Luo S-T, Ding G-H, Qiao F. Characterization and Comparison of the Two Mitochondrial Genomes in the Genus Rana. Genes. 2023; 14(9):1786. https://doi.org/10.3390/genes14091786
Chicago/Turabian StyleWang, Yan-Mei, Chi-Ying Zhang, Si-Te Luo, Guo-Hua Ding, and Fen Qiao. 2023. "Characterization and Comparison of the Two Mitochondrial Genomes in the Genus Rana" Genes 14, no. 9: 1786. https://doi.org/10.3390/genes14091786
APA StyleWang, Y. -M., Zhang, C. -Y., Luo, S. -T., Ding, G. -H., & Qiao, F. (2023). Characterization and Comparison of the Two Mitochondrial Genomes in the Genus Rana. Genes, 14(9), 1786. https://doi.org/10.3390/genes14091786