

Identification and Validation of Reference Genes for RT-qPCR Analysis in Reed Canary Grass during Abiotic Stress
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Abiotic Stress Treatments
2.2. RNA Extraction and Reverse Transcription
2.3. Primer Design and Validation
2.4. Quantitative RT-qPCR
2.5. Data Analysis
2.6. Validation of Reference Genes
3. Results
3.1. The Effectiveness of the Primers of 13 Specific Reference Genes
3.2. Expression of Reference Gene
3.3. Candidate Reference Gene Stability Analysis
3.3.1. GeNorm Analysis
3.3.2. BestKeeper Analysis
3.3.3. NormFinder Analysis
3.3.4. RefFinder Analysis
3.4. Validation the Stability of Reference Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Oram, R.N.; Ferreira, V.; Culvenor, R.A.; Hopkins, A.A.; Stewart., A. The first century of Phalaris aquatica L. cultivation and genetic improvement: A review. Crop Pasture Sci. 2009, 60, 1–15. [Google Scholar] [CrossRef]
- Rebecca, B.; Michelle, D.; Luke, P.; Sukhjiwan, K.; Richard, C.; Kevin, S.; German, S.; John, F.; Noel, C. Generation and Characterisation of a Reference Transcriptome for Phalaris (Phalaris aquatica L.). Agronomy 2017, 7, 14. [Google Scholar]
- Li, P.; Yan, J.J.; Bai, S.Q.; Chen, Z.H.; You, M.H.; Su, G.P.; Dao, Z.X. A preliminary study on the silage effect of Elymus sibiricus L. and Phalaris aquatica L. in alpine pastures of northwestern Sichuan. Acta Agrestia Sin. 2012, 20, 368–372. [Google Scholar]
- Polechońska, L.; Klink, A. Trace metal bioindication and phytoremediation potentialities of Phalaris arundinacea L. (reed canary grass). J. Geochem. Explor. 2014, 146, 27–33. [Google Scholar] [CrossRef]
- Antonkiewicz, J.; Kołodziej, B.; Bielińska, E.J. The use of reed canary grass and giant miscanthus in the phytoremediation of municipal sewage sludge. Environ. Sci. Pollut. Res. 2016, 23, 9505–9517. [Google Scholar] [CrossRef] [PubMed]
- Melts, I.; Ivask, M.; Geetha, M.; Takeuchi, K.; Heinsoo, K. Combining bioenergy and nature conservation: An example in wetlands. Renew. Sustain. Energy Rev. 2019, 111, 293–302. [Google Scholar] [CrossRef]
- Song, J.Z.; Ling, P.P.; Fu, W.G. Effect of water stress and rewatering on the physiological and biochemical characteristics of Phalaris arundinacea. Acta Prataculturae Sin. 2012, 21, 62–69. [Google Scholar]
- Zhang, Y.L.; Nie, W.W.; Ren, X.Z.; Zhu, T.X. Effects of Mixed Saline Stress on the Absorption and Transportation of K+ and Na+ of Two Reed Canary grass. Chin. J. Grassl. 2010, 32, 28–33. [Google Scholar]
- Manfred, K.; Niina, H.; Jim, G.; Paul, C.; John, F.; Krishna, A.S.; Filippo, U.; Tia, V.; Laxmi, P.; Susanne, B. Transcriptome characterization and differentially expressed genes under waterlogging and drought stress in the biomass grasses Phalaris arundinacea and Dactylis glomerata. Ann. Bot.-London 2019, 4, 14. [Google Scholar]
- Kulcheski, F.R.; Marcelino-Guimaraes, F.C.; Nepomuceno, A.L.; Abdelnoor, R.V.; Margis, R. The use of microRNAs as reference genes for quantitative polymerase chain reaction in soybean. Anal. Biochem. 2010, 406, 185–192. [Google Scholar] [CrossRef]
- Wen, L.; Tan, B.; Guo, W.W. Estimating transgene copy number in precocious trifoliate orange by TaqMan real-time PCR. Plant Cell Tissue Organ Cult. 2012, 109, 363–371. [Google Scholar] [CrossRef]
- Bustin, S.A.; Vladimir, B.; Garson, J.A.; Jan, H.; Jim, H.; Mikael, K.; Reinhold, M.; Tania, N.; Pfaffl, M.W.; Shipley, G.L. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Qu, R.; Miao, Y.; Cui, Y.; Cao, Y.; Zhou, Y.; Tang, X.; Yang, J.; Wang, F. Selection of reference genes for the quantitative real-time PCR normalization of gene expression in Isatis indigotica fortune. BMC Mol. Biol. 2019, 20, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Sun, H.; Zhang, R.; Yang, Q.; Zhang, X. Selection of reference gene from Gracilaria lemaneiformis under temperature stress. J. Appl. Phycol. 2014, 27, 1365–1372. [Google Scholar] [CrossRef]
- Yang, Q.; Yin, J.; Li, G.; Qi, L.; Yang, F.; Wang, R.; Li, G. Reference gene selection for qRT-PCR in Caragana korshinskii Kom. under different stress conditions. Mol. Biol. Rep. 2014, 41, 2325–2334. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ishikawa, T.; Michiue, T.; Zhu, B.L.; Guan, D.W.; Maeda, H. Stability of endogenous reference genes in postmortem human brains for normalization of quantitative real-time PCR data: Comprehensive evaluation using geNorm, NormFinder, and BestKeeper. Int. J. Legal Med. 2012, 126, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zhang, C.; Lan, H.; Gao, S.; Liu, H.; Jian, L.; Cao, M.; Pan, G.; Rong, T.; Zhang, S. Validation of Potential Reference Genes for qPCR in Maize across Abiotic Stresses, Hormone Treatments, and Tissue Types. PLoS ONE 2014, 9, e95445. [Google Scholar] [CrossRef]
- Wu, T.; Li, W.F.; Zhang, J.H.; Han, S.Y.; Qi, L.W. Stability analysis of reference genes for larch gene expression studies by quantitative real-time PCR. For. Res. 2013, 26, 1–8. [Google Scholar]
- Schmer, M.R.; Vogel, E.K.; Mitchell, R.; Perrin, M.R. Net energy of cellulosic ethanol from switchgrass. Proc. Natl. Acad. Sci. USA 2008, 105, 464–469. [Google Scholar] [CrossRef]
- Jia, J.X.; Xiong, Y.; Xiong, Y.L.; Ji, X.F.; Li, D.X.; Bai, S.Q.; Yan, L.J.; You, M.H.; Ma, X.; Zhang, J.B. Transcriptomic sequencing analysis, development, and validation of EST-SSR markers in reed canary grass. Grass Res. 2023, 3, 17. [Google Scholar] [CrossRef]
- Vandesompele, J.; Preter, K.D.; Pattyn, F.; Poppe, B.; Roy, N.V.; Paepe, A.D.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 341–3411. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-Based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res. 2018, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper-Excel-Based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Clark, O.; Thomsen, M. Restoring wetlands dominated by Phalaris arundinacea (Reed Canary Grass) with multiple treatments: Haying, spraying, and establishing aggressive competitors. Ecol. Restor. 2020, 38, 6–9. [Google Scholar] [CrossRef]
- Maeda, Y.; Hirano, S.; Yoshiba, M.; Tadano, T. Variations in salt tolerance of reed canarygrass (Phalaris arundinacea L.) plants grown at sites with different degrees of cattle urine contamination (Plant Nutrition). Soil Sci. Plant Nutr. 2006, 52, 13–20. [Google Scholar] [CrossRef]
- Jones, T.A.; Carlson, I.T. Inheritance of the Virescent Trait in Reed Canarygrass. J. Hered. 1988, 79, 218–221. [Google Scholar] [CrossRef]
- Wang, X.; He, Y.; Zhang, C.; Tian, Y.A.; Lin, H. Physiological and transcriptional responses of Phalaris arundinacea under waterlogging conditions. J. Plant Physiol. 2021, 261, 153428. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Q.; Jiang, Y.; Li, Y.; Zhang, H.; Li, R. Reference genes identification for normalization of qPCR under multiple stresses in Hordeum brevisubulatum. Plant Methods 2018, 14, 110. [Google Scholar] [CrossRef]
- Boava, L.P.; Laia, M.L.; Jacob, T.R.; Dabbas, K.M.; Alves, J.F.G.; Ferro, J.A.; Ferro, M.I.; Furtado, E.L. Selection of endogenous genes for gene expression studies in Eucalyptus under biotic (Puccinia psidii) and abiotic (acibenzolar-S-methyl) stresses using RT-qPCR. BMC Res. Notes 2010, 3, 43–52. [Google Scholar] [CrossRef]
- Andrade, L.M.D.; Brito, M.D.S. Reference genes for normalization of qPCR assays in sugarcane plants under water deficit. Plant Methods 2017, 13, 28. [Google Scholar] [CrossRef]
- Udvardi, M.K.; Scheible, C. Eleven golden rules of quantitative RT-PCR. Plant Cell Online 2008, 20, 1736–1737. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, L.; Mauriat, M.; Guénin, S.; Pelloux, J.; Lefebvre, J.F.; Louvet, R.; Rusterucci, C.; Moritz, T.; Guerineau, F.; Bellini, C.; et al. The lack of a systematic validation of reference genes: A serious pitfall undervalued in reverse transcription-polymerase chain reaction (RT-PCR) analysis in plants. Plant Biotechnol. J. 2008, 6, 609–618. [Google Scholar] [CrossRef]
- Kim, H.T.; Kim, K.P.; Lledias, F.; Kisselev, A.F.; Scaglione, K.M.; Skowyra, D.; Gygi, S.P.; Goldberg, A.L. Certain Pairs of Ubiquitin-conjugating Enzymes (E2s) and Ubiquitin-Protein Ligases (E3s) Synthesize Nondegradable Forked Ubiquitin Chains Containing All Possible Isopeptide Linkages. J. Biol. Chem. 2007, 282, 17375. [Google Scholar] [CrossRef] [PubMed]
- Bahmani, R.; Kim, D.G.; Lee, B.D.; Hwang, S. Over-expression of tobacco UBC1 encoding a ubiquitin-conjugating enzyme increases cadmium tolerance by activating the 20S/26S proteasome and by decreasing Cd accumulation and oxidative stress in tobacco (Nicotiana tabacum). Plant Mol. Biol. 2017, 94, 433–451. [Google Scholar] [CrossRef]
- Chung, E.; Cho, C.W.; So, H.A.; Kang, J.S.; Chung, Y.S.; Lee, J.H. Overexpression of vrubc1, a mung bean e2 ubiquitin-conjugating enzyme, enhances osmotic stress tolerance in Arabidopsis. PLoS ONE 2013, 8, e66056. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.R.; Feng, Y.Y.; Yang, M.J.; Xiao, Y.; Liu, Y.S.; Yuan, Y.; Li, Z.; Zhang, Y.; Zhuo, M.; Zhang, J. Systematic screening, and validation of reliable reference genes for qRT-PCR analysis in Okra (Abelmoschus esculentus). Sci. Rep. 2022, 12, 12913. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lu, H.; He, Z.; Wang, C.; Wang, Y.; Ji, X. Selection of appropriate reference genes for quantitative real-time reverse transcription PCR in Betula platyphylla under salt and osmotic stress conditions. PLoS ONE 2019, 14, e225926. [Google Scholar] [CrossRef]
- Wang, X.; Kong, X.; Liu, S.; Huang, H.; Xu, Y. Selection of Reference Genes for Quantitative Real-Time PCR in Chrysoperla nipponensis (Neuroptera: Chrysopidae) Under Tissues in Reproduction and Diapause. J. Insect Sci. 2020, 20, 20. [Google Scholar] [CrossRef]
- Tang, X.; Zhang, N.; Si, H.; Calderón-Urrea, A. Selection and validation of reference genes for RT-qPCR analysis in potato under abiotic stress. Plant Methods 2017, 13, 85. [Google Scholar] [CrossRef]
- Li, Y.; Deng, Y.; Li, Z.; Liu, Z.; Cui, X. Composition, physicochemical properties, and anti-fatigue activity of water-soluble okra (Abelmoschus esculentus) stem pectins. Int. J. Biol. Macromol. 2020, 165, 2630–2639. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Du, H.; Jin, Q.; Lin, Y.; Zhong, Y.; Yao, C.; Xu, X.; Wu, T. Stability determination of candidate reference genes in cucumber plants subjected to stresses from Phytophthora melonis. Height. Sci. Publ. Corp. 2019, 3, 28–35. [Google Scholar]
- Jiang, C.X.H.L. Genome-wide identification of plasma membrane aquaporin gene family in Populus and functional identification of PIP-1;1 involved in osmotic stress. Environ. Exp. Bot. 2020, 179, 10. [Google Scholar] [CrossRef]
- Vega, J.; Teshome, A.; Klaas, M.; Grant, J.; Barth, S. Physiological and transcriptional response to drought stress among bioenergy grass Miscanthus species. Biotechnol. Biofuels 2021, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, Z.; Fang, S.; Liu, Y.; Shang, X. Integrative analysis of metabolome and transcriptome reveals molecular regulatory mechanism of flavonoid biosynthesis in Cyclocarya paliurus under salt stress. Ind. Crop. Prod. 2021, 170, 113823. [Google Scholar] [CrossRef]
- Yu, R.; Tang, Y.; Liu, C.; Du, X.; Miao, C.; Shi, G. Comparative transcriptomic analysis reveals the roles of ROS scavenging genes in response to cadmium in two pak choi cultivars. Sci. Rep. 2017, 7, 9217. [Google Scholar] [CrossRef]






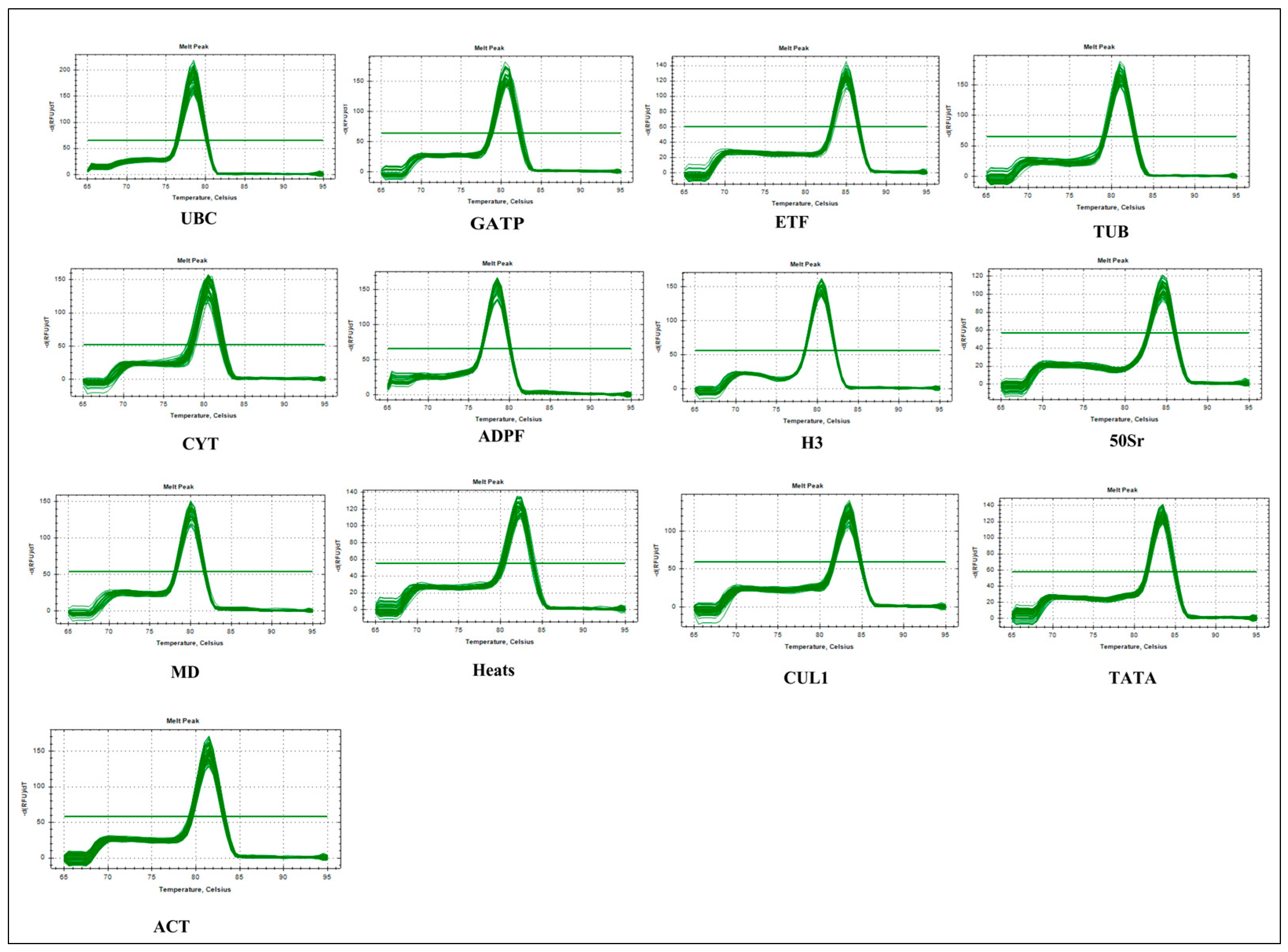{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank | Drought | Salt | Cd | Waterlogging |
|---|---|---|---|---|
| 1 | H3 (0.87 ± 0.26) | CYT (1.4 ± 0.42) | H3 (1 ± 0.29) | H3 (1.26 ± 0.35) |
| 2 | UBC (0.94 ± 0.29) | ETF (1.42 ± 0.4) | UBC (1.22 ± 0.37) | ETF (1.27 ± 0.34) |
| 3 | MD (1.17 ± 0.36) | UBC (1.54 ± 0.46) | CYT (1.67 ± 0.51) | TUB (1.49 ± 0.4) |
| 4 | ACT (1.65 ± 0.49) | ACT (1.59 ± 0.44) | CUL1 (2.12 ± 0.52) | CUL1 (1.78 ± 0.44) |
| 5 | ETF (1.97 ± 0.55) | MD (1.73 ± 0.53) | TATA (2.2 ± 0.64) | UBC (2.13 ± 0.63) |
| 6 | CUL1 (2.05 ± 0.51) | TATA (1.83 ± 0.53) | GATP (2.39 ± 0.69) | 50Sr (2.33 ± 0.57) |
| 7 | 50Sr (2.65 ± 0.72) | Heats (1.85 ± 0.53) | Heats (2.48 ± 0.71) | ADPF (2.34 ± 0.69) |
| 8 | ADPF (2.66 ± 0.82) | TUB (2.02 ± 0.53) | MD (2.49 ± 0.76) | Heats (2.54 ± 0.74) |
| 9 | Heats (2.8 ± 0.85) | 50Sr (2.06 ± 0.47) | 50Sr (2.57 ± 0.64) | MD (2.73 ± 0.86) |
| 10 | CYT (2.95 ± 0.9) | ADPF (2.13 ± 0.63) | ADPF (2.89 ± 0.82) | GATP (3.14 ± 0.92) |
| 11 | TATA (3.38 ± 1) | H3 (2.66 ± 0.78) | ETF (3.01 ± 0.8) | CYT (4.35 ± 1.28) |
| 12 | TUB (3.59 ± 0.96) | CUL1 (2.69 ± 0.66) | TUB (3.01 ± 0.82) | TATA (4.4 ± 1.29) |
| 13 | GATP (4.1 ± 1.21) | GATP (3.3 ± 0.94) | ACT (4.26 ± 1.24) | ACT (4.48 ± 1.26) |
| Rank | Drought | Salt | Cd | Waterlogging |
|---|---|---|---|---|
| 1 | ACT (0.353) | ETF (0.362) | H3 (0.436) | H3 (2.13 ± 0.63) |
| 2 | UBC (0.372) | CYT (0.382) | UBC (0.487) | ETF (3.14 ± 0.92) |
| 3 | H3 (0.421) | ETF (0.362) | 50Sr (0.501) | TUB (1.27 ± 0.34) |
| 4 | MD (0.43) | CYT (0.382) | CYT (0.541) | UBC (1.49 ± 0.4) |
| 5 | CUL1 (0.614) | TUB (0.436) | TATA (0.566) | ADPF (4.35 ± 1.28) |
| 6 | ETF (0.749) | ACT (0.523) | CUL1 (0.666) | GATP (2.34 ± 0.69) |
| 7 | CYT (0.918) | UBC (0.584) | TUB (0.732) | Heats (1.26 ± 0.35) |
| 8 | 50Sr (0.944) | MD (0.601) | Heats (0.768) | MD (2.33 ± 0.57) |
| 9 | Heats (0.958) | TATA (0.675) | GATP (0.794) | CUL1 (2.73 ± 0.86) |
| 10 | TATA (1.022) | ADPF (0.676) | ETF (0.833) | TATA (2.54 ± 0.74) |
| 11 | TUB (1.114) | 50Sr (0.684) | MD (0.977) | CYT (1.78 ± 0.44) |
| 12 | ADPF (1.149) | CUL1 (0.763) | ADPF (1.098) | 50Sr (4.4 ± 1.29) |
| 13 | GATP (1.288) | Heats (0.822) | ACT (1.141) | ACT (4.48 ± 1.26) |
| Method | Stability (High-Low) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | |
| Drought | |||||||||||||
| BestKeeper | H3 | UBC | MD | ACT | CUL1 | ETF | 50Sr | ADPF | Heats | CYT | TUB | TATA | GATP |
| Normfinder | ACT | UBC | H3 | MD | CUL1 | ETF | CYT | 50Sr | Heats | TATA | TUB | ADPF | GATP |
| Genorm | UBC | H3 | MD | CUL1 | ACT | ETF | 50Sr | CYT | Heats | TATA | TUB | ADPF | GATP |
| RefFinder | UBC | H3 | ACT | MD | CUL1 | ETF | 50Sr | CYT | Heats | TATA | ADPF | TUB | GATP |
| Salt | |||||||||||||
| BestKeeper | ETF | CYT | ACT | UBC | 50Sr | TUB | TATA | MD | Heats | ADPF | CUL1 | H3 | GATP |
| Normfinder | ETF | CYT | TUB | ACT | UBC | MD | TATA | ADPF | 50Sr | CUL1 | Heats | H3 | GATP |
| Genorm | ETF | TUB | CYT | ACT | UBC | MD | TATA | 50Sr | ADPF | CUL1 | Heats | H3 | GATP |
| RefFinder | ETF | CYT | TUB | ACT | UBC | MD | TATA | 50Sr | ADPF | CUL1 | Heats | H3 | GATP |
| Cd | |||||||||||||
| BestKeeper | H3 | UBC | CYT | CUL1 | 50Sr | TATA | GATP | Heats | MD | TUB | ADPF | ETF | ACT |
| Normfinder | H3 | UBC | 50Sr | CYT | TATA | CUL1 | TUB | Heats | GATP | ETF | MD | ADPF | ACT |
| Genorm | UBC | H3 | CYT | 50Sr | TATA | CUL1 | Heats | TUB | GATP | ETF | MD | ADPF | ACT |
| RefFinder | H3 | UBC | CYT | 50Sr | TATA | CUL1 | Heats | TUB | GATP | MD | ETF | ADPF | ACT |
| Waterlogging | |||||||||||||
| BestKeeper | ETF | H3 | TUB | CUL1 | 50Sr | UBC | ADPF | Heats | MD | GATP | ACT | CYT | TATA |
| Normfinder | H3 | ETF | TUB | UBC | ADPF | GATP | Heats | MD | CUL1 | TATA | CYT | 50Sr | ACT |
| Genorm | ETF | TUB | UBC | ADPF | H3 | CUL1 | GATP | Heats | MD | 50Sr | CYT | TATA | ACT |
| RefFinder | ETF | TUB | H3 | UBC | ADPF | CUL1 | GATP | Heats | MD | 50Sr | CYT | TATA | ACT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, X.; Xiong, Y.; Xiong, Y.; Li, D.; Yu, Q.; Lei, X.; You, M.; Bai, S.; Zhang, J.; Ma, X. Identification and Validation of Reference Genes for RT-qPCR Analysis in Reed Canary Grass during Abiotic Stress. Genes 2023, 14, 1790. https://doi.org/10.3390/genes14091790
Jia X, Xiong Y, Xiong Y, Li D, Yu Q, Lei X, You M, Bai S, Zhang J, Ma X. Identification and Validation of Reference Genes for RT-qPCR Analysis in Reed Canary Grass during Abiotic Stress. Genes. 2023; 14(9):1790. https://doi.org/10.3390/genes14091790
Chicago/Turabian StyleJia, Xuejie, Yi Xiong, Yanli Xiong, Daxu Li, Qinqin Yu, Xiong Lei, Minghong You, Shiqie Bai, Jianbo Zhang, and Xiao Ma. 2023. "Identification and Validation of Reference Genes for RT-qPCR Analysis in Reed Canary Grass during Abiotic Stress" Genes 14, no. 9: 1790. https://doi.org/10.3390/genes14091790
APA StyleJia, X., Xiong, Y., Xiong, Y., Li, D., Yu, Q., Lei, X., You, M., Bai, S., Zhang, J., & Ma, X. (2023). Identification and Validation of Reference Genes for RT-qPCR Analysis in Reed Canary Grass during Abiotic Stress. Genes, 14(9), 1790. https://doi.org/10.3390/genes14091790