


STAT4 Gene Variant rs7574865 Is Associated with Rheumatoid Arthritis Activity and Anti-CCP Levels in the Western but Not in the Southern Population of Mexico


 , , ,
, , ,  , and
, and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Autoantibodies and Laboratory Assessment
2.3. Genotyping of STAT4 Variants (rs7574865 and rs897200)
2.4. Statistics
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bartlett, D.B.; Willis, L.H.; Slentz, C.A.; Hoselton, A.; Kelly, L.; Huebner, J.L.; Kraus, V.B.; Moss, J.; Muehlbauer, M.J.; Spielmann, G.; et al. Ten Weeks of High-Intensity Interval Walk Training Is Associated with Reduced Disease Activity and Improved Innate Immune Function in Older Adults with Rheumatoid Arthritis: A Pilot Study. Arthritis Res. Ther. 2018, 20, 127. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Baruah, C.; Saikia, A.K.; Bose, S. Associative Role of HLA-DRB1 SNP Genotypes as Risk Factors for Susceptibility and Severity of Rheumatoid Arthritis: A North-East Indian Population-Based Study. Int. J. Immunogenet. 2018, 45, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.M.M.T.; Vino, S. Genetic Markers as Therapeutic Target in Rheumatoid Arthritis: A Game Changer in Clinical Therapy? Rheumatol. Int. 2016, 36, 1601–1607. [Google Scholar] [CrossRef]
- Smolen, J.S.; Breedveld, F.C.; Burmester, G.R.; Bykerk, V.; Dougados, M.; Emery, P.; Kvien, T.K.; Navarro-Compán, M.V.; Oliver, S.; Schoels, M.; et al. Treating Rheumatoid Arthritis to Target: 2014 Update of the Recommendations of an International Task Force. Ann. Rheum. Dis. 2016, 75, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Smolen, J.S.; Aletaha, D.; McInnes, I.B. Rheumatoid Arthritis. Lancet 2016, 388, 2023–2038. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Montoya, J.; Alvarez-Nemegyei, J.; Sanin, L.H.; Pérez-Barbosa, L.; Trejo-Valdivia, B.; Santana, N.; Goycochea-Robles, M.V.; Cardiel, M.H.; Riega-Torres, J.; Maradiaga, M.; et al. Association of Regional and Cultural Factors with the Prevalence of Rheumatoid Arthritis in the Mexican Population. J. Clin. Rheumatol. 2015, 21, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Padyukov, L. Genetics of Rheumatoid Arthritis. Semin. Immunopathol. 2022, 44, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Ciccacci, C.; Conigliaro, P.; Perricone, C.; Rufini, S.; Triggianese, P.; Politi, C.; Novelli, G.; Perricone, R.; Borgiani, P. Polymorphisms in STAT-4, IL-10, PSORS1C1, PTPN2 and MIR146A Genes Are Associated Differently with Prognostic Factors in Italian Patients Affected by Rheumatoid Arthritis. Clin. Exp. Immunol. 2016, 186, 157–163. [Google Scholar] [CrossRef]
- Scally, S.W.; Law, S.-C.; Ting, Y.T.; van Heemst, J.; Sokolove, J.; Deutsch, A.J.; Bridie Clemens, E.; Moustakas, A.K.; Papadopoulos, G.K.; van der Woude, D.; et al. Molecular Basis for Increased Susceptibility of Indigenous North Americans to Seropositive Rheumatoid Arthritis. Ann. Rheum. Dis. 2017, 76, 1915–1923. [Google Scholar] [CrossRef]
- Mikhaylenko, D.S.; Nemtsova, M.V.; Bure, I.V.; Kuznetsova, E.B.; Alekseeva, E.A.; Tarasov, V.V.; Lukashev, A.N.; Beloukhova, M.I.; Deviatkin, A.A.; Zamyatnin, A.A. Genetic Polymorphisms Associated with Rheumatoid Arthritis Development and Antirheumatic Therapy Response. Int. J. Mol. Sci. 2020, 21, 4911. [Google Scholar] [CrossRef]
- Bandinelli, F.; Benucci, M.; Salaffi, F.; Manetti, M.; Infantino, M.; Damiani, A.; Manfredi, M.; Grossi, V.; Matucci, A.; Li Gobbi, F.; et al. Do New and Old Biomarkers of Early Undifferentiated Arthritis Correlate with Arthritis Impact Measurement Scales? Clin. Exp. Rheumatol. 2021, 39, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda-Delgado, J.; Rizo-Pinto, A.; Granados-Arriola, J.; Mena-Vela, B.A.; Cetina-Díaz, J.H.; García-Silva, R.; Hernández-Doño, S.; Cruz-Salvatierra, M.A.; Pérez-Tirado, J.M.; Vázquez-Guzmán, C.; et al. Role of HLA-DRB1*04 in the Susceptibility and HLA-DRB1*08 in the Protection for Development of Rheumatoid Arthritis in a Population of Southern Mexico: Brief Report. Clin. Rheumatol. 2020, 39, 2875–2879. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-C.; Huang, C.-M.; Liu, T.-Y.; Wu, N.; Chan, C.-J.; Shih, P.-Y.; Chen, H.-H.; Chen, S.-Y.; Tsai, F.-J. Effects of Human Leukocyte Antigen DRB1 Genetic Polymorphism on Anti-Cyclic Citrullinated Peptide (ANTI-CCP) and Rheumatoid Factor (RF) Expression in Rheumatoid Arthritis (RA) Patients. Int. J. Mol. Sci. 2023, 24, 12036. [Google Scholar] [CrossRef] [PubMed]
- Benucci, M.; Damiani, A.; Li Gobbi, F.; Bandinelli, F.; Infantino, M.; Grossi, V.; Manfredi, M.; Noguier, G.; Meacci, F. Correlation between HLA Haplotypes and the Development of Antidrug Antibodies in a Cohort of Patients with Rheumatic Diseases. Biologics 2018, 12, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.M.; John, P.; Fan, K.H.; Bhatti, A.; Aziz, W.; Ahmed, B.; Feingold, E.; Demirci, F.Y.; Kamboh, M.I. Investigating the GWAS-Implicated Loci for Rheumatoid Arthritis in the Pakistani Population. Dis. Markers 2020, 2020. [Google Scholar] [CrossRef] [PubMed]
- Tizaoui, K.; Terrazzino, S.; Cargnin, S.; Lee, K.H.; Gauckler, P.; Li, H.; Shin, J.I.; Kronbichler, A. The Role of PTPN22 in the Pathogenesis of Autoimmune Diseases: A Comprehensive Review. Semin. Arthritis Rheum. 2021, 51, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Tizaoui, K.; Shin, J.I.; Jeong, G.H.; Yang, J.W.; Park, S.; Kim, J.H.; Hwang, S.Y.; Park, S.J.; Koyanagi, A.; Smith, L. Genetic Polymorphism of PTPN22 in Autoimmune Diseases: A Comprehensive Review. Medicina 2022, 58, 1034. [Google Scholar] [CrossRef]
- Karami, J.; Aslani, S.; Jamshidi, A.; Garshasbi, M.; Mahmoudi, M. Genetic Implications in the Pathogenesis of Rheumatoid Arthritis; an Updated Review. Gene 2019, 702, 8–16. [Google Scholar] [CrossRef]
- Odqvist, L.; Jevnikar, Z.; Riise, R.; Öberg, L.; Rhedin, M.; Leonard, D.; Yrlid, L.; Jackson, S.; Mattsson, J.; Nanda, S.; et al. Genetic Variations in A20 DUB Domain Provide a Genetic Link to Citrullination and Neutrophil Extracellular Traps in Systemic Lupus Erythematosus. Ann. Rheum. Dis. 2019, 78, 1363–1370. [Google Scholar] [CrossRef]
- Plenge, R.M.; Seielstad, M.; Padyukov, L.; Lee, A.T.; Remmers, E.F.; Ding, B.; Liew, A.; Khalili, H.; Chandrasekaran, A.; Davies, L.R.L.; et al. TRAF1-C5 as a Risk Locus for Rheumatoid Arthritis—A Genomewide Study. N. Engl. J. Med. 2007, 357, 1199–1209. [Google Scholar] [CrossRef]
- Kurreeman, F.A.S.; Padyukov, L.; Marques, R.B.; Schrodi, S.J.; Seddighzadeh, M.; Stoeken-Rijsbergen, G.; Van Der Helm-Van Mil, A.H.M.; Allaart, C.F.; Verduyn, W.; Houwing-Duistermaat, J.; et al. A Candidate Gene Approach Identifies the TRAF1/C5 Region as a Risk Factor for Rheumatoid Arthritis. PLoS Med. 2007, 4, 1515–1524. [Google Scholar] [CrossRef]
- Tarakji, I.; Habbal, W.; Monem, F. Association Between STAT4 Rs7574865 Polymorphism and Rheumatoid Arthritis: Debate Unresolved. Open Rheumatol. J. 2018, 12, 172–178. [Google Scholar] [CrossRef]
- Gao, W.; Dong, X.; Yang, Z.; Mao, G.; Xing, W. Association between Rs7574865 Polymorphism in STAT4 Gene and Rheumatoid Arthritis: An Updated Meta-Analysis. Eur. J. Intern. Med. 2020, 71, 101–103. [Google Scholar] [CrossRef]
- Hiz, M.M.; Kiliç, S.; Iik, S.; Ogretmen, Z.; Silan, F. Contribution of the STAT4 Rs7574865 Gene Polymorphism to the Susceptibility to Autoimmune Thyroiditis in Healthy Turk Population and Psoriatic Subgroups. Cent. Eur. J. Immunol. 2015, 40, 437–441. [Google Scholar] [CrossRef]
- Jahreis, S.; Kuhn, S.; Madaj, A.M.; Bauer, M.; Polte, T. Mold Metabolites Drive Rheumatoid Arthritis in Mice via Promotion of IFN-γ- and IL-17-Producing T Cells. Food Chem. Toxicol. 2017, 109, 405–413. [Google Scholar] [CrossRef]
- Banerjee, S.; Biehl, A.; Gadina, M.; Hasni, S.; Schwartz, D.M. JAK–STAT Signaling as a Target for Inflammatory and Autoimmune Diseases: Current and Future Prospects. Drugs 2017, 77, 521–546. [Google Scholar] [CrossRef]
- Salaffi, F.; Giacobazzi, G.; Carlo, M.D. Pain and the JAK-STAT Pathway. Pain Res. Manag. 2018, 2018, 15. [Google Scholar]
- Patel, Z.; Lu, X.; Miller, D.; Forney, C.R.; Lee, J.; Lynch, A.; Parks, L.; Magnusen, A.F.; Chen, X.; Pujato, M.; et al. A Plausibly Causal Functional Lupus-Associated Risk Variant in the STAT1–STAT4 Locus. Hum. Mol. Genet. 2018, 27, 2392–2404. [Google Scholar] [CrossRef] [PubMed]
- Hagberg, N.; Joelsson, M.; Leonard, D.; Reid, S.; Eloranta, M.L.; Mo, J.; Nilsson, M.K.; Syvänen, A.C.; Bryceson, Y.T.; Rönnblom, L. The STAT4 SLE Risk Allele Rs7574865[T] Is Associated with Increased IL-12-Induced IFN-γ Production in t Cells from Patients with SLE. Ann. Rheum. Dis. 2018, 77, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Lamana, A.; López-Santalla, M.; Castillo-González, R.; Ortiz, A.M.; Martín, J.; García-Vicuña, R.; González-Álvaro, I. The Minor Allele of Rs7574865 in the STAT4 Gene Is Associated with Increased mRNA and Protein Expression. PLoS ONE 2015, 10, e142683. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Yang, Z.; Du, L.; Jiang, Z.; Shu, Q.; Chen, Y.; Li, F.; Zhou, Q.; Ohno, S.; Chen, R.; et al. Identification of a Susceptibility Locus in STAT4 for Behçet’s Disease in Han Chinese in a Genome-Wide Association Study. Arthritis Rheum. 2012, 64, 4104–4113. [Google Scholar] [CrossRef] [PubMed]
- Sabri, A.; Grant, A.V.; Cosker, K.; El Azbaoui, S.; Abid, A.; Abderrahmani Rhorfi, I.; Souhi, H.; Janah, H.; Alaoui-Tahiri, K.; Gharbaoui, Y.; et al. Association Study of Genes Controlling IL-12-Dependent IFN-γ Immunity: STAT4 Alleles Increase Risk of Pulmonary Tuberculosis in Morocco. J. Infect. Dis. 2014, 210, 611–618. [Google Scholar] [CrossRef]
- Malemud, C.J. The Role of the JAK/STAT Signal Pathway in Rheumatoid Arthritis. Ther. Adv. Musculoskelet. Dis. 2018, 10, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Mondal, H.; Lotfollahzadeh, S. Hematocrit. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Cappelli, L.C.; Thomas, M.A.; Bingham, C.O.; Shah, A.A.; Darrah, E. Immune Checkpoint Inhibitor–Induced Inflammatory Arthritis as a Model of Autoimmune Arthritis. Immunol. Rev. 2020, 294, 106–123. [Google Scholar] [CrossRef] [PubMed]
- Smolen, J.S.; Aletaha, D.; Barton, A.; Burmester, G.R.; Emery, P.; Firestein, G.S.; Kavanaugh, A.; McInnes, I.B.; Solomon, D.H.; Strand, V.; et al. Rheumatoid Arthritis. Nat. Rev. Dis. Primers 2018, 4, 18001. [Google Scholar] [CrossRef]
- Stahl, E.A.; Raychaudhuri, S.; Remmers, E.F.; Xie, G.; Eyre, S.; Thomson, B.P.; Li, Y.; Kurreeman, F.A.S.; Zhernakova, A.; Hinks, A.; et al. Genome-Wide Association Study Meta-Analysis Identifies Seven New Rheumatoid Arthritis Risk Loci. Nat. Genet. 2010, 42, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Viatte, S.; Plant, D.; Raychaudhuri, S. Genetics and Epigenetics of Rheumatoid Arthritis. Nat. Rev. Rheumatol. 2013, 9, 141–153. [Google Scholar] [CrossRef]
- Chaouali, M.; Fernandes, V.; Ghazouani, E.; Pereira, L.; Kochkar, R. Association of STAT4, TGFβ1, SH2B3 and PTPN22 Polymorphisms with Autoimmune Hepatitis. Exp. Mol. Pathol. 2018, 105, 279–284. [Google Scholar] [CrossRef]
- Karlson, E.W.; Deane, K. Environmental and Gene-Environment Interactions and Risk of Rheumatoid Arthritis. Rheum. Dis. Clin. N. Am. 2012, 38, 405–426. [Google Scholar] [CrossRef]
- Alpízar-Rodríguez, D.; Finckh, A. Environmental Factors and Hormones in the Development of Rheumatoid Arthritis. Semin. Immunopathol. 2017, 39, 461–468. [Google Scholar] [CrossRef]
- Sato, K.; Suematsu, A.; Okamoto, K.; Yamaguchi, A.; Morishita, Y.; Kadono, Y.; Tanaka, S.; Kodama, T.; Akira, S.; Iwakura, Y.; et al. Th17 Functions as an Osteoclastogenic Helper T Cell Subset That Links T Cell Activation and Bone Destruction. J. Exp. Med. 2006, 203, 2673–2682. [Google Scholar] [CrossRef] [PubMed]
- Simon, L.S.; Taylor, P.C.; Choy, E.H.; Sebba, A.; Quebe, A.; Knopp, K.L.; Porreca, F. The Jak/STAT Pathway: A Focus on Pain in Rheumatoid Arthritis. Semin. Arthritis Rheum. 2021, 51, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Ma, H.; Zhang, H.; Deng, C.; Xin, P. Recent Advances on Signaling Pathways and Their Inhibitors in Rheumatoid Arthritis. Clin. Immunol. 2021, 230, 108793. [Google Scholar] [CrossRef] [PubMed]
- Deane, K.D.; Demoruelle, M.K.; Kelmenson, L.B.; Kuhn, K.A.; Norris, J.M.; Holers, V.M. Genetic and Environmental Risk Factors for Rheumatoid Arthritis. Best Pract. Res. Clin. Rheumatol. 2017, 31, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yin, J. HLA Risk Alleles and Gut Microbiome in Ankylosing Spondylitis and Rheumatoid Arthritis. Best Pract. Res. Clin. Rheumatol. 2019, 33, 101499. [Google Scholar] [CrossRef] [PubMed]
- Rubi-Castellanos, R.; Martínez-Cortés, G.; Muñoz-Valle, J.F.; González-Martín, A.; Cerda-Flores, R.M.; Anaya-Palafox, M.; Rangel-Villalobos, H. Pre-Hispanic Mesoamerican Demography Approximates the Present-Day Ancestry of Mestizos throughout the Territory of Mexico. Am. J. Phys. Anthropol. 2009, 139, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Estrada, A.; Gignoux, C.R.; Carlos Fernández-López, J.; Zakharia, F.; Sikora, M.; Contreras, A.V.; Acuña-Alonzo, V.; Sandoval, K.; Eng, C.; Romero-Hidalgo, S.; et al. Human genetics. The genetics of Mexico recapitulates Native American substructure and affects biomedical traits. Science 2014, 344, 1280–1285. [Google Scholar]
- Cahua-Pablo, J.Á.; Cruz, M.; Tello-Almaguer, P.V.; del Alarcón-Romero, L.C.; Parra, E.J.; Villerías-Salinas, S.; Valladares-Salgado, A.; Tello-Flores, V.A.; Méndez-Palacios, A.; Pérez-Macedonio, C.P.; et al. Analysis of Admixture Proportions in Seven Geographical Regions of the State of Guerrero, Mexico. Am. J. Hum. Biol. 2017, 29, e23032. [Google Scholar] [CrossRef]
- Aguilar-Velázquez, J.A.; Martínez-Cortés, G.; Inclán-Sánchez, A.; Favela-Mendoza, A.F.; Velarde-Félix, J.S.; Rangel-Villalobos, H. Forensic Parameters and Admixture in Mestizos from Five Geographic Regions of Mexico Based on 20 Autosomal STRs (Powerplex 21 System). Int. J. Leg. Med. 2018, 132, 1293–1296. [Google Scholar] [CrossRef]
- Martínez-Cortés, G.; Salazar-Flores, J.; Gabriela Fernández-Rodríguez, L.; Rubi-Castellanos, R.; Rodríguez-Loya, C.; Velarde-Félix, J.S.; Franciso Muñoz-Valle, J.; Parra-Rojas, I.; Rangel-Villalobos, H. Admixture and Population Structure in Mexican-Mestizos Based on Paternal Lineages. J. Hum. Genet. 2012, 57, 568–574. [Google Scholar] [CrossRef]
- Sohail, M.; Palma-Martínez, M.J.; Chong, A.Y.; Quinto-Cortés, C.D.; Barberena-Jonas, C.; Medina-Muñoz, S.G.; Ragsdale, A.; Delgado-Sánchez, G.; Cruz-Hervert, L.P.; Ferreyra-Reyes, L.; et al. Mexican Biobank Advances Population and Medical Genomics of Diverse Ancestries. Nature 2023, 622, 775–783. [Google Scholar] [CrossRef]
- Shen, L.; Liu, R.; Zhang, H.; Huang, Y.; Sun, R.; Tang, P. Replication Study of STAT4 Rs7574865 G/T Polymorphism and Risk of Rheumatoid Arthritis in a Chinese Population. Gene 2013, 526, 259–264. [Google Scholar] [CrossRef]
- Krasselt, M.; Baerwald, C. Sex, Symptom Severity, and Quality of Life in Rheumatology. Clin. Rev. Allergy Immunol. 2019, 56, 346–361. [Google Scholar] [CrossRef]
- Fernández-Torres, J.; Zamudio-Cuevas, Y.; Martínez-Nava, G.A.; Aztatzi-Aguilar, O.G.; Sierra-Vargas, M.P.; Lozada-Pérez, C.A.; Suárez-Ahedo, C.; Landa-Solís, C.; Olivos-Meza, A.; Del Razo, L.M.; et al. Impact of Cadmium Mediated by Tobacco Use in Musculoskeletal Diseases. Biol. Trace Elem. Res. 2022, 200, 2008–2015. [Google Scholar] [CrossRef]
- Hedström, A.K.; Rönnelid, J.; Klareskog, L.; Alfredsson, L. Complex Relationships of Smoking, HLA–DRB1 Genes, and Serologic Profiles in Patients With Early Rheumatoid Arthritis: Update From a Swedish Population-Based Case–Control Study. Arthritis Rheumatol. 2019, 71, 1504–1511. [Google Scholar] [CrossRef] [PubMed]
- Alpízar-Rodríguez, D.; Finckh, A.; Gilbert, B. The Role of Nutritional Factors and Intestinal Microbiota in Rheumatoid Arthritis Development. Nutrients 2021, 13, 96. [Google Scholar] [CrossRef] [PubMed]
- Shen, R.; Ren, X.; Jing, R.; Shen, X.; Chen, J. Rheumatoid Factor, Anti-Cyclic Citrullinated Peptide Antibody, C-Reactive Protein, and Erythrocyte Sedimentation Rate for the Clinical Diagnosis of Rheumatoid Arthritis. Lab. Med. 2015, 46, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Shamanna, P.; Arundhathi, S.; Kumar, R.P.; Vanishree, V. A Retrospective Case-Control Hospital Based Study on the Diagnostic Utility of Anti-Cyclic Citrullinated Peptide Antibodies in Comparison with Rheumatoid Factor, C-Reactive Protein and Erythrocyte Sedimentation Rate as a Diagnostic Inflammatory Biomarker. Trop. J. Pathol. Microbiol. 2019, 5, 437–442. [Google Scholar] [CrossRef]
- Yang, D.; Li, H.; Zhang, S.; Yang, G.; Liu, S.; Liu, Z. Clinical Analysis of Blood Markers and Imaging for Diagnosis and Prognosis of Rheumatoid Arthritis. Rheumatology 2020, 1–11. [Google Scholar] [CrossRef]
- de Durán-Avelar, M.J.; Vibanco-Pérez, N.; Hernández-Pacheco, R.R.; Castro-Zambrano, A.d.C.; Ortiz-Martínez, L.; Zambrano-Zaragoza, J.F. STAT4 Rs7574865 G/T Polymorphism Is Associated with Rheumatoid Arthritis and Disease Activity, but Not with Anti-CCP Antibody Levels in a Mexican Population. Clin. Rheumatol. 2016, 35, 2909–2914. [Google Scholar] [CrossRef] [PubMed]
- Settin, A.; Salama, A.; Elshazli, R. Signal Transducer and Activator of Transcription 4 (STAT4) G>T Gene Polymorphism in Egyptian Cases with Rheumatoid Arthritis. Hum. Immunol. 2014, 75, 863–866. [Google Scholar] [CrossRef]
- Ebrahimiyan, H.; Mostafaei, S.; Aslani, S.; Jamshidi, A.; Mahmoudi, M. Studying the Association between STAT4 Gene Polymorphism and Susceptibility to Rheumatoid Arthritis Disease: An Updated Meta-Analysis. Iran. J. Immunol. 2019, 16, 71–83. [Google Scholar] [CrossRef]
- Gu, E.; Lu, J.; Xing, D.; Chen, X.; Xie, H.; Liang, J.; Li, L. Rs7574865 Polymorphism in Signal Transducers and Activators of Transcription 4 Gene and Rheumatoid Arthritis: An Updated Meta-Analysis of 28 Case-Control Comparisons. Int. J. Rheum. Dis. 2015, 18, 3–16. [Google Scholar] [CrossRef]
- Wu, S.; Wang, M.; Wang, Y.; Zhang, M.; He, J.Q. Polymorphisms of the STAT4 Gene in the Pathogenesis of Tuberculosis. Biosci. Rep. 2018, 38, BSR20180498. [Google Scholar] [CrossRef] [PubMed]
- Cénit, M.C.; Márquez, A.; Cordero-Coma, M.; Gorroño-Echebarría, M.B.; Fonollosa, A.; Adán, A.; Martínez-Berriotxoa, A.; Valle, D.D.; Pato, E.; Blanco, R.; et al. No Evidence of Association between Common Autoimmunity STAT4 and IL23R Risk Polymorphisms and Non-Anterior Uveitis. PLoS ONE 2013, 8, 1–6. [Google Scholar] [CrossRef] [PubMed]
- El-Lebedy, D.; Raslan, H.; Ibrahim, A.; Ashmawy, I.; El-Aziz, S.A.; Mohammed, A.M. Association of STAT4 Rs7574865 and PTPN22 Rs2476601 Polymorphisms with Rheumatoid Arthritis and Non-Systemically Reacting Antibodies in Egyptian Patients. Clin. Rheumatol. 2017, 36, 1981–1987. [Google Scholar] [CrossRef] [PubMed]
- Mirkazemi, S.; Akbarian, M.; Jamshidi, A.R.; Mansouri, R.; Ghoroghi, S.; Salimi, Y.; Tahmasebi, Z.; Mahmoudi, M. Association of STAT4 Rs7574865 with Susceptibility to Systemic Lupus Erythematosus in Iranian Population. Inflammation 2013, 36, 1548–1552. [Google Scholar] [CrossRef] [PubMed]
- Shaul, O. How Introns Enhance Gene Expression. Int. J. Biochem. Cell Biol. 2017, 91, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, N.; Ansari, A. Enhancement of Transcription by a Splicing-Competent Intron Is Dependent on Promoter Directionality. PLoS Genet. 2016, 12, e1006047. [Google Scholar] [CrossRef] [PubMed]
- Howe, F.S.; Fischl, H.; Murray, S.C.; Mellor, J. Is H3K4me3 Instructive for Transcription Activation? BioEssays 2017, 39, e201600095. [Google Scholar] [CrossRef]
- Sakabe, N.J.; Savic, D.; Nobrega, M.A. Transcriptional Enhancers in Development and Disease. Genome Biol. 2012, 13, 238. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
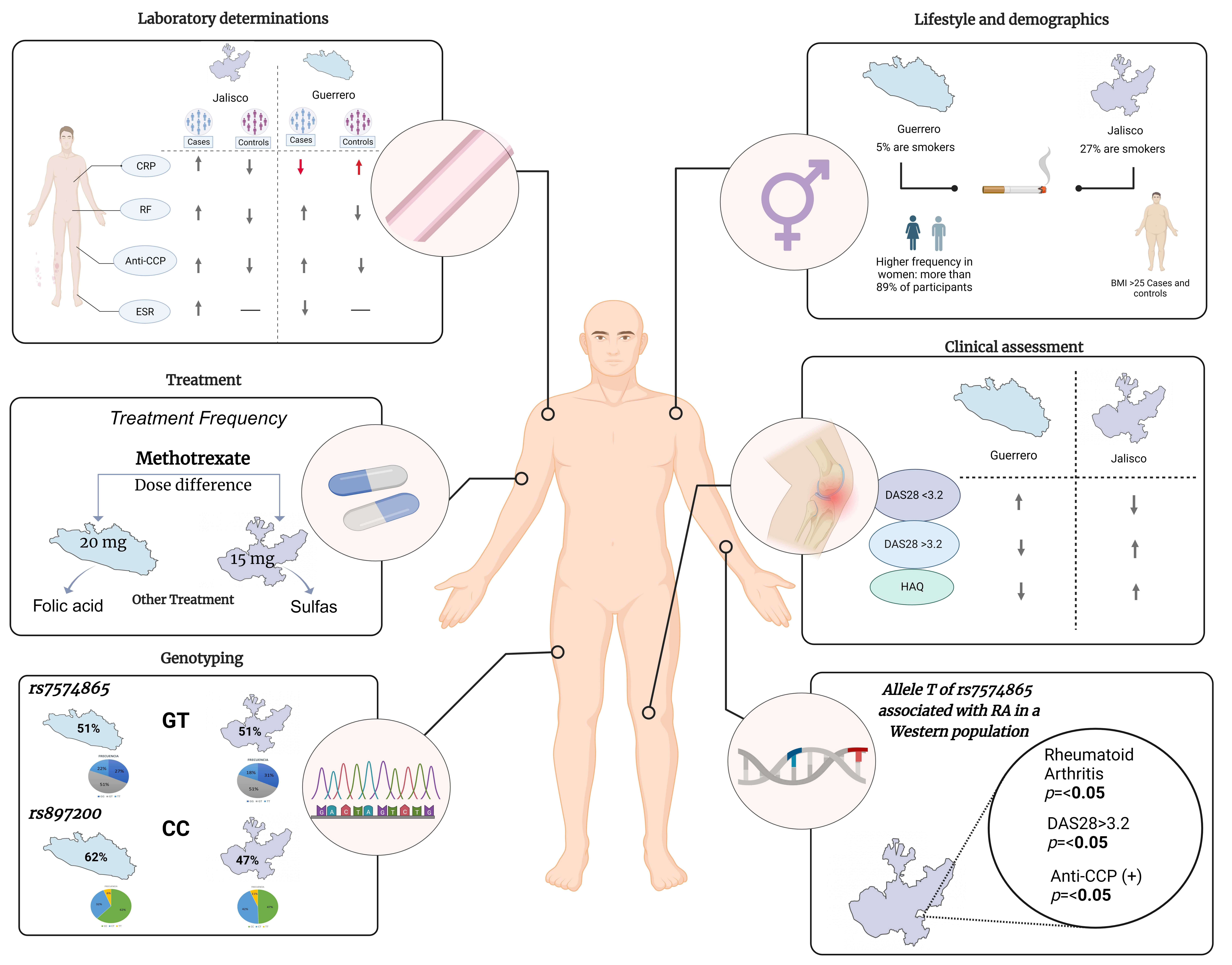
| Variable | Southern Controls (n = 120) | Southern Cases (n = 120) | * p | Western Controls (n = 116) | Western Cases (n = 120) | * p |
|---|---|---|---|---|---|---|
| Demographics | ||||||
| Age (years) f | 46 (35–56) | 44 (36–58) | 0.770 b | 38 (30–50) | 51 (42–61) | <0.001 **b |
| Gender | ||||||
| Female/Male g | 92.5 (111)/7.5 (9) | 91.7 (110)/8.3 (10) | 0.811 c | 89.7 (104)/13.3 (16) | 82.8 (96)/17.2 (20) | 0.404 c |
| Family history of RA g | - | 25 (30) | - | - | 60.8 (73) | <0.001 **c |
| Clinical assessment | ||||||
| Time of disease evolution (years) d,f | - | 6 (3–11.75) | - | - | 6 (2–14) | 0.930 b |
| DAS28 d,g | ||||||
| Remission <2.6 g | - | 17.5 (21) | - | - | 12.5 (15) | <0.001 **c |
| Low activity ≥2.6 to < 3.2 g | - | 41.7 (50) | - | - | 10.8 (13) | <0.001 **c |
| Moderate activity ≥3.2 to <5.1 g | - | 28.3 (34) | - | - | 47.5 (57) | <0.001 **c |
| High activity >5.1 g | - | 12.5 (15) | - | - | 29.2 (35) | <0.001 **c |
| HAQ (0–3) d,f | - | 0.2050 (0.04–0.6050) | - | - | 1.1150 (0.3725–1.87) | <0.001 **b |
| VAS d,f | - | 40 (20–50) | - | - | 40 (20–50) | 0.419 b |
| ESR (mm/h) d,e | - | 32.18 ± 14.374 | - | - | 36.61 ± 17.082 | 0.030 *a |
| CRP (mg/L) f | 26.50 (3–58) | 15.75 (2.3–298.2) | 0.002 b | 0.2650 (0.0950–0.9485) | 1.53 (0.34–5.68) | <0.001 **b |
| RF (IU/mL) f | 11.75 (0–326) | 181.45 (0–300) | <0.001 **b | 8 (6.30–9.48) | 101.10 (43.8–288.9) | <0.001 **b |
| Anti-CCP (U/mL) e | 0.00 (0–100) | 148.42 (0–900) | <0.001 **b | 8.04 ± 8.736 | 253.67 ± 179.21 | <0.001 **a |
| Treatment | ||||||
| Corticosteroids d,g | - | 42.5 (51) | - | - | 13.3 (16) | <0.001 **c |
| DMARDs d,g | ||||||
| Methotrexate d,g | - | 81.7 (98) | - | - | 88.3 (106) | <0.001 **c |
| Chloroquine d,g | - | 42.5 (51) | - | - | 13.3 (16) | <0.001 **c |
| Sulfasalazine d,g | - | 24.2 (29) | - | - | 72.5 (87) | <0.001 **c |
| Southern (Control/Cases) | |||||||
|---|---|---|---|---|---|---|---|
| Clinical Assessment | Genotypes | OR (95% CI) | OR (95% CI) | ||||
| STAT4: rs7574865 T > G | GG | GT | TT | GT | p | TT | p |
| DAS28 (<3.20) | 44/18 | 56/30 | 20/12 | 1.310 (0.647–2.651) | 0.453 | 1.467 (0.595–3.613) | 0.700 |
| DAS28 (>3.20) | 44/14 | 56/30 | 20/15 | 1.684 (0.798–3.554) | 0.169 | 2.357 (0.958–5.797) | 0.059 |
| RF-negative (<20 IU/mL) | 32/4 | 47/7 | 16/3 | 1.191 (0.322–4.407) | 0.792 | 1.50 (0.299–7.525) | 0.620 |
| RF-positive (>20 IU/mL) | 12/28 | 9/53 | 4/24 | 2.524 (0.949–6.712) | 0.059 | 2.571 (0.732–9.03) | 0.132 |
| Anti-CCP-negative (<5 U/mL) | 39/3 | 52/5 | 18/2 | 1.250 (0.282–5.548) | 0.768 | 1.444 (0.222–9.413) | 0.699 |
| Anti-CCP-positive (>5 U/mL) | 5/29 | 4/55 | 2/25 | 2.371 (0.591–9.514) | 0.213 | 2.155 (0.384–12.094) | 0.374 |
| STAT4: rs897200 T > C | CC | CT | TT | CT | p | TT | p |
| DAS28 (<3.20) | 69/34 | 40/22 | 11/4 | 1.116 (0.575–2.166) | 0.745 | 0.738 (2.19–2.489) | 0.623 |
| DAS28 (>3.20) | 69/41 | 40/16 | 11/3 | 0.673 (0.335–1.351) | 0.264 | 0.459 (0.121–1.742) | 0.243 |
| RF-negative (<20 IU/mL) | 56/6 | 31/7 | 8/1 | 2.108 (0.651–6.827) | 0.206 | 1.167 (0.124–10.99) | 0.892 |
| RF-positive (>20 IU/mL) | 13/69 | 9/31 | 3/6 | 0.649 (0.251–1.677) | 0.37 | 0.377 (0.83–1.701) | 0.190 |
| Anti-CCP-negative (<5 U/mL) | 64/7 | 35/4 | 10/0 | 1.045 (0.286–3.818) | 0.947 | 0.410 (0.022–7.716) | 0.298 |
| Anti-CCP-positive (>5 U/mL) | 5/68 | 5/34 | 1/7 | 0.50 (0.135–1.846) | 0.291 | 0.515 (0.052–5.050) | 0.562 |
| Western (control/cases) | |||||||
| Clinical Assessment | Genotypes | OR (95% CI) | OR (95% CI) | ||||
| STAT4: rs7574865 T > G | GG | GT | TT | GT | p | TT | p |
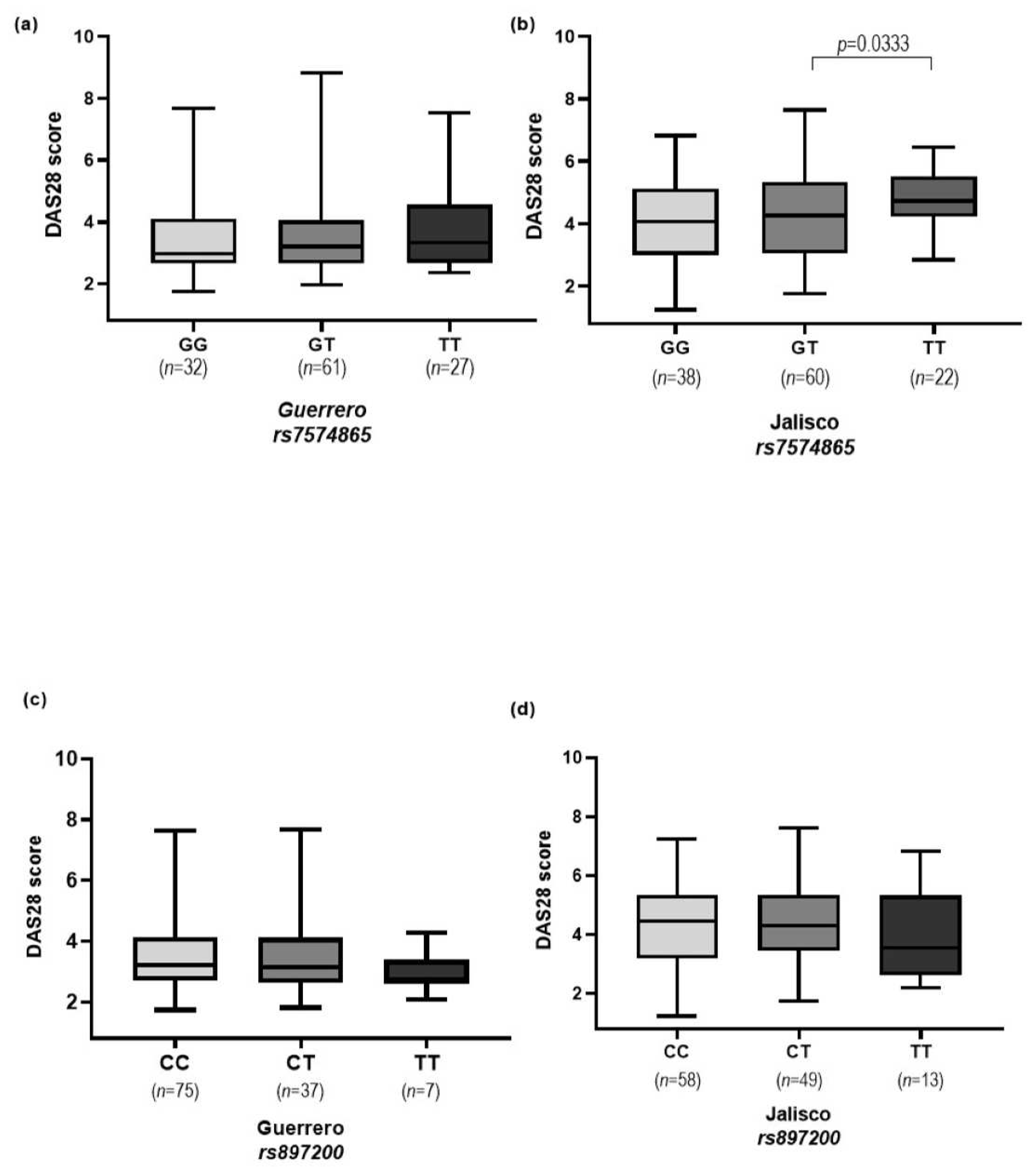
| DAS28 (<3.20) | 60/6 | 45/14 | 11/1 | 3.111 (1.109–8.728) | 0.025 * | 0.909 (0.099–8.307) | 0.932 |
| DAS28 (>3.20) | 60/22 | 45/40 | 11/16 | 2.424 (1.268–4.635) | 0.006 * | 3.967 (1.597–9.854) | 0.002 * |
| RF-negative (<20 IU/mL) | 37/4 | 27/8 | 8/3 | 2.741 (0.748–10.044) | 0.118 | 3.469 (0.646–18.625) | 0.130 |
| RF-positive (>20 IU/mL) | 2/22 | 0/46 | 0/14 | 10.333 (0.476–224.342) | 0.046 | 3.222 (0.144–72.049) | 0.267 |
| Anti-CCP-negative (<5 U/mL) | 23/2 | 19/0 | 3/0 | 0.241 (0.011–5.324) | 0.206 | 1.343 (0.053–34.198) | 0.611 |
| Anti-CCP-positive (>5 U/mL) | 37/26 | 26/54 | 8/17 | 2.956 (1.489–5.869) | 0.001 ** | 3.024 (1.137–8.046) | 0.023 * |
| STAT4: rs897200 T > C | CC | CT | TT | CT | p | TT | p |
| DAS28 (<3.20) | 48/14 | 51/6 | 17/5 | 0.403 (0.143–1.135) | 0.078 | 1.008 (0.316–3.221) | 0.988 |
| DAS28 (>3.20) | 48/42 | 51/40 | 17/7 | 0.896 (0.499–1.610) | 0.714 | 0.471 (0.178–1.245) | 0.123 |
| RF-negative (<20 IU/mL) | 24/7 | 37/5 | 11/3 | 0.463 (0.132–1.629) | 0.223 | 0.935 (0.203–4.315) | 0.931 |
| RF-positive (>20 IU/mL) | 0/46 | 2/41 | 0/9 | 0.178 (0.008–3.826) | 0.139 | 0.204 (0.004–10.951) | 1.000 |
| Anti-CCP-negative (<5 U/mL) | 25/3 | 13/0 | 7/0 | 0.270 (0.013–5.617) | 0.220 | 0.486 (0.022–10.495) | 0.365 |
| Anti-CCP-positive (>5 U/mL) | 23/53 | 38/46 | 10/12 | 0.525 (0.274–1.008) | 0.051 | 0.521 (0.197–1.376) | 0.184 |
| Southern | |||||
|---|---|---|---|---|---|
| Genotype | RA (n = 120) % (n) | Controls (n = 120) % (n) | OR | 95% IC | p-Value |
| STAT4: rs7574865 T > G | |||||
| GG 1 | 26.7 (32) | 36.7 (44) | Reference | ||
| GT 2 | 50.8 (61) | 46.7 (56) | 1.498 | (0.837–2.681) | 0.172 |
| TT 3 | 22.5 (27) | 16.7 (20) | 1.856 | (0.889–3.875) | 0.097 |
| GT + TT 4 | 73.3 (88) | 63.4 (76) | 1.592 | (0.919–2.757) | 0.095 |
| GG + GT 5 | 77.5 (93) | 83.4 (100) | Reference | ||
| TT 3 | 22.5 (27) | 16.7 (20) | 1.452 | (0.763–2.763) | 0.254 |
| G 6 | 52.1 (125) | 60.05 (144) | Reference | ||
| T 7 | 47.9 (115) | 40.05 (96) | 1.380 | (0.961–1.981) | 0.080 |
| STAT4: rs897200 T > C | |||||
| CC 1 | 62.5 (75) | 57.5 (69) | Reference | ||
| CT 2 | 31.7 (38) | 33.3 (40) | 0.874 | (0.504–1.517) | 0.632 |
| TT 3 | 5.8 (7) | 9.2 (11) | 0.585 | (0.215–1.595) | 0.291 |
| CT + TT 4 | 37.5 (45) | 42.5 (51) | 0.812 | (0.484–1.362) | 0.429 |
| CC + CT 5 | 94.2 (113) | 90.8 (109) | Reference | ||
| TT 3 | 5.8 (7) | 9.2 (11) | 0.614 | (0.230–1.641) | 0.326 |
| C 6 | 78.3 (188) | 74.1 (178) | Reference | ||
| T 7 | 21.6 (26) | 25.9 (62) | 0.794 | (0.521–1.211) | 0.283 |
| Western | |||||
| Genotype | RA (n = 120) %(n) | Controls (n = 116) %(n) | OR | 95% IC | p-Value |
| STAT4: rs7574865 T > G | |||||
| GG 1 | 30.8 (37) | 51.7 (60) | Reference | ||
| GT 2 | 50.8 (61) | 38.8 (45) | 2.198 | (1.253–3.857) | 0.005 * |
| TT 3 | 18.3 (22) | 9.5 (11) | 3.243 | (1.412–7.450) | 0.004 * |
| GT + TT 4 | 69.1 (83) | 48.3 (56) | 2.403 | (1.412–4.090) | 0.001 * |
| GG + GT 5 | 81.6 (98) | 90.5 (105) | Reference | ||
| TT 3 | 18.3 (22) | 9.5 (11) | 2.143 | (0.988–4.648) | 0.05 |
| G 6 | 56.2 (135) | 71.1 (165) | Reference | ||
| T 7 | 43.7 (105) | 28.9 (67) | 1.915 | (1.308–2.806) | 0.005 * |
| STAT4: rs897200 T > C | |||||
| CC 1 | 47.5 (57) | 40.5 (47) | Reference | ||
| CT 2 | 41.7 (50) | 44.8 (52) | 0.793 | (0.459–1.371) | 0.405 |
| TT 3 | 10.8 (13) | 14.7 (17) | 0.631 | (0.278–1.430) | 0.267 |
| CT + TT 4 | 52.5 (63) | 59.5 (69) | 0.753 | (0.450–1.261) | 0.280 |
| CC + CT 5 | 89.2 (107) | 85.3 (99) | Reference | ||
| TT 3 | 10.8 (13) | 14.7 (17) | 0.708 | (0.327–1.531) | 0.378 |
| C 6 | 68.3 (164) | 62.9 (146) | Reference | ||
| T 7 | 31.7 (76) | 37.1 (86) | 0.787 | (0.538–1.151) | 0.216 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bravo-Villagra, K.M.; Muñoz-Valle, J.F.; Baños-Hernández, C.J.; Cerpa-Cruz, S.; Navarro-Zarza, J.E.; Parra-Rojas, I.; Aguilar-Velázquez, J.A.; García-Arellano, S.; López-Quintero, A. STAT4 Gene Variant rs7574865 Is Associated with Rheumatoid Arthritis Activity and Anti-CCP Levels in the Western but Not in the Southern Population of Mexico. Genes 2024, 15, 241. https://doi.org/10.3390/genes15020241
Bravo-Villagra KM, Muñoz-Valle JF, Baños-Hernández CJ, Cerpa-Cruz S, Navarro-Zarza JE, Parra-Rojas I, Aguilar-Velázquez JA, García-Arellano S, López-Quintero A. STAT4 Gene Variant rs7574865 Is Associated with Rheumatoid Arthritis Activity and Anti-CCP Levels in the Western but Not in the Southern Population of Mexico. Genes. 2024; 15(2):241. https://doi.org/10.3390/genes15020241
Chicago/Turabian StyleBravo-Villagra, Karla Mayela, José Francisco Muñoz-Valle, Christian Johana Baños-Hernández, Sergio Cerpa-Cruz, José Eduardo Navarro-Zarza, Isela Parra-Rojas, José Alonso Aguilar-Velázquez, Samuel García-Arellano, and Andres López-Quintero. 2024. "STAT4 Gene Variant rs7574865 Is Associated with Rheumatoid Arthritis Activity and Anti-CCP Levels in the Western but Not in the Southern Population of Mexico" Genes 15, no. 2: 241. https://doi.org/10.3390/genes15020241
APA StyleBravo-Villagra, K. M., Muñoz-Valle, J. F., Baños-Hernández, C. J., Cerpa-Cruz, S., Navarro-Zarza, J. E., Parra-Rojas, I., Aguilar-Velázquez, J. A., García-Arellano, S., & López-Quintero, A. (2024). STAT4 Gene Variant rs7574865 Is Associated with Rheumatoid Arthritis Activity and Anti-CCP Levels in the Western but Not in the Southern Population of Mexico. Genes, 15(2), 241. https://doi.org/10.3390/genes15020241