
Identification of 4CL Genes in Desert Poplars and Their Changes in Expression in Response to Salt Stress

Abstract
:1. Introduction
2. Materials and Methods
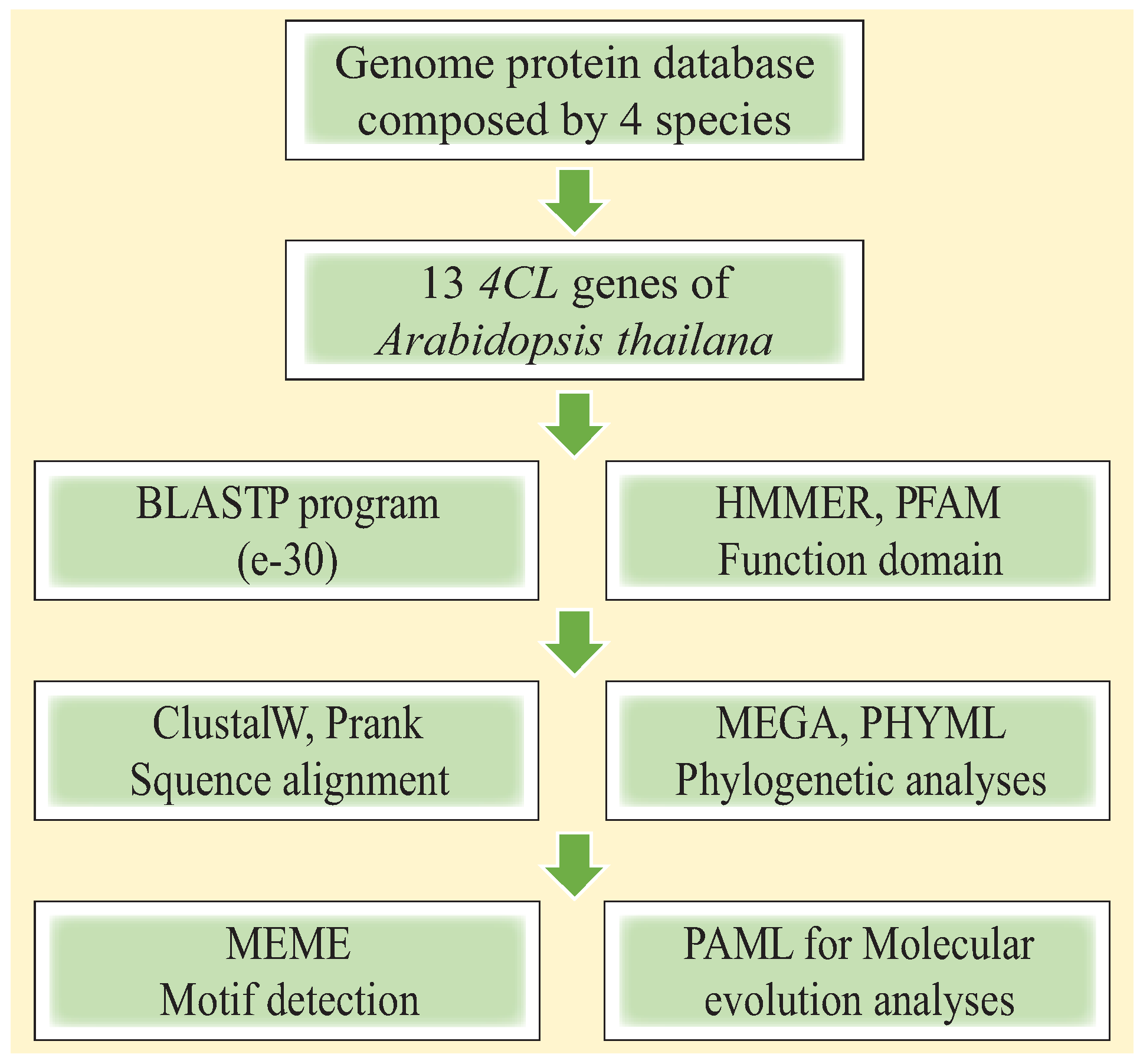
2.1. Gene Sequence Collection and Identification of 4CL Genes
2.2. Phylogenetic Analysis
2.3. Molecular Evolution Analysis

2.4. Expression of 4CL Genes in Salt Stress and Control Conditions
3. Results and Discussion
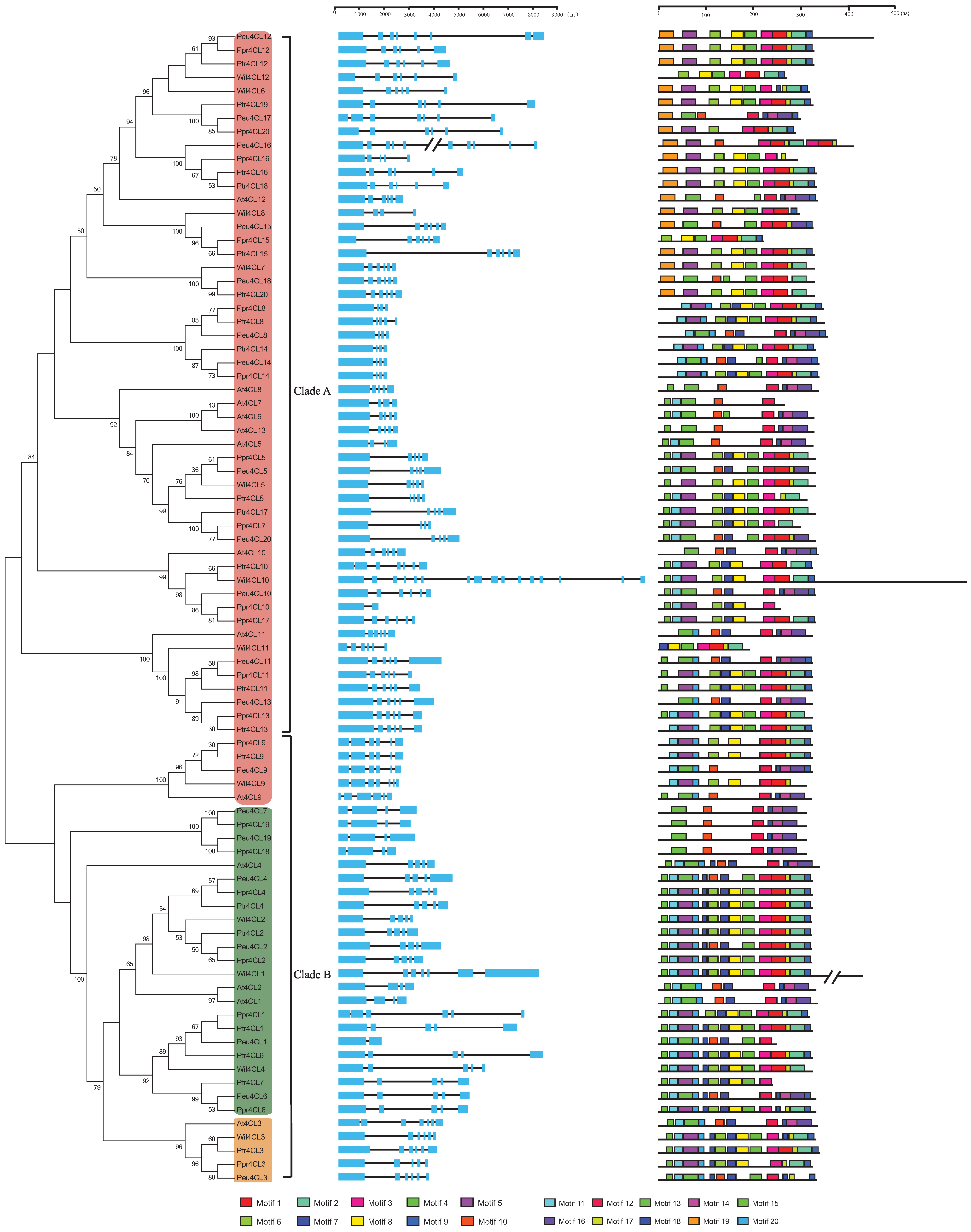
3.1. Identification of the 4CL Genes from Four Species of Salicaceae
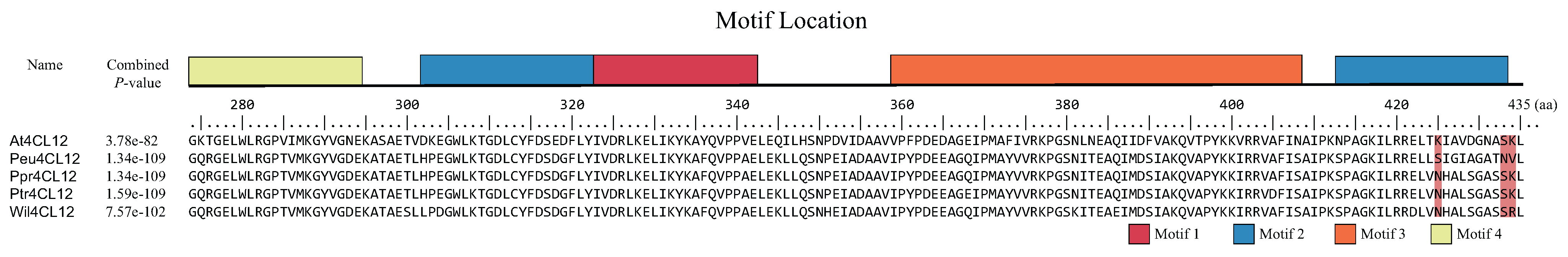
3.2. Gene Structure and Motif Identification of 4CL Genes in Poplars


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organisms | 4CL groups | Total 4CLs | ||
|---|---|---|---|---|
| Class I | Class II | Class-4CL like | ||
| Arabidopsis thaliana | 3 | 1 | 9 | 13 |
| Populus trichocarpa | 5 | 1 | 14 | 20 |
| Populus pruinosa | 4 | 1 | 15 | 20 |
| Populus euphratica | 4 | 1 | 15 | 20 |
| Salix suchowensis | 3 | 1 | 8 | 12 |
3.3. Phylogenetic Analysis of 4CL Genes
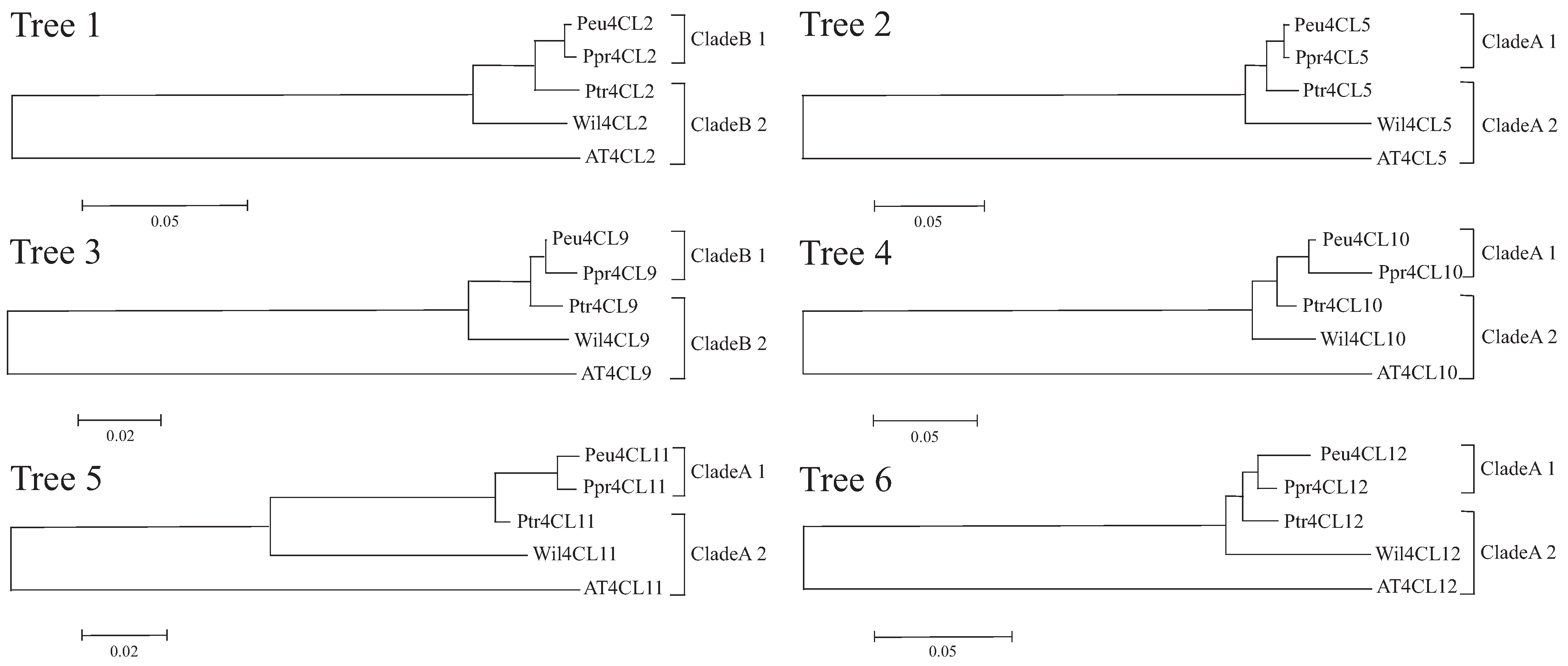
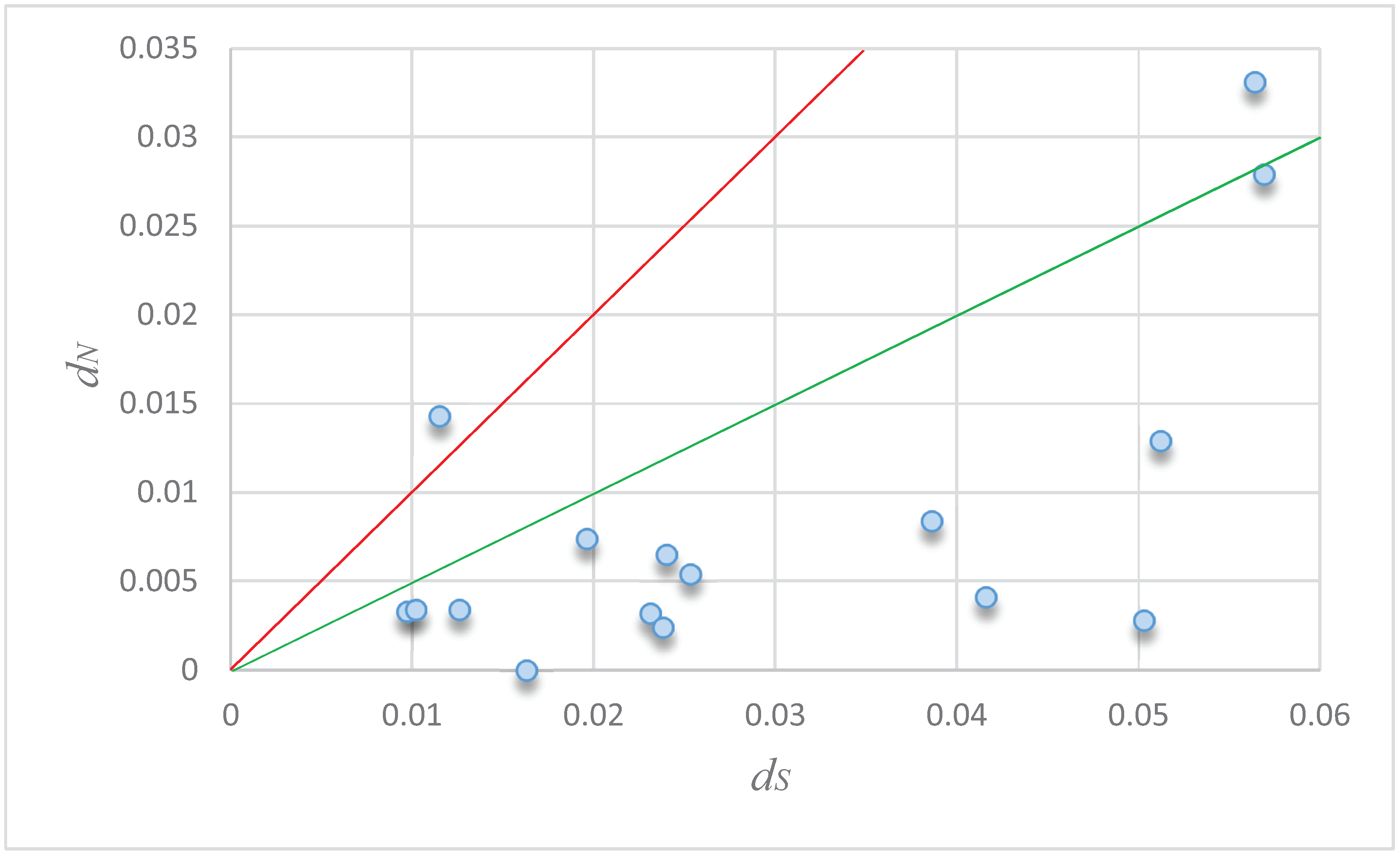
3.4. Molecular Evolution
| Branch model | ω | ln L | χ2 | p |
|---|---|---|---|---|
| One-ratio | ω = 0.14106 for all branches | −8739.54 | ||
| Two-ratios | ω0 = 0.08243 for Clade I | −8719.53 | 40.04 | < 0.001 |
| ω1 = 0.21307 for Clade II | ||||
| Two-ratios | ω0 = 0.08243 for Clade II | |||
| ω1 = 0.21307 for Clade I |

| Tree | Branch model (tree) | ω | ln L | χ2 | p |
|---|---|---|---|---|---|
| Tree 1 | |||||
| One-ratio | ω = 0.09995 for all branches | −4106.505177 | 3.473988 | ||
| Two-ratios | ω0 = 0.12256 for Clade B1 | −4104.768183 | |||
| ω1 = 0.05817 for Clade B2 | |||||
| Tree 2 | |||||
| One-ratio | ω = 0.14031 for all branches | −3879.08919 | 3.149266 | ||
| Two-ratios | ω0 = 0.3196 for Clade A1 | −3877.514557 | |||
| ω1 = 0.1330 for Clade A2 | |||||
| Tree 3 | |||||
| One-ratio | ω = 0.07578 for all branches | −4312.023369 | 0.768848 | ||
| Two-ratios | ω0 = 0.10875 for Clade B1 | −4311.638945 | |||
| ω1 = 0.07299 for Clade B2 | |||||
| Tree 4 | |||||
| One-ratio | ω = 0.19107 for all branches | −3400.606728 | 3.567392 | ||
| Two-ratios | ω0 = 0.34271 for Clade A1 | −3398.823032 | |||
| ω1 = 0.17030 for Clade A2 | |||||
| Tree 5 | |||||
| One-ratio | ω = 0.09507 for all branches | −2426.383206 | 7.862458 | < 0.05 | |
| Two-ratios | ω0 = 0.40447 for Clade A1 | −2422.451977 | |||
| ω1 = 0.08459 for Clade A2 | |||||
| Tree 6 | |||||
| One-ratio | ω = 0.13484 for all branches | −3524.776614 | 5.583034 | < 0.05 | |
| Two-ratios | ω0 = 0.30109 for Clade A1 | −3521.985097 | |||
| ω1 = 0.12124 for Clade A2 |

| Mode | Estimates of parameters | ln L | χ2 | p | Positively selected sites | |
|---|---|---|---|---|---|---|
| Tree6 | Branch model | ω = 0.13484 for all branches | −3524.776614 | 5.583034 | < 0.05 | |
| One-ratio | ω0 = 0.30109 for Clade A1 | −3521.985097 | ||||
| Two-ratios | ω1 = 0.12124 for Clade A2 | |||||
| Branch-site model | ||||||
| Model A0 | ω0 = 0.05407, p0 = 0.82301, ωl = 0.05407, p1 = 0.17699 | −3482.762192 | ||||
| (ω2 = 1) | ω2a fore = 0.40000, ω2a back = 0.05407, P2a = 0.00000 | |||||
| ω2b fore = 1.00000, ω2b back = 0.40000, P2b = 0.00000 | ||||||
| Model A1 | ω0 = 0.05457, p0 = 0.82708, ωl = 1.00000, p1 = 0.15615 | −3471.911359 | 21.701666 | < 0.01 | 425 *, 433 **, 434 * | |
| (ω2 > 1) | ω2a fore = 90.73377, ω2a back = 0.05457, P2a = 0.01411 | |||||
| ω2b fore = 90.73377, ω2b back = 1.00000, P2b = 0.00266 |
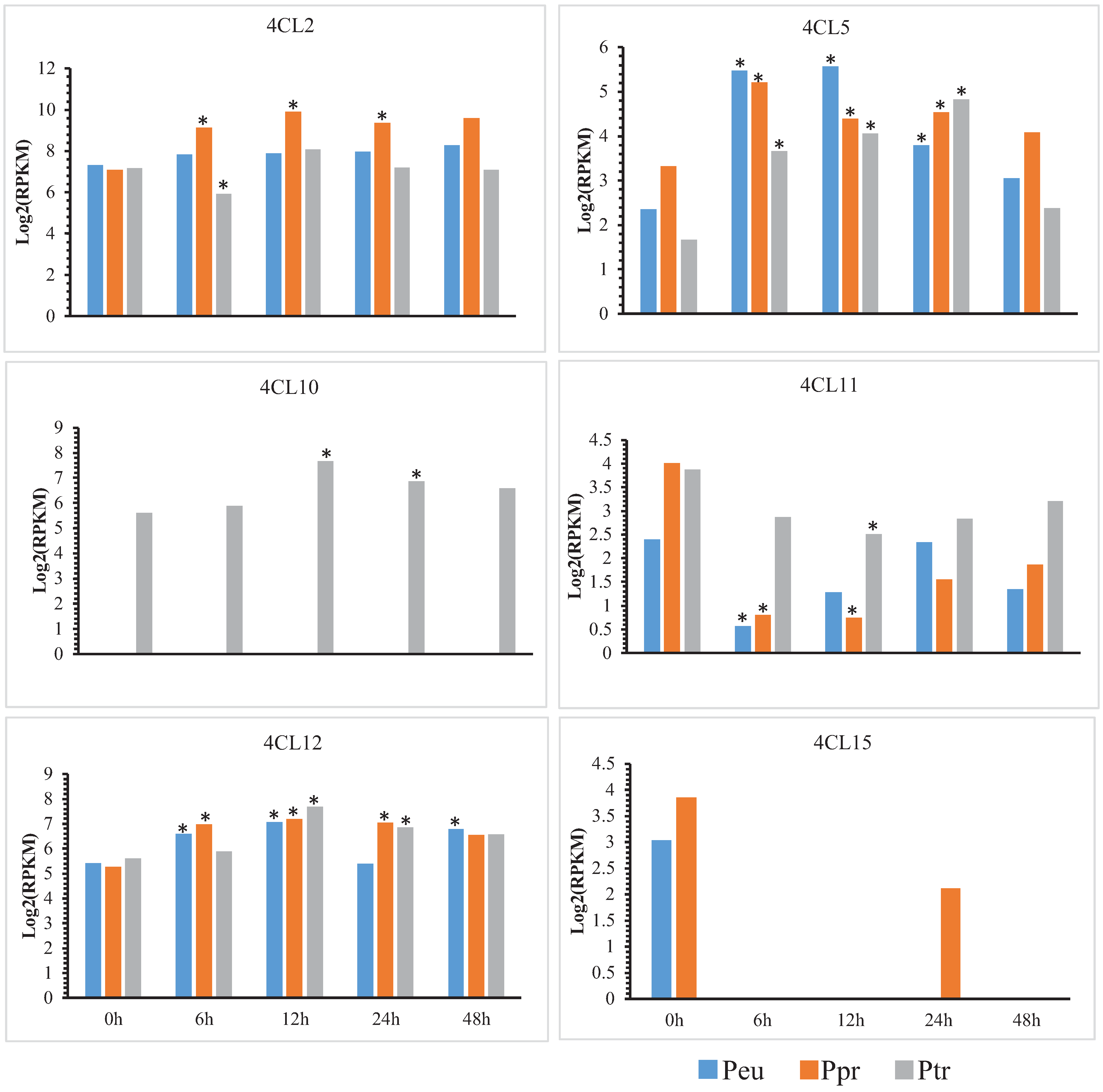
3.5. Changes in Expression of Orthologs Evolved under Salt Stress

4. Conclusions

Supplementary Files
Supplementary File 1Data Deposit
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lee, D.; Meyer, K.; Chapple, C.; Douglas, C.J. Antisense suppression of 4-coumarate:Coenzyme a ligase activity in Arabidopsis leads to altered lignin subunit composition. Plant Cell 1997, 9, 1985–1998. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.J.; Kawaoka, A.; Tsai, C.J.; Lung, J.; Osakabe, K.; Ebinuma, H.; Chiang, V.L. Compartmentalized expression of two structurally and functionally distinct 4-coumarate:CoA ligase genes in aspen (Populus tremuloides). Proc. Natl. Acad. Sci. USA 1998, 95, 5407–5412. [Google Scholar] [CrossRef] [PubMed]
- Hahlbrock, K.; Scheel, D. Physiology and molecular biology of phenylpropanoid metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 347–369. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Yu, S.; Yu, J.; Zhan, Z.; Li, M.; Liu, G.; Wang, X.; Huang, L. Predicting the function of 4-coumarate:CoA ligase (LJ4CL1) in Lonicera japonica. Int. J. Mol. Sci. 2014, 15, 2386–2399. [Google Scholar] [CrossRef] [PubMed]
- Bairoch, A. Prosite: A dictionary of sites and patterns in proteins. Nucleic Acids Res. 1991, 19, 2241–2245. [Google Scholar] [CrossRef] [PubMed]
- Fulda, M.; Heinz, E.; Wolter, F.P. The fadD gene of Escherichia coli K12 is located close to rnd at 39.6 min of the chromosomal-map and is a new member of the AMP-binding protein family. Mol. Gen. Genet. 1994, 242, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Becker-André, M.; Schulze-Lefert, P.; Hahlbrock, K. Structural comparison, modes of expression, and putative cis-acting elements of the two 4-coumarate:CoA ligase genes in potato. J. Biol. Chem. 1991, 266, 8551–8559. [Google Scholar] [PubMed]
- Ragg, H.; Kuhn, D.N.; Hahlbrock, K. Coordinated regulation of 4-coumarate:CoA ligase and phenylalanine ammonia-lyase mRNAs in cultured plant cells. J. Biol. Chem. 1981, 256, 10061–10065. [Google Scholar] [PubMed]
- Zhao, Y.; Kung, S.D.; Dube, S.K. Nucleotide sequence of rice 4-coumarate:CoA ligase gene, 4-CL.1. Nucleic Acids Res. 1990. [Google Scholar] [CrossRef]
- Uhlmann, A.; Ebel, J. Molecular cloning and expression of 4-coumarate:Coenzyme a ligase, an enzyme involved in the resistance response of soybean (Glycine max L.) against pathogen attack. Plant Physiol. 1993, 102, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Voo, K.S.; Whetten, R.W.; O’Malley, D.M.; Sederoff, R.R. 4-Coumarate:Coenzyme a ligase from loblolly pine xylem (isolation, characterization, and complementary DNA cloning). Plant Physiol. 1995, 108, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Ellard, M.; Wanner, L.A.; Davis, K.R.; Douglas, C.J. The Arabidopsis thaliana 4-coumarate:CoA ligase (4CL) gene: Stress and developmentally regulated expression and nucleotide sequence of its cDNA. Plant Mol. Biol. 1995, 28, 871–884. [Google Scholar] [CrossRef] [PubMed]
- Ehlting, J.; Büttner, D.; Wang, Q.; Douglas, C.J.; Somssich, I.E.; Kombrink, E. Three 4-coumarate:Coenzyme a ligases in Arabidopsis thaliana represent two evolutionarily divergent classes in angiosperms. Plant J. 1999, 19, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Kajita, S.; Katayama, Y.; Omori, S. Alterations in the biosynthesis of lignin in transgenic plants with chimeric genes for 4-coumarate:Coenzyme a ligase. Plant Cell Physiol. 1996, 37, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Douglas, C.J. Two divergent members of a tobacco 4-coumarate:Coenzyme a ligase (4CL) gene family (cDNA structure, gene inheritance and expression, and properties of recombinant proteins). Plant Physiol. 1996, 112, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Harding, S.A.; Leshkevich, J.; Chiang, V.L.; Tsai, C.J. Differential substrate inhibition couples kinetically distinct 4-coumarate:Coenzyme a ligases with spatially distinct metabolic roles in quaking aspen. Plant Physiol. 2002, 128, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Allina, S.M.; Pri-Hadash, A.; Theilmann, D.A.; Ellis, B.E.; Douglas, C.J. 4-coumarate:Coenzyme a ligase in hybrid poplar properties of native enzymes, cDNA cloning, and analysis of recombinant enzymes. Plant Physiol. 1998, 116, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Cukovic, D.; Ehlting, J.; VanZiffle, J.A.; Douglas, C.J. Structure and evolution of 4-coumarate:Coenzyme a ligase (4CL) gene families. Biol. Chem. 2001, 382, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Raes, J.; Rohde, A.; Christensen, J.H.; van de Peer, Y.; Boerjan, W. Genome-wide characterization of the lignification toolbox in Arabidopsis. Plant Physiol. 2003, 133, 1051–1071. [Google Scholar] [CrossRef] [PubMed]
- Neves, G.; Marchiosi, R.; Ferrarese, M.; Siqueira-Soares, R.; Ferrarese-Filho, O. Root growth inhibition and lignification induced by salt stress in soybean. J. Agron. Crop Sci. 2010, 196, 467–473. [Google Scholar] [CrossRef]
- Schmelzer, E.; Kruger-Lebus, S.; Hahlbrock, K. Temporal and spatial patterns of gene expression around sites of attempted fungal infection in parsley leaves. Plant Cell 1989, 1, 993–1001. [Google Scholar] [CrossRef] [PubMed]
- Lindermayr, C.; Möllers, B.; Fliegmann, J.; Uhlmann, A.; Lottspeich, F.; Meimberg, H.; Ebel, J. Divergent members of a soybean (Glycine max L.) 4-coumarate:Coenzyme a ligase gene family. Eur. J. Biochem. 2002, 269, 1304–1315. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.A.; Bedgar, D.L.; Moinuddin, S.G.; Kim, K.W.; Cardenas, C.L.; Cochrane, F.C.; Shockey, J.M.; Helms, G.L.; Amakura, Y.; Takahashi, H.; et al. Characterization in vitro and in vivo of the putative multigene 4-coumarate:CoA ligase network in Arabidopsis: Syringyl lignin and sinapate/sinapyl alcohol derivative formation. Phytochemistry 2005, 66, 2072–2091. [Google Scholar] [CrossRef]
- Ottow, E.A.; Brinker, M.; Teichmann, T.; Fritz, E.; Kaiser, W.; Brosché, M.; Kangasjärvi, J.; Jiang, X.; Polle, A. Populus euphratica displays apoplastic sodium accumulation, osmotic adjustment by decreases in calcium and soluble carbohydrates, and develops leaf succulence under salt stress. Plant Physiol. 2005, 139, 1762–1772. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Li, J.; Fritz, E.; Wang, S.; Hüttermann, A. Sodium and chloride distribution in roots and transport in three poplar genotypes under increasing NaCl stress. For. Ecol. Manage. 2002, 168, 217–230. [Google Scholar] [CrossRef]
- Browicz, K. Chorology of Populus euphratica olivier. Arbor. Kornickie 1977, 22, 5–27. [Google Scholar]
- Thevs, N.; Buras, A.; Zerbe, S.; Kühnel, E.; Abdusalih, N.; Ovezberdiyeva, A. Structure and wood biomass of near-natural floodplain forests along the central asian rivers tarim and amu darya. Forestry 2011. [Google Scholar] [CrossRef]
- Ma, T.; Wang, J.; Zhou, G.; Yue, Z.; Hu, Q.; Chen, Y.; Liu, B.; Qiu, Q.; Wang, Z.; Zhang, J.; et al. Genomic insights into salt adaptation in a desert poplar. Nat. Commun. 2013. [Google Scholar] [CrossRef] [PubMed]
- Tuskan, G.A.; Difazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar] [PubMed]
- Nordberg, H.; Cantor, M.; Dusheyko, S.; Hua, S.; Poliakov, A.; Shabalov, I.; Smirnova, T.; Grigoriev, I.V.; Dubchak, I. The genome portal of the Department of Energy Joint Genome Institute: 2014 updates. Nucleic Acids Res. 2014, 42, D26–D31. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Liu, J. Genome of Populus pruinosa. Unpublished data. 2015. [Google Scholar]
- Dai, X.; Hu, Q.; Cai, Q.; Feng, K.; Ye, N.; Tuskan, G.A.; Milne, R.; Chen, Y.; Wan, Z.; Wang, Z.; et al. The willow genome and divergent evolution from poplar after the common genome duplication. Cell Res. 2014, 24, 1274–1277. [Google Scholar] [CrossRef] [PubMed]
- Cell wall genomics. Available online: http://cellwall.genomics.purdue.edu/families/index.html (accessed on 10 September 2015).
- Lamesch, P.; Berardini, T.Z.; Li, D.; Swarbreck, D.; Wilks, C.; Sasidharan, R.; Muller, R.; Dreher, K.; Alexander, D.L.; Garcia-Hernandez, M. The Arabidopsis information resource (tair): Improved gene annotation and new tools. Nucleic Acids Res. 2012, 40, D1202–D1210. [Google Scholar] [CrossRef] [PubMed]
- Eddy, S.R. Profile hidden markov models. Bioinformatics 1998, 14, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Edgar, R.C. Muscle: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Mega6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Ferraiolo, J.; Jun, F.; Jackson, D. Scalable Vector Graphics (SVG) 1.0 Specification; Iuniverse: Bloomington, IN, USA, 2000. [Google Scholar]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.; Milpetz, F.; Bork, P.; Ponting, C.P. Smart, a simple modular architecture research tool: Identification of signaling domains. Proc. Natl. Acad. Sci. USA 1998, 95, 5857–5864. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Doerks, T.; Bork, P. Smart 7: Recent updates to the protein domain annotation resource. Nucleic Acids Res. 2012, 40, D302–D305. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.P.; Remm, M.; Sonnhammer, E.L. Inparanoid: A comprehensive database of eukaryotic orthologs. Nucleic Acids Res. 2005, 33, D476–D480. [Google Scholar] [CrossRef] [PubMed]
- Alexeyenko, A.; Tamas, I.; Liu, G.; Sonnhammer, E.L. Automatic clustering of orthologs and inparalogs shared by multiple proteomes. Bioinformatics 2006, 22, e9–e15. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wong, W.S.; Nielsen, R. Bayes empirical bayes inference of amino acid sites under positive selection. Mol. Biol. Evol. 2005, 22, 1107–1118. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Nielsen, R.; Goldman, N.; Pedersen, A.M. Codon-substitution models for heterogeneous selection pressure at amino acid sites. Genetics 2000, 155, 431–449. [Google Scholar] [PubMed]
- Luo, W.; Wan, D.; Liu, J. Transcriptome changes in four poplars in response to salt stress. Unpublished data. 2015. [Google Scholar]
- Zhang, J.; Jiang, D.; Liu, B.; Luo, W.; Lu, J.; Ma, T.; Wan, D. Transcriptome dynamics of a desert poplar (Populus pruinosa) in response to continuous salinity stress. Plant Cell Rep. 2014, 33, 1565–1579. [Google Scholar] [CrossRef] [PubMed]
- Gu, R.S.; Jiang, X.N.; Guo, Z.C. Organogenesis and plantlet regeneration in vitro of Populus euphratica (In Chinese). Acta Bot. Sin. 1999, 41, 29–33. [Google Scholar]
- Zhang, F.; Yang, Y.; He, W.; Zhao, X.; Zhang, L. Effects of salinity on growth and compatible solutes of callus induced from Populus euphratica. In Vitro Cell. Dev. Biol. Plant 2004, 40, 491–494. [Google Scholar] [CrossRef]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Rep. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. Edger: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Feng, J.; Lu, J.; Yang, Y.; Zhang, X.; Wan, D.; Liu, J. Transcriptome differences between two sister desert poplar species under salt stress. BMC Genomics 2014. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhang, D.; Hu, J.; Zhou, X.; Ye, X.; Reichel, K.L.; Stewart, N.R.; Syrenne, R.D.; Yang, X.; Gao, P.; et al. Comparative genome analysis of lignin biosynthesis gene families across the plant kingdom. BMC Bioinformatics 2009. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J. Positive selection, not negative selection, in the pseudogenization of rcsa in Yersinia pestis. Proc. Natl. Acad. Sci. USA 2008. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.-H.; Ma, T.; Luo, W.-C.; Xu, J.-M.; Liu, J.-Q.; Wan, D.-S. Identification of 4CL Genes in Desert Poplars and Their Changes in Expression in Response to Salt Stress. Genes 2015, 6, 901-917. https://doi.org/10.3390/genes6030901
Zhang C-H, Ma T, Luo W-C, Xu J-M, Liu J-Q, Wan D-S. Identification of 4CL Genes in Desert Poplars and Their Changes in Expression in Response to Salt Stress. Genes. 2015; 6(3):901-917. https://doi.org/10.3390/genes6030901
Chicago/Turabian StyleZhang, Cai-Hua, Tao Ma, Wen-Chun Luo, Jian-Mei Xu, Jian-Quan Liu, and Dong-Shi Wan. 2015. "Identification of 4CL Genes in Desert Poplars and Their Changes in Expression in Response to Salt Stress" Genes 6, no. 3: 901-917. https://doi.org/10.3390/genes6030901
APA StyleZhang, C. -H., Ma, T., Luo, W. -C., Xu, J. -M., Liu, J. -Q., & Wan, D. -S. (2015). Identification of 4CL Genes in Desert Poplars and Their Changes in Expression in Response to Salt Stress. Genes, 6(3), 901-917. https://doi.org/10.3390/genes6030901