Cupriavidus metallidurans Strains with Different Mobilomes and from Distinct Environments Have Comparable Phenomes

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media and Culture Conditions
2.2. Growth in the Presence of Metals
2.3. NA4 CRISPR Deletion Construction
2.4. Construction of Plasmids
2.5. Conjugation Assay for Testing CRISPR-Cas
2.6. Plasmid Profiling
2.7. Phenotype Microarray Analysis
2.8. Computational Methods
3. Results and Discussion
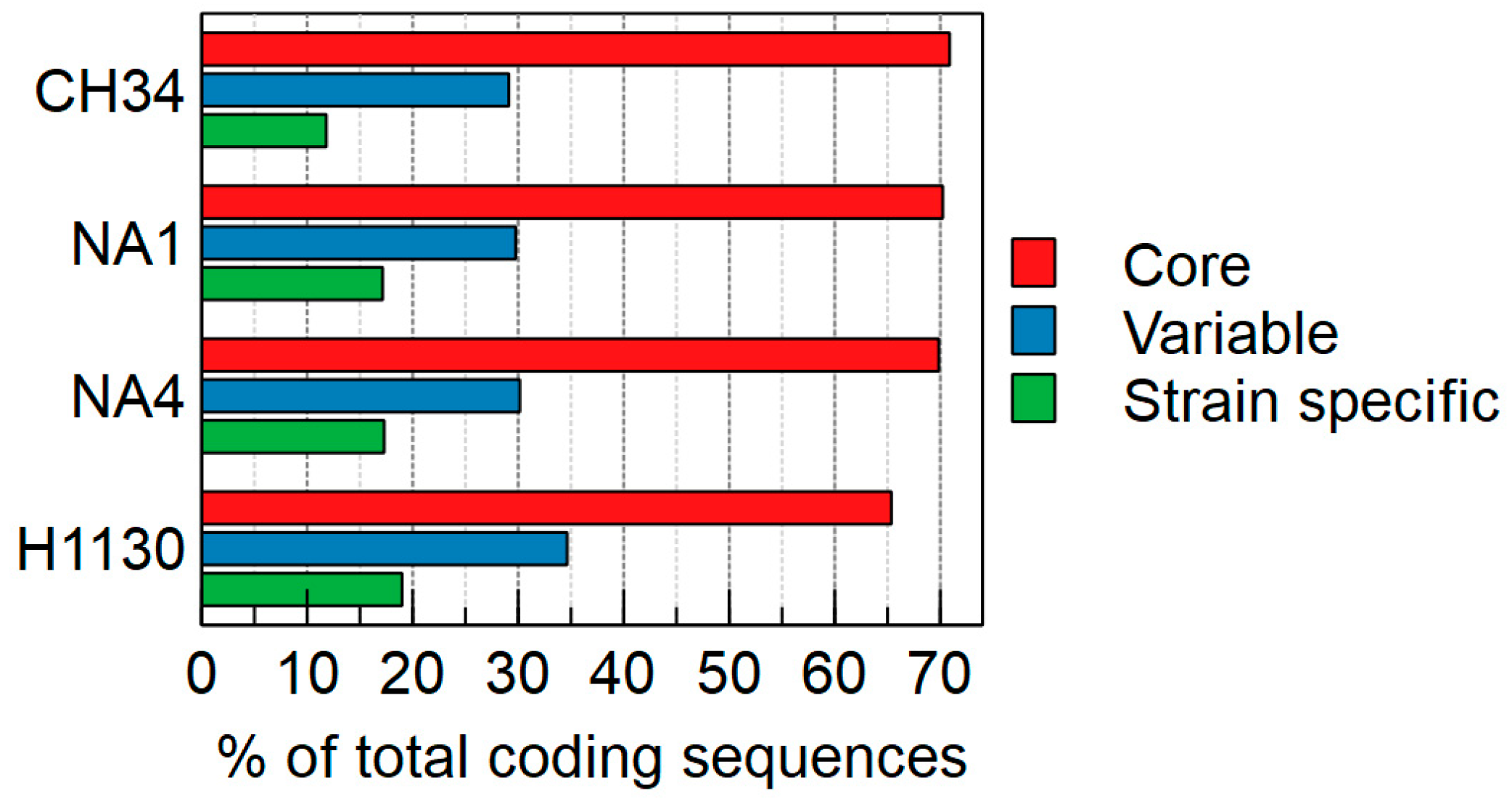
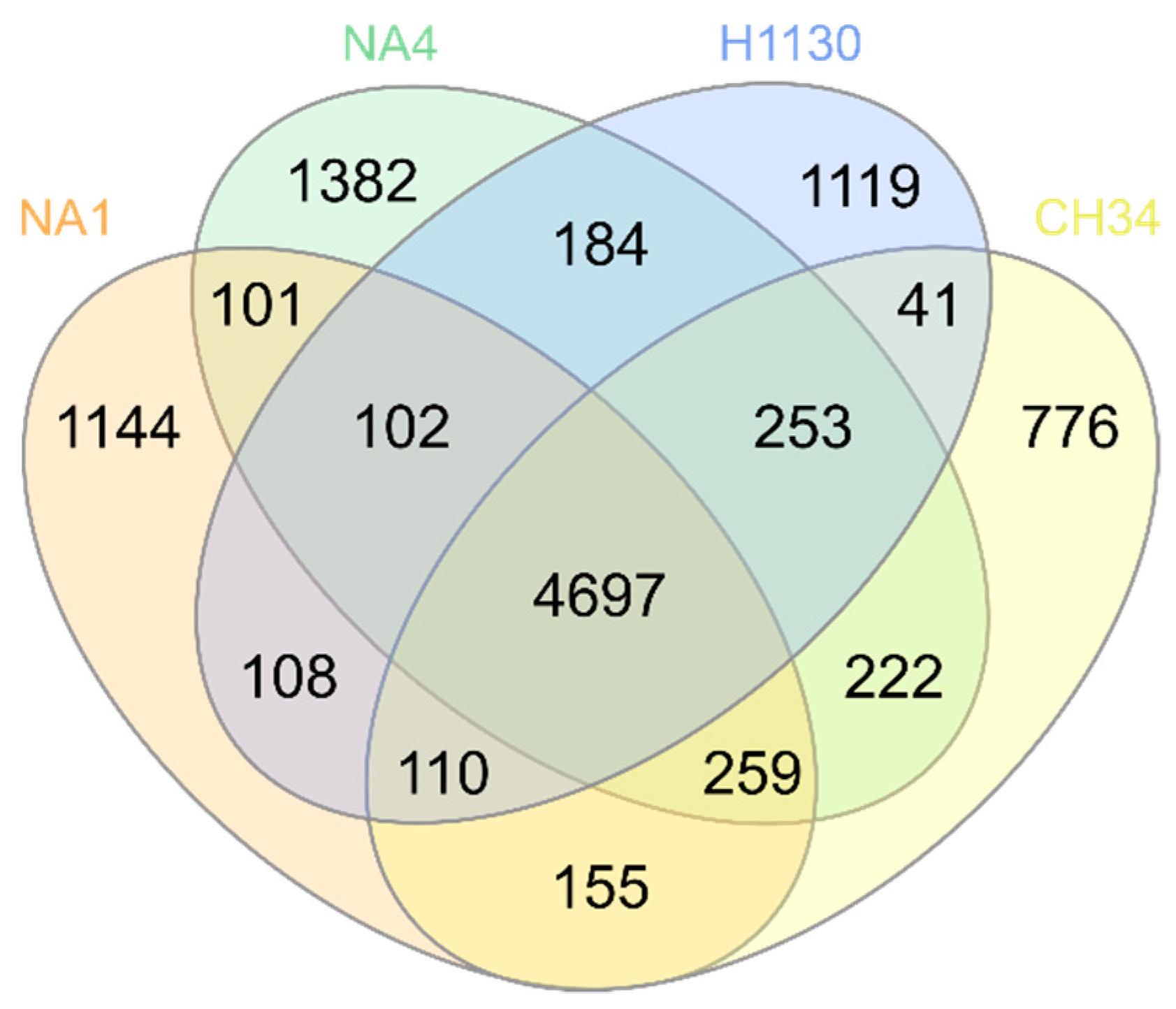
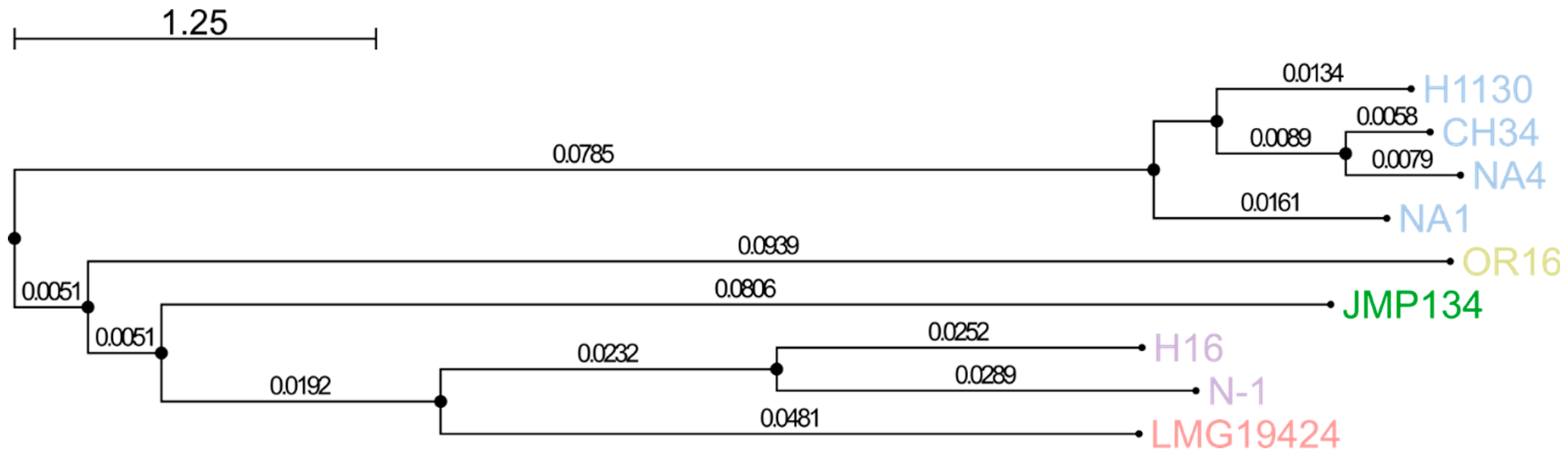
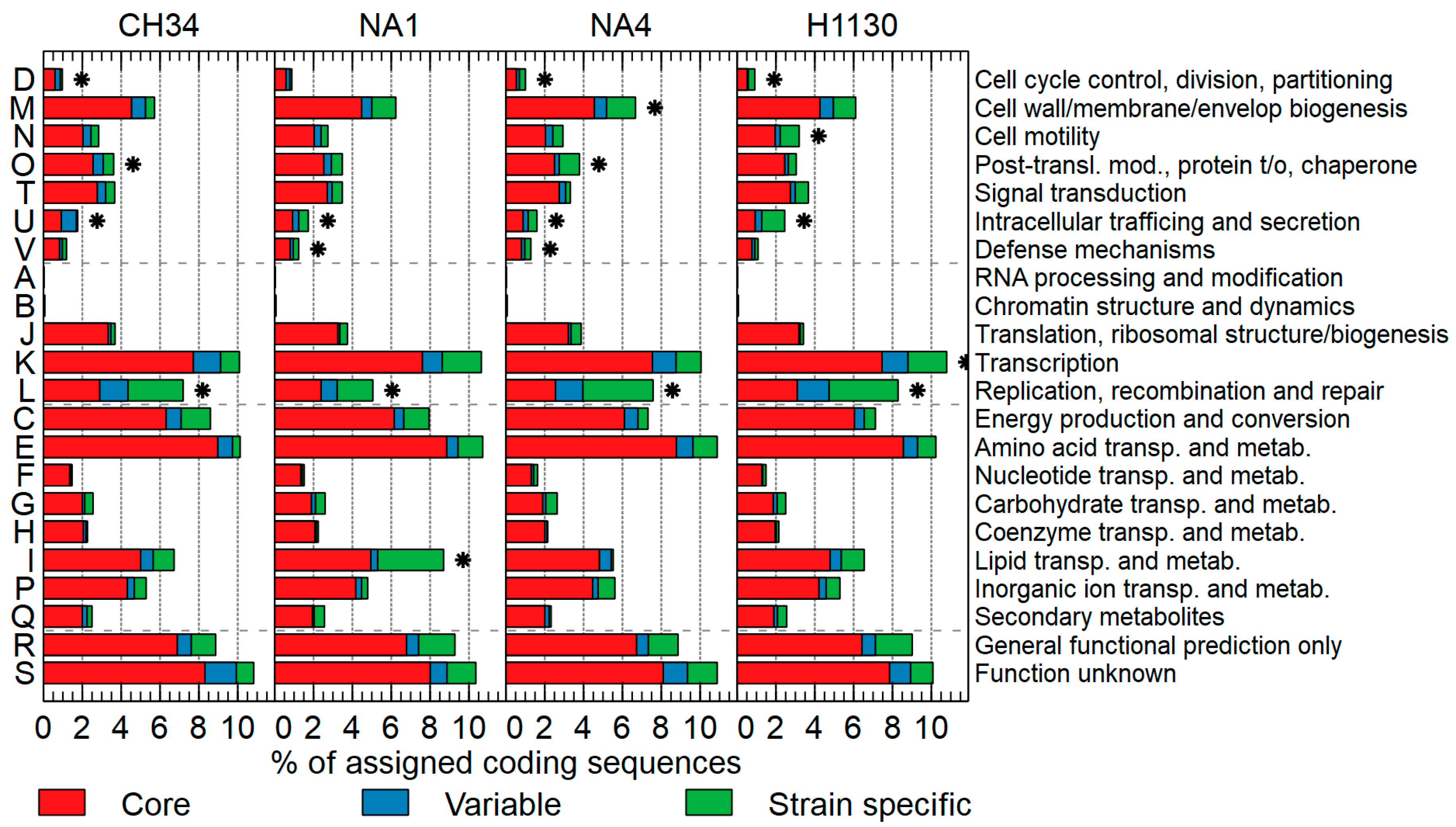
3.1. Comparison of General Genome Features
3.2. The Mobilome
3.2.1. Insertion Sequence Elements and Transposons
3.2.2. Genomic Islands
3.2.3. Prophages
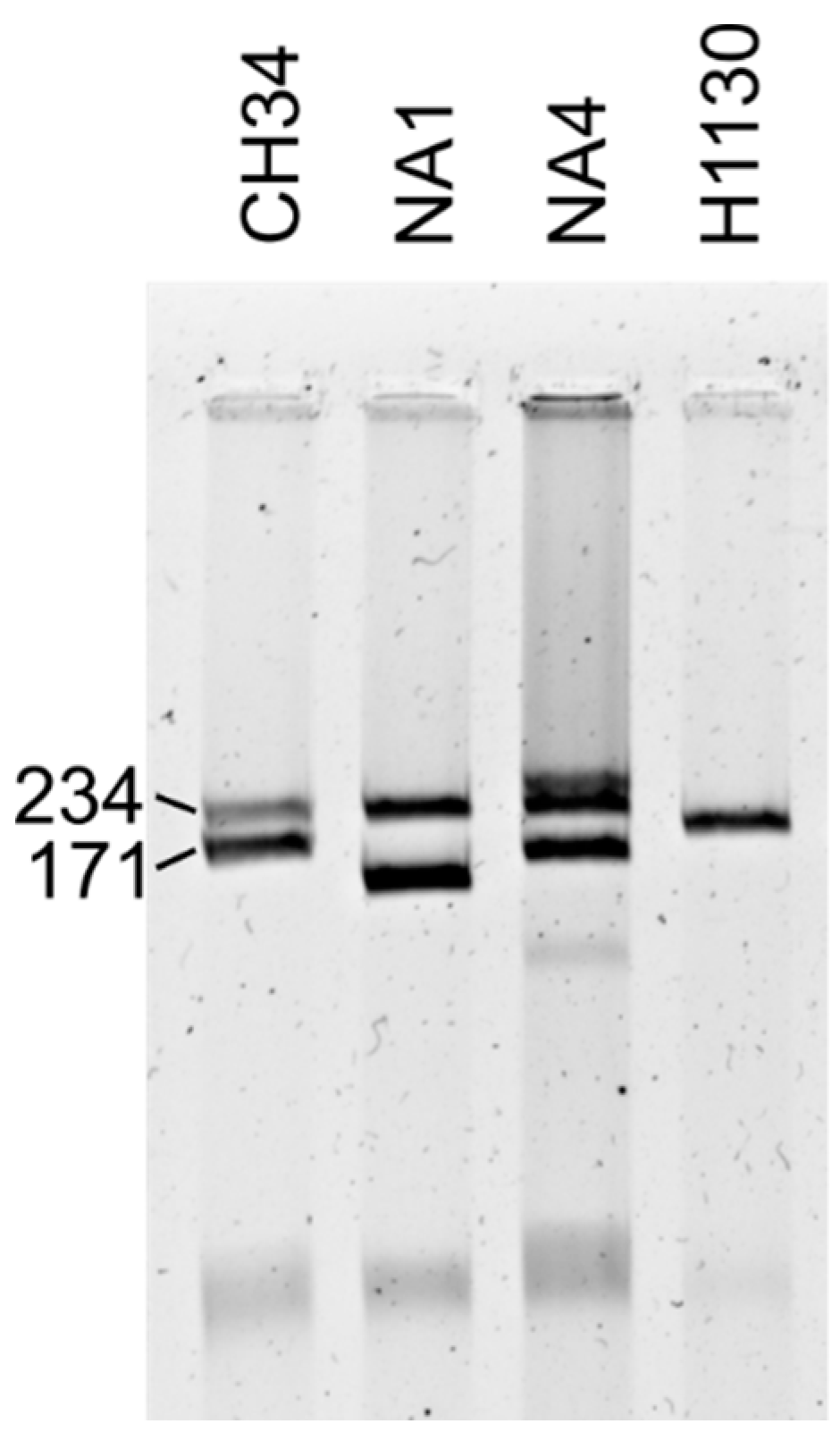
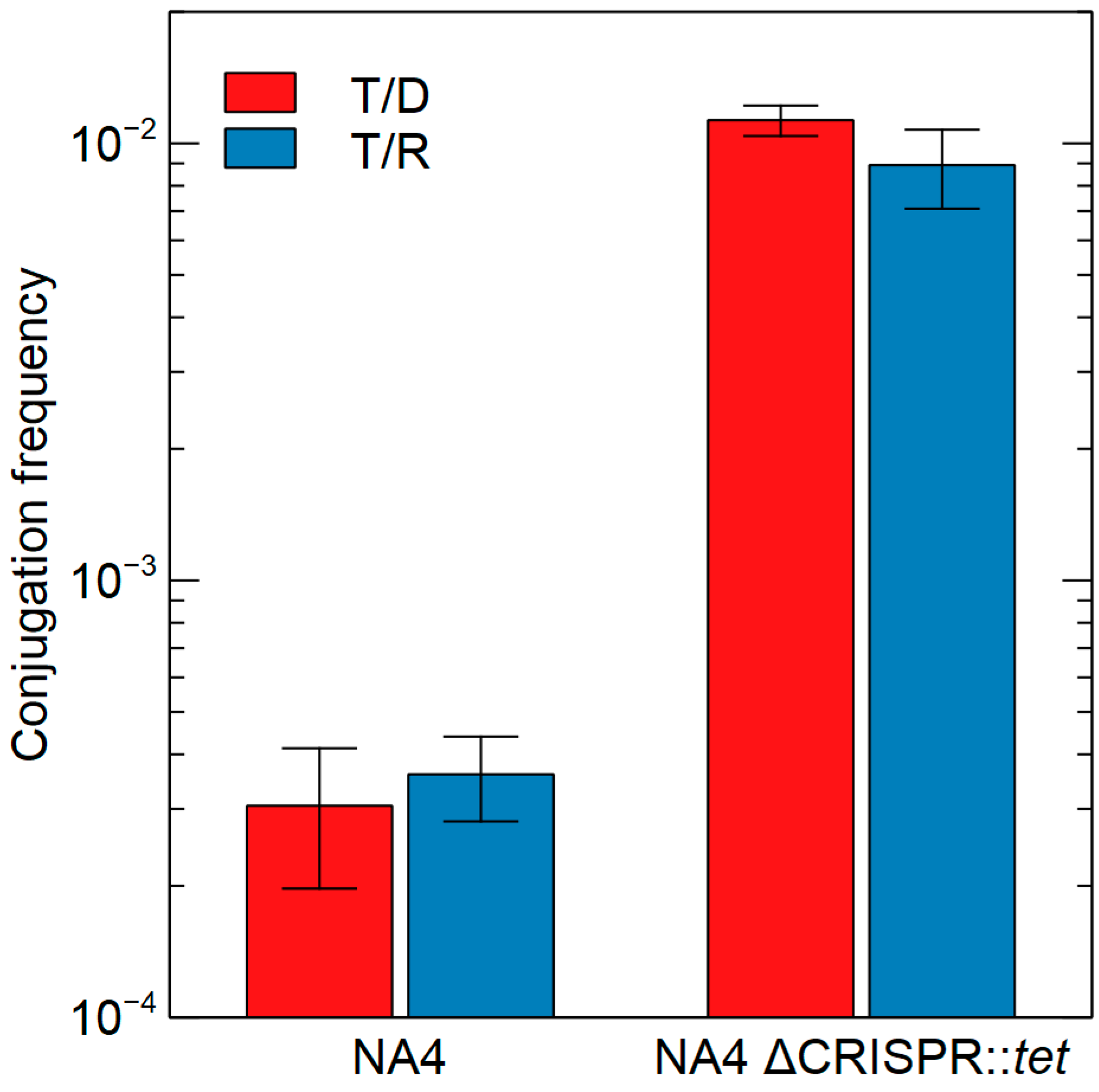
3.2.4. CRISPR-Cas
3.3. The Resistome
3.3.1. Antibiotic Resistance
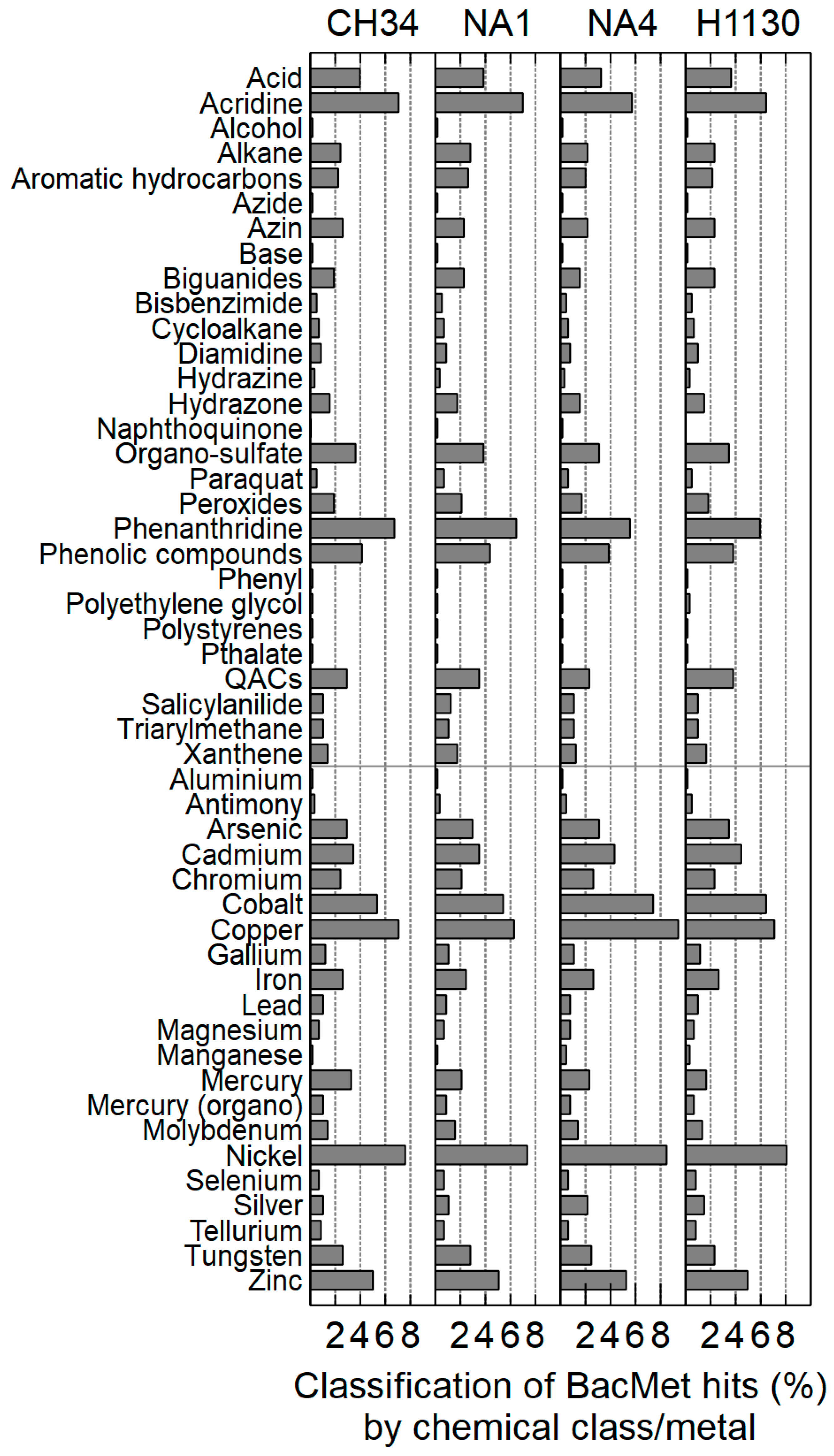
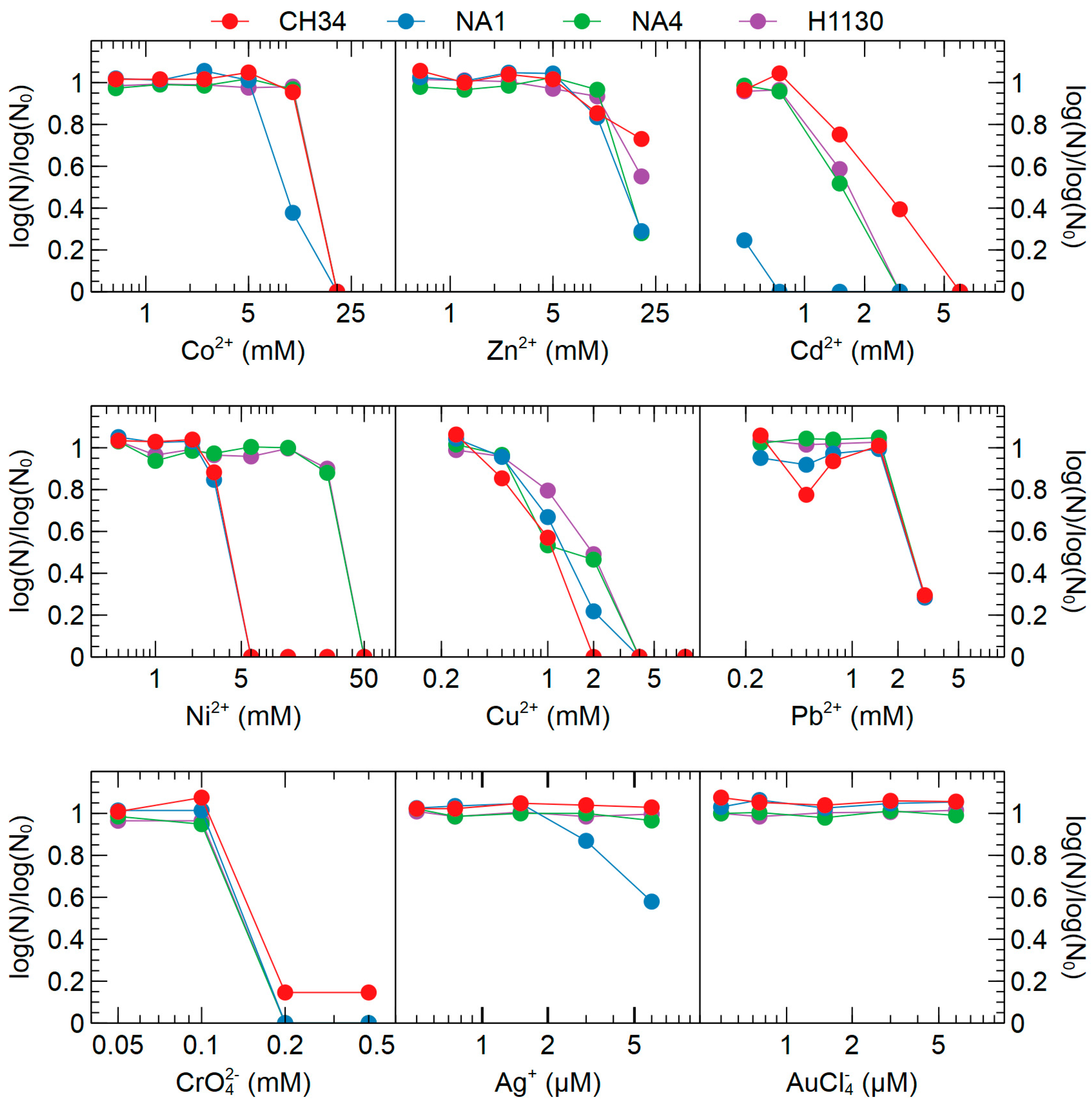
3.3.2. Metal Resistance
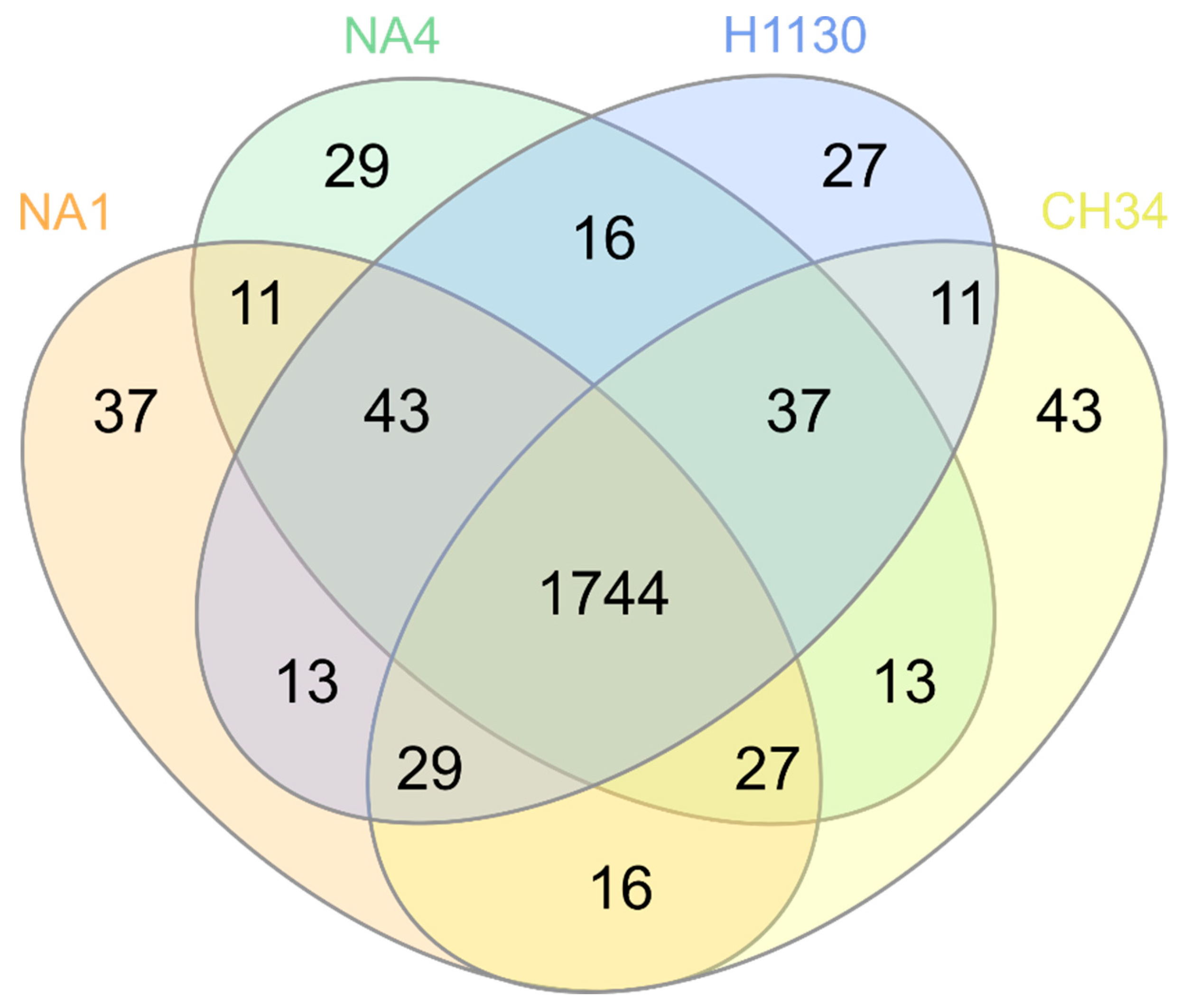
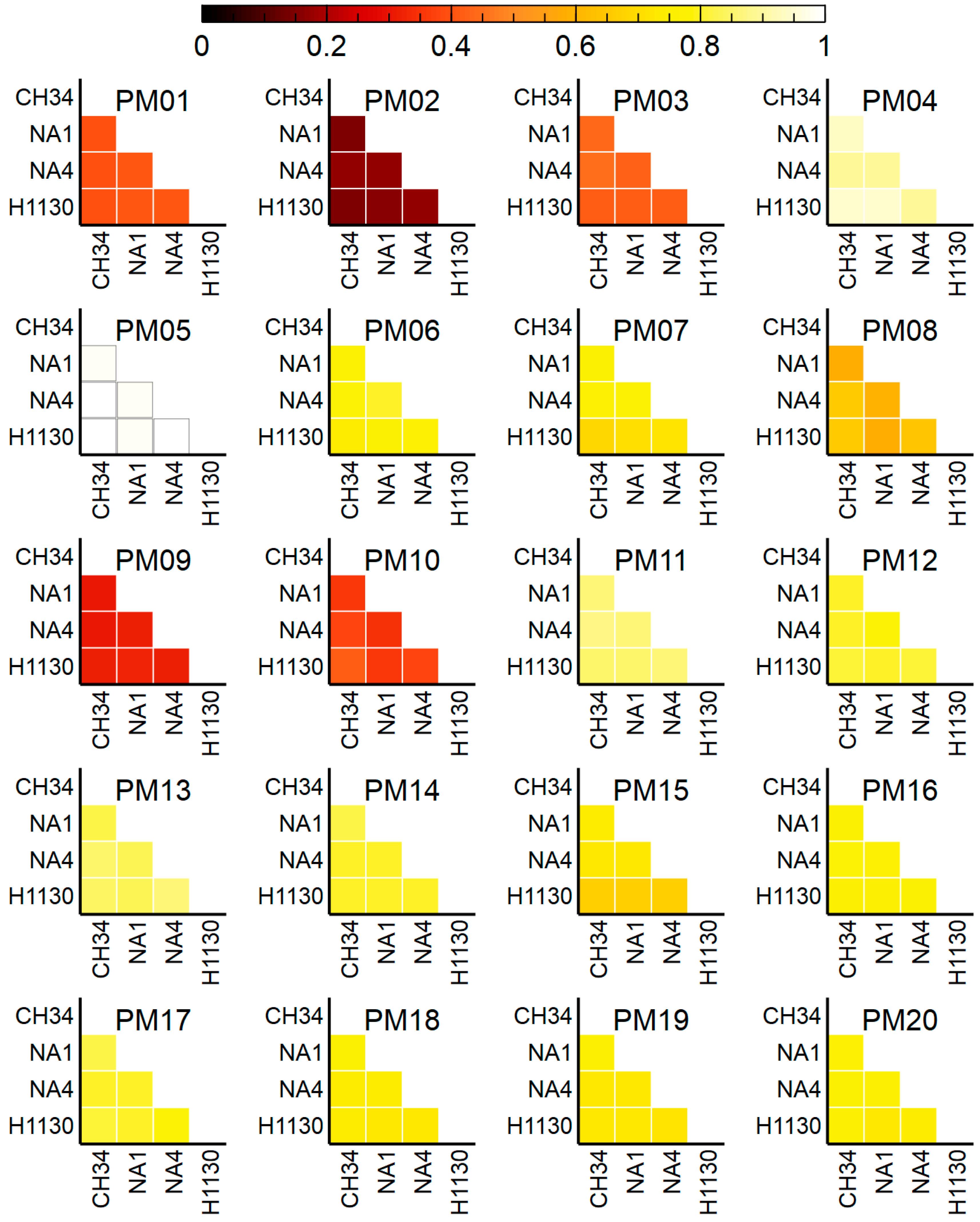
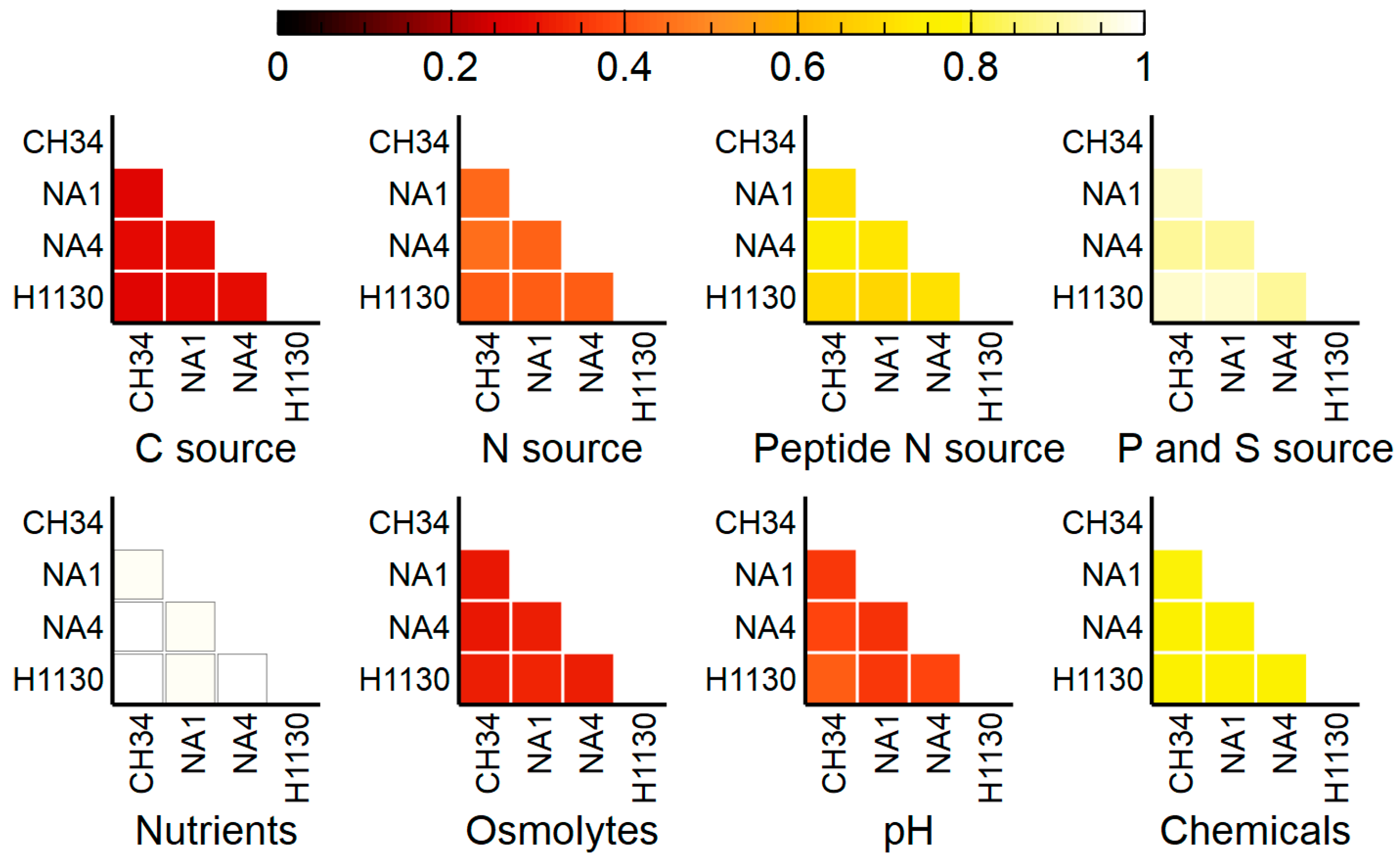
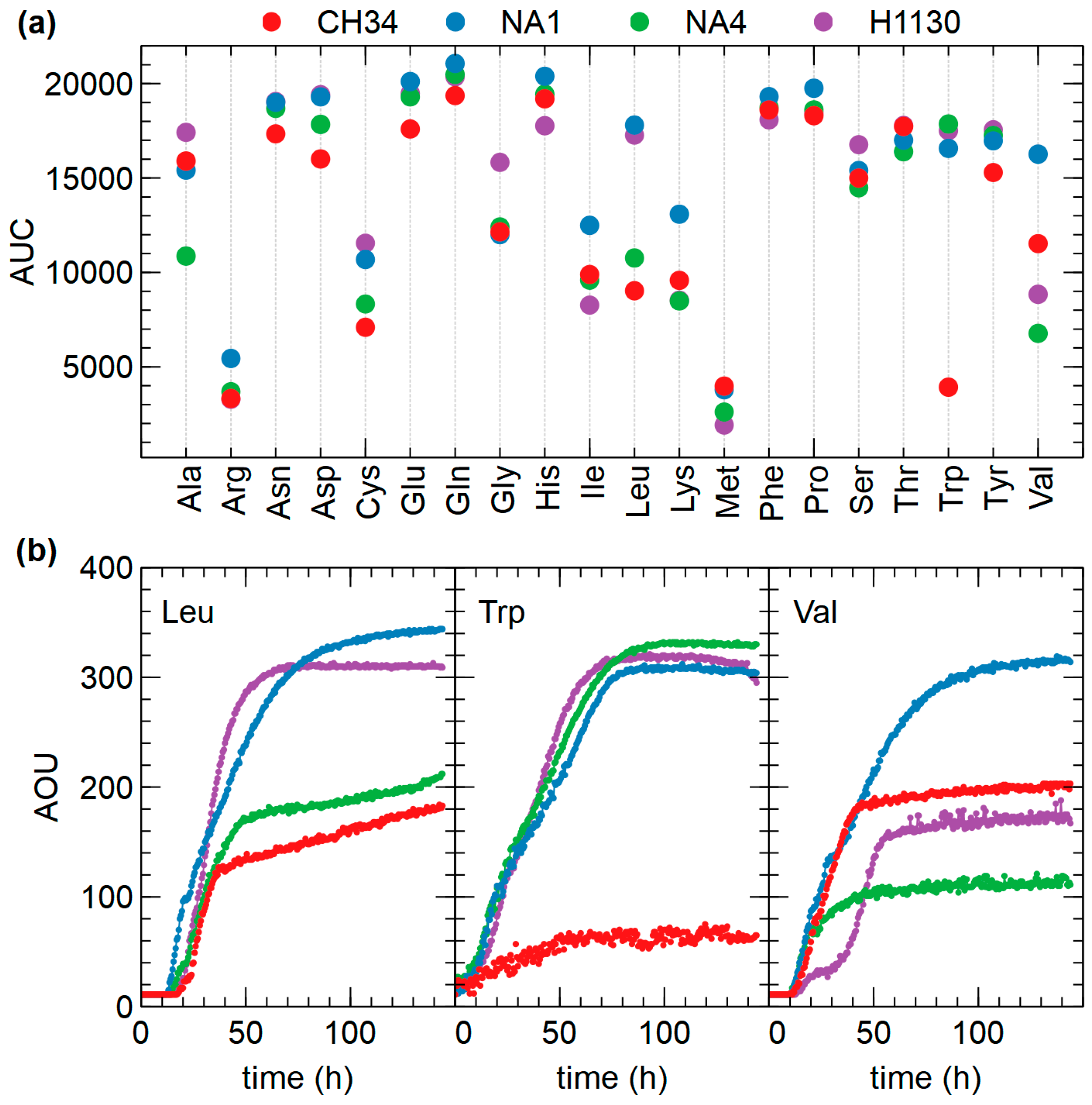
3.4. Phenotypic Microarrays
3.4.1. C, N, P and S Sources
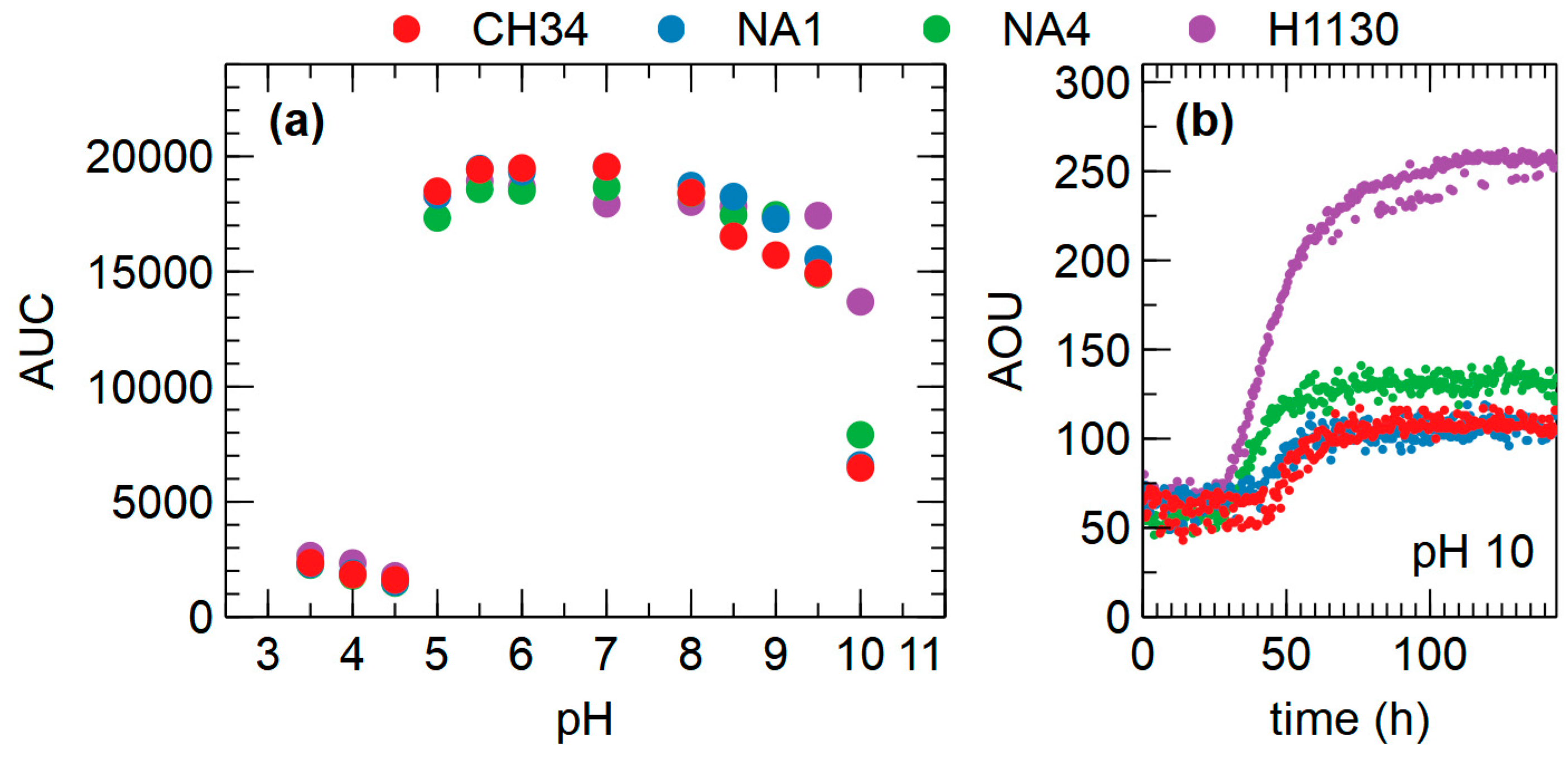
3.4.2. Osmolytes and pH
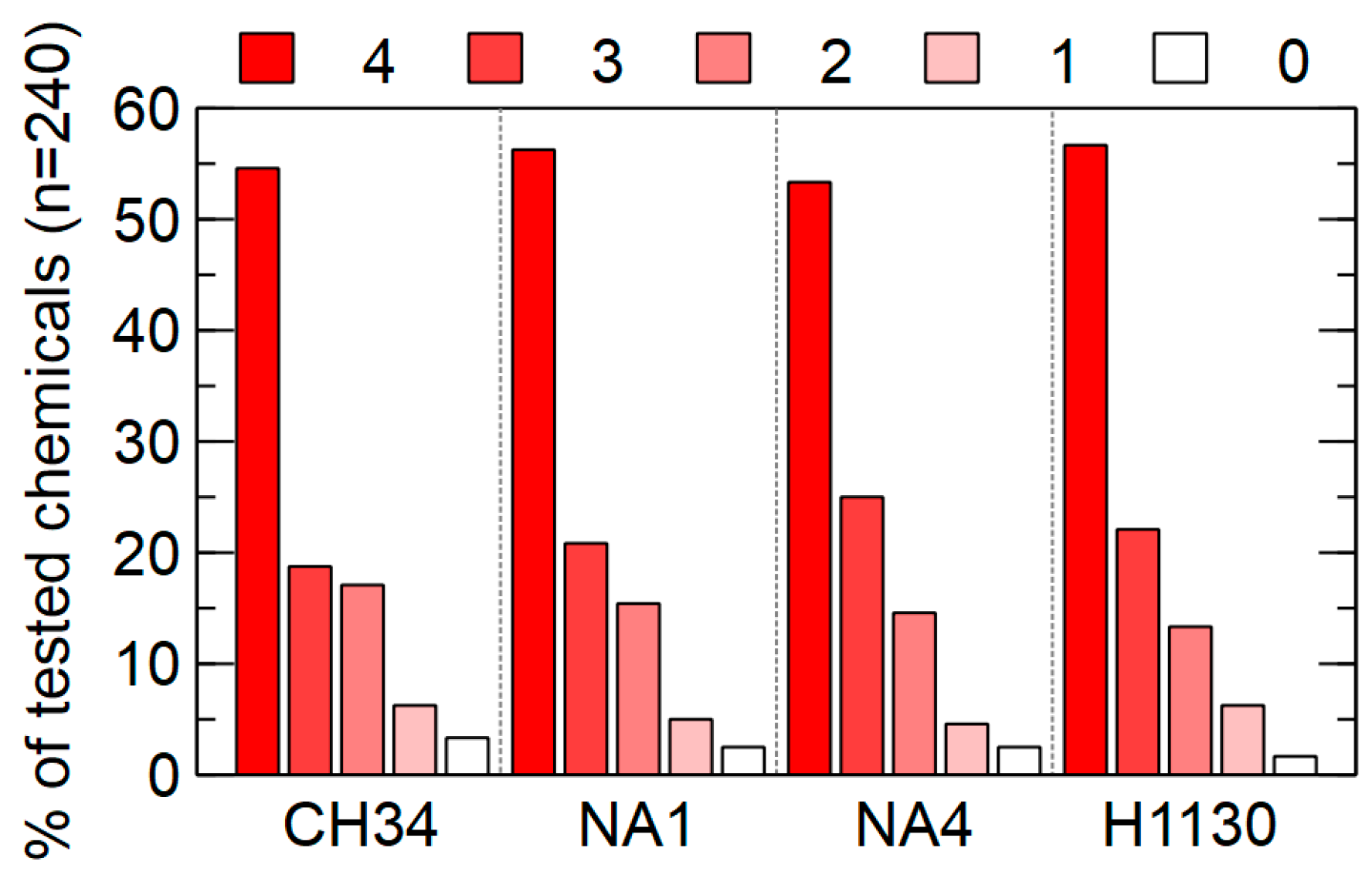
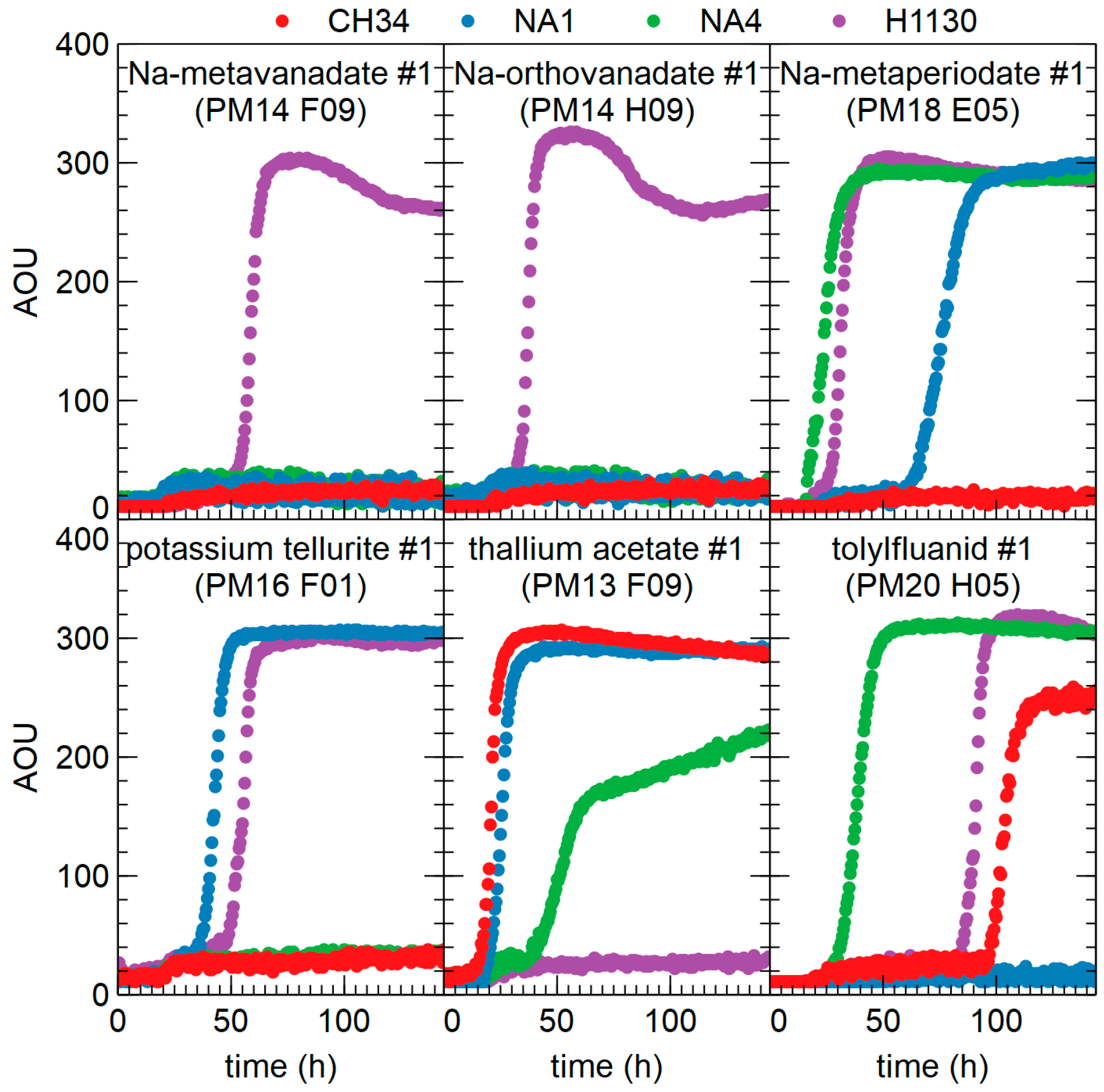
3.4.3. Chemicals
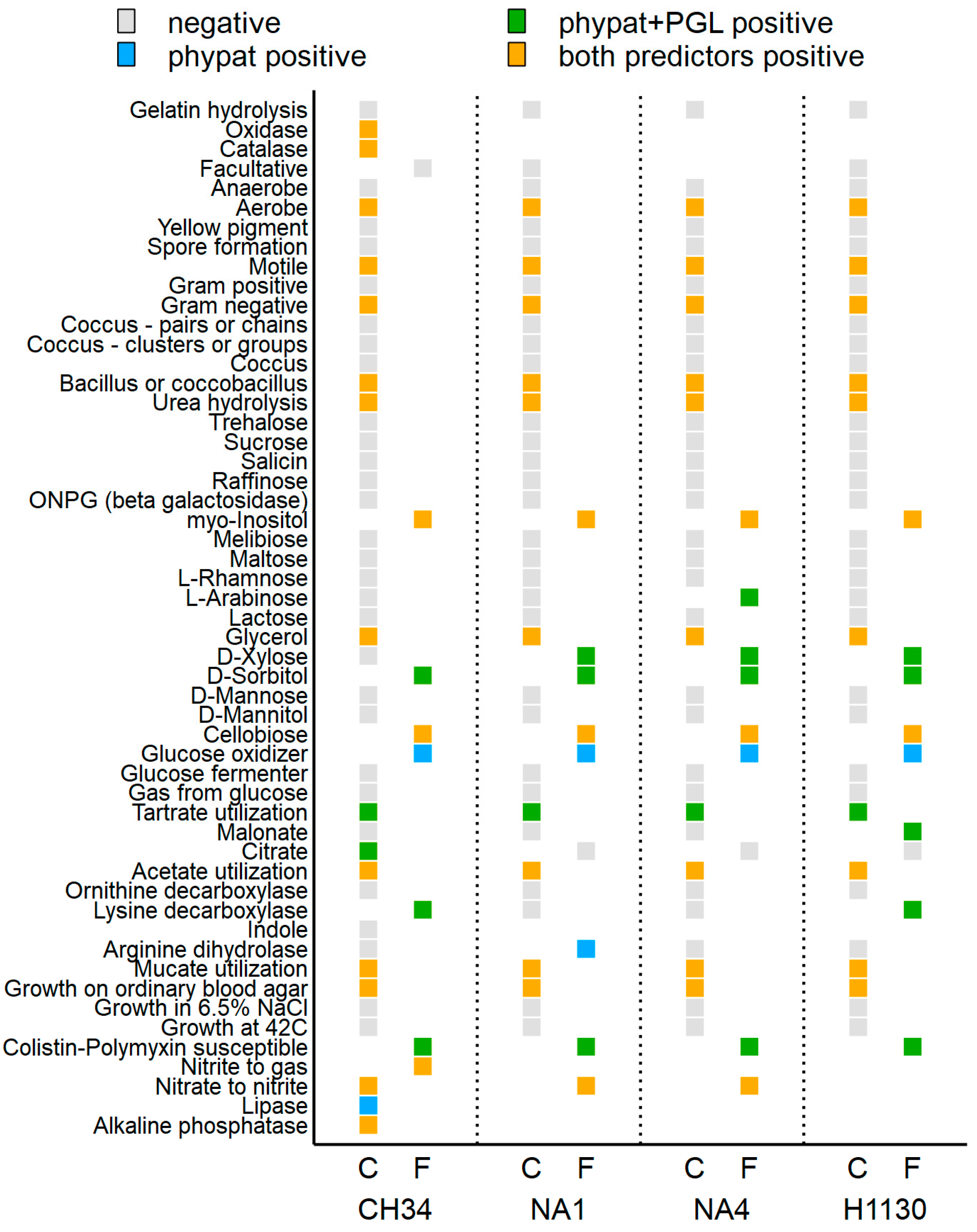
3.4.4. Trait Prediction
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mergeay, M.; Nies, D.; Schlegel, H.G.; Gerits, J.; Charles, P.; Van Gijsegem, F. Alcaligenes eutrophus CH34 is a facultative chemolithotroph with plasmid-bound resistance to heavy metals. J. Bacteriol. 1985, 162, 328–334. [Google Scholar] [PubMed]
- Mergeay, M.; Van Houdt, R. Metal Response in Cupriavidus Metallidurans, Volume I: From Habitats to Genes and Proteins; Springer International Publishing: Cham, Switzerland, 2015; p. 89. [Google Scholar]
- Janssen, P.J.; Van Houdt, R.; Moors, H.; Monsieurs, P.; Morin, N.; Michaux, A.; Benotmane, M.A.; Leys, N.; Vallaeys, T.; Lapidus, A.; et al. The complete genome sequence of Cupriavidus metallidurans strain CH34, a master survivalist in harsh and anthropogenic environments. PLoS ONE 2010, 5, e10433. [Google Scholar] [CrossRef] [PubMed]
- Monchy, S.; Benotmane, M.A.; Janssen, P.; Vallaeys, T.; Taghavi, S.; van der Lelie, D.; Mergeay, M. Plasmids pMOL28 and pMOL30 of Cupriavidus metallidurans are specialized in the maximal viable response to heavy metals. J. Bacteriol. 2007, 189, 7417–7425. [Google Scholar] [CrossRef] [PubMed]
- Vandenbussche, G.; Mergeay, M.; Van Houdt, R. Metal Response in Cupriavidus metallidurans, Volume II: Insights into the Structure-Function Relationship of Proteins; Springer International Publishing: Cham, Switzerland, 2015; p. 70. [Google Scholar]
- Diels, L.; Mergeay, M. DNA probe-mediated detection of resistant bacteria from soils highly polluted by heavy metals. Appl. Environ. Microbiol. 1990, 56, 1485–1491. [Google Scholar] [PubMed]
- Van Houdt, R.; Monsieurs, P.; Mijnendonckx, K.; Provoost, A.; Janssen, A.; Mergeay, M.; Leys, N. Variation in genomic islands contribute to genome plasticity in Cupriavidus metallidurans. BMC Genom. 2012, 13, 111. [Google Scholar] [CrossRef] [PubMed]
- Kunito, T.; Kusano, T.; Oyaizu, H.; Senoo, K.; Kanazawa, S.; Matsumoto, S. Cloning and sequence analysis of czc genes in Alcaligenes sp. strain CT14. Biosci. Biotechnol. Biochem. 1996, 60, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Reith, F.; Rogers, S.L.; McPhail, D.C.; Webb, D. Biomineralization of gold: Biofilms on bacterioform gold. Science 2006, 313, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.; Schlegel, H.G. Nickel and cobalt resistance of various bacteria isolated from soil and highly polluted domestic and industrial wastes. FEMS Microbiol. Lett. 1989, 62, 315–328. [Google Scholar] [CrossRef]
- Miyake-Nakayama, C.; Masujima, S.; Ikatsu, H.; Miyoshi, S.-I.; Shinoda, S. Isolation and characterization of a new dichloromethane degrading bacterium, Ralstonia metallidurans, PD11. Biocontrol Sci. 2004, 9, 89–93. [Google Scholar] [CrossRef]
- Mijnendonckx, K.; Provoost, A.; Ott, C.M.; Venkateswaran, K.; Mahillon, J.; Leys, N.; Van Houdt, R. Characterization of the survival ability of Cupriavidus metallidurans and Ralstonia pickettii from space-related environments. Microbol. Ecol. 2013, 65, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Mora, M.; Perras, A.; Alekhova, T.A.; Wink, L.; Krause, R.; Aleksandrova, A.; Novozhilova, T.; Moissl-Eichinger, C. Resilient microorganisms in dust samples of the International Space Station-survival of the adaptation specialists. Microbiome 2016, 4, 65. [Google Scholar] [CrossRef] [PubMed]
- Coenye, T.; Spilker, T.; Reik, R.; Vandamme, P.; Lipuma, J.J. Use of PCR analyses to define the distribution of Ralstonia species recovered from patients with cystic fibrosis. J. Clin. Microbiol. 2005, 43, 3463–3466. [Google Scholar] [CrossRef] [PubMed]
- Langevin, S.; Vincelette, J.; Bekal, S.; Gaudreau, C. First case of invasive human infection caused by Cupriavidus metallidurans. J. Clin. Microbiol. 2011, 49, 744–745. [Google Scholar] [CrossRef] [PubMed]
- D’Inzeo, T.; Santangelo, R.; Fiori, B.; De Angelis, G.; Conte, V.; Giaquinto, A.; Palucci, I.; Scoppettuolo, G.; Di Florio, V.; Giani, T.; et al. Catheter-related bacteremia by Cupriavidus metallidurans. Diagn. Microbiol. Infect. Dis. 2015, 81, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.W.; Lower, R.P.; Kim, N.K.; Young, J.P. Introducing the bacterial ‘chromid’: Not a chromosome, not a plasmid. Trends Microbiol. 2010, 18, 141–148. [Google Scholar] [CrossRef] [PubMed]
- DiCenzo, G.C.; Finan, T.M. The divided bacterial genome: Structure, function, and evolution. Microbiol. Mol. Biol. Rev. 2017, 81, e00019-17. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, E. Microbial Megaplasmids; Springer: Berlin/Heidelberg, Germany, 2009; p. 348. [Google Scholar]
- Schwartz, E.; Henne, A.; Cramm, R.; Eitinger, T.; Friedrich, B.; Gottschalk, G. Complete nucleotide sequence of pHG1: A Ralstonia eutropha H16 megaplasmid encoding key enzymes of H2-based lithoautotrophy and anaerobiosis. J. Mol. Biol. 2003, 332, 369–383. [Google Scholar] [CrossRef]
- Amadou, C.; Pascal, G.; Mangenot, S.; Glew, M.; Bontemps, C.; Capela, D.; Carrere, S.; Cruveiller, S.; Dossat, C.; Lajus, A.; et al. Genome sequence of the β-rhizobium Cupriavidus taiwanensis and comparative genomics of rhizobia. Genom. Res. 2008, 18, 1472–1483. [Google Scholar] [CrossRef] [PubMed]
- Trefault, N.; De la Iglesia, R.; Molina, A.M.; Manzano, M.; Ledger, T.; Perez-Pantoja, D.; Sanchez, M.A.; Stuardo, M.; Gonzalez, B. Genetic organization of the catabolic plasmid pJP4 from Ralstonia eutropha JMP134 (pJP4) reveals mechanisms of adaptation to chloroaromatic pollutants and evolution of specialized chloroaromatic degradation pathways. Environ. Microbiol. 2004, 6, 655–668. [Google Scholar] [CrossRef] [PubMed]
- Van Houdt, R.; Mergeay, M. Genomic context of metal response genes in Cupriavidus metallidurans with a focus on strain CH34. In Metal Response in Cupriavidus Metallidurans, Volume I: From Habitats to Genes and Proteins; Mergeay, M., Van Houdt, R., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 21–44. [Google Scholar]
- Mijnendonckx, K.; Provoost, A.; Monsieurs, P.; Leys, N.; Mergeay, M.; Mahillon, J.; Van Houdt, R. Insertion sequence elements in Cupriavidus metallidurans CH34: Distribution and role in adaptation. Plasmid 2011, 65, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Van Houdt, R.; Monchy, S.; Leys, N.; Mergeay, M. New mobile genetic elements in Cupriavidus metallidurans CH34, their possible roles and occurrence in other bacteria. Antonie Leeuwenhoek 2009, 96, 205–226. [Google Scholar] [CrossRef] [PubMed]
- Juhas, M.; van der Meer, J.R.; Gaillard, M.; Harding, R.M.; Hood, D.W.; Crook, D.W. Genomic islands: Tools of bacterial horizontal gene transfer and evolution. FEMS Microbiol. Rev. 2009, 33, 376–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandecraen, J.; Chandler, M.; Aertsen, A.; Van Houdt, R. The impact of insertion sequences on bacterial genome plasticity and adaptability. Crit. Rev. Microbiol. 2017, 43, 709–730. [Google Scholar] [CrossRef] [PubMed]
- Obeng, N.; Pratama, A.A.; Elsas, J.D.V. The significance of mutualistic phages for bacterial ecology and evolution. Trends Microbiol. 2016, 24, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Bondy-Denomy, J.; Davidson, A.R. When a virus is not a parasite: The beneficial effects of prophages on bacterial fitness. J. Microbiol. 2014, 52, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Bochner, B.R.; Gadzinski, P.; Panomitros, E. Phenotype microarrays for high-throughput phenotypic testing and assay of gene function. Genom. Res. 2001, 11, 1246–1255. [Google Scholar] [CrossRef] [PubMed]
- Mergeay, M.; Houba, C.; Gerits, J. Extrachromosomal inheritance controlling resistance to cadmium, cobalt, copper and zinc ions: Evidence from curing a Pseudomonas. Arch. Int. Physiol. Biochim. Biophys. 1978, 86, 440–442. [Google Scholar]
- Ferrieres, L.; Hemery, G.; Nham, T.; Guerout, A.M.; Mazel, D.; Beloin, C.; Ghigo, J.M. Silent mischief: Bacteriophage Mu insertions contaminate products of Escherichia coli random mutagenesis performed using suicidal transposon delivery plasmids mobilized by broad-host-range RP4 conjugative machinery. J. Bacteriol. 2010, 192, 6418–6427. [Google Scholar] [CrossRef] [PubMed]
- Schafer, A.; Tauch, A.; Jager, W.; Kalinowski, J.; Thierbach, G.; Puhler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: Selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef]
- Chang, A.C.; Cohen, S.N. Construction and characterization of amplifiable multicopy DNA cloning vehicles derived from the p15A cryptic miniplasmid. J. Bacteriol. 1978, 134, 1141–1156. [Google Scholar] [PubMed]
- Blatny, J.M.; Brautaset, T.; Winther-Larsen, H.C.; Haugan, K.; Valla, S. Construction and use of a versatile set of broad-host-range cloning and expression vectors based on the RK2 replicon. Appl. Environ. Microbiol. 1997, 63, 370–379. [Google Scholar] [PubMed]
- Andrup, L.; Barfod, K.K.; Jensen, G.B.; Smidt, L. Detection of large plasmids from the Bacillus cereus group. Plasmid 2008, 59, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Vaas, L.A.; Sikorski, J.; Hofner, B.; Fiebig, A.; Buddruhs, N.; Klenk, H.P.; Goker, M. opm: An R package for analysing OmniLog® phenotype microarray data. Bioinformatics 2013, 29, 1823–1824. [Google Scholar] [CrossRef] [PubMed]
- Vaas, L.A.; Sikorski, J.; Michael, V.; Goker, M.; Klenk, H.P. Visualization and curve-parameter estimation strategies for efficient exploration of phenotype microarray kinetics. PLoS ONE 2012, 7, e34846. [Google Scholar] [CrossRef] [PubMed]
- Vallenet, D.; Belda, E.; Calteau, A.; Cruveiller, S.; Engelen, S.; Lajus, A.; Le Fevre, F.; Longin, C.; Mornico, D.; Roche, D.; et al. MicroScope—An integrated microbial resource for the curation and comparative analysis of genomic and metabolic data. Nucleic Acids Res. 2013, 41, D636–647. [Google Scholar] [CrossRef] [PubMed]
- Miele, V.; Penel, S.; Duret, L. Ultra-fast sequence clustering from similarity networks with SiLiX. BMC Bioinf. 2011, 12, 116. [Google Scholar] [CrossRef] [PubMed]
- Ondov, B.D.; Treangen, T.J.; Melsted, P.; Mallonee, A.B.; Bergman, N.H.; Koren, S.; Phillippy, A.M. Mash: Fast genome and metagenome distance estimation using MinHash. Genom. Biol. 2016, 17, 132. [Google Scholar] [CrossRef] [PubMed]
- McArthur, A.G.; Waglechner, N.; Nizam, F.; Yan, A.; Azad, M.A.; Baylay, A.J.; Bhullar, K.; Canova, M.J.; De Pascale, G.; Ejim, L.; et al. The comprehensive antibiotic resistance database. Antimicrob. Agents Chemother. 2013, 57, 3348–3357. [Google Scholar] [CrossRef] [PubMed]
- McArthur, A.G.; Wright, G.D. Bioinformatics of antimicrobial resistance in the age of molecular epidemiology. Curr. Opin. Microbiol. 2015, 27, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2017, 45, D566–D573. [Google Scholar] [CrossRef] [PubMed]
- Pal, C.; Bengtsson-Palme, J.; Rensing, C.; Kristiansson, E.; Larsson, D.G. BacMet: Antibacterial biocide and metal resistance genes database. Nucleic Acids Res. 2014, 42, D737–D743. [Google Scholar] [CrossRef] [PubMed]
- Monsieurs, P.; Provoost, A.; Mijnendonckx, K.; Leys, N.; Gaudreau, C.; Van Houdt, R. Genome sequence of Cupriavidus metallidurans Strain H1130, isolated from an invasive human infection. Genom. Announc. 2013, 1, e01051-13. [Google Scholar] [CrossRef] [PubMed]
- Monsieurs, P.; Mijnendonckx, K.; Provoost, A.; Venkateswaran, K.; Ott, C.M.; Leys, N.; Van Houdt, R. Genome sequences of Cupriavidus metallidurans strains NA1, NA4, and NE12, isolated from space equipment. Genom. Announc. 2014, 2, e00719-14. [Google Scholar] [CrossRef] [PubMed]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef] [PubMed]
- Varani, A.M.; Siguier, P.; Gourbeyre, E.; Charneau, V.; Chandler, M. ISsaga is an ensemble of web-based methods for high throughput identification and semi-automatic annotation of insertion sequences in prokaryotic genomes. Genom. Biol. 2011, 12, R30. [Google Scholar] [CrossRef] [PubMed]
- Ricker, N.; Qian, H.; Fulthorpe, R.R. The limitations of draft assemblies for understanding prokaryotic adaptation and evolution. Genomics 2012, 100, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Sadouk, A.; van der Lelie, D.; Taghavi, S.; Ferhat, A.; Nuyten, J.M.; Borremans, B.; Mergeay, M.; Toussaint, A. Cloning and sequencing of IS1086, an Alcaligenes eutrophus insertion element related to IS30 and IS4351. J. Bacteriol. 1992, 174, 8133–8138. [Google Scholar] [CrossRef] [PubMed]
- Collard, J.M.; Provoost, A.; Taghavi, S.; Mergeay, M. A new type of Alcaligenes eutrophus CH34 zinc resistance generated by mutations affecting regulation of the cnr cobalt-nickel resistance system. J. Bacteriol. 1993, 175, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Grass, G.; Grosse, C.; Nies, D.H. Regulation of the cnr cobalt and nickel resistance determinant from Ralstonia sp. strain CH34. J. Bacteriol. 2000, 182, 1390–1398. [Google Scholar] [CrossRef] [PubMed]
- Talat, M.-E. Genetic Mechanism of Heavy Metal Resistance of Pseudomonas aeruginosa CMG103. Ph.D. Thesis, University of Karachi, Karachi, Pakistan, 2000. [Google Scholar]
- Schneider, D.; Faure, D.; Noirclerc-Savoye, M.; Barriere, A.C.; Coursange, E.; Blot, M. A broad-host-range plasmid for isolating mobile genetic elements in gram-negative bacteria. Plasmid 2000, 44, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Tibazarwa, C.; Wuertz, S.; Mergeay, M.; Wyns, L.; van der Lelie, D. Regulation of the cnr cobalt and nickel resistance determinant of Ralstonia eutropha (Alcaligenes eutrophus) CH34. J. Bacteriol. 2000, 182, 1399–1409. [Google Scholar] [CrossRef] [PubMed]
- Vandecraen, J.; Monsieurs, P.; Mergeay, M.; Leys, N.; Aertsen, A.; Van Houdt, R. Zinc-induced transposition of insertion sequence elements contributes to increased adaptability of Cupriavidus metallidurans. Front. Microbiol. 2016, 7, 359. [Google Scholar] [CrossRef] [PubMed]
- Providenti, M.A.; Shaye, R.E.; Lynes, K.D.; McKenna, N.T.; O’Brien, J.M.; Rosolen, S.; Wyndham, R.C.; Lambert, I.B. The locus coding for the 3-nitrobenzoate dioxygenase of Comamonas sp. strain JS46 is flanked by IS1071 elements and is subject to deletion and inversion events. Appl. Environ. Microbiol. 2006, 72, 2651–2660. [Google Scholar] [CrossRef] [PubMed]
- Clement, P.; Pieper, D.H.; Gonzalez, B. Molecular characterization of a deletion/duplication rearrangement in tfd genes from Ralstonia eutropha JMP134(pJP4) that improves growth on 3-chlorobenzoic acid but abolishes growth on 2,4-dichlorophenoxyacetic acid. Microbiology 2001, 147, 2141–2148. [Google Scholar] [CrossRef] [PubMed]
- Mindlin, S.; Petrova, M. Mercury resistance transposons. In Bacterial Integrative Mobile Genetic Elements; Roberts, A.P., Mullany, P., Eds.; Landes Biosciences: Austin, TX, USA, 2013; pp. 33–52. [Google Scholar]
- Miyazaki, R.; Minoia, M.; Pradervand, N.; Sentchilo, V.; Sulser, S.; Reinhard, F.; van der Meer, J.R. The clc Element and Related Genomic Islands in Proteobacteria. In Bacterial Integrative Mobile Genetic Elements; Roberts, A.P., Mullany, P., Eds.; Landes Bioscience: Austin, TX, USA, 2013; pp. 261–272. [Google Scholar]
- Klockgether, J.; Wurdemann, D.; Reva, O.; Wiehlmann, L.; Tummler, B. Diversity of the abundant pKLC102/PAGI-2 family of genomic islands in Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 2443–2459. [Google Scholar] [CrossRef] [PubMed]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [PubMed]
- Van Houdt, R.; Toussaint, A.; Ryan, M.P.; Pembroke, J.T.; Mergeay, M.; Adley, C.C. The Tn4371 ICE Family of Bacterial Mobile Genetic Elements. In Bacterial Integrative Mobile Genetic Elements; Roberts, A.P., Mullany, P., Eds.; Landes Bioscience: Austin, TX, USA, 2013; pp. 179–200. [Google Scholar]
- Smith, D.J.; Park, J.; Tiedje, J.M.; Mohn, W.W. A large gene cluster in Burkholderia xenovorans encoding abietane diterpenoid catabolism. J. Bacteriol. 2007, 189, 6195–6204. [Google Scholar] [CrossRef] [PubMed]
- Byun-McKay, A.; Godard, K.A.; Toudefallah, M.; Martin, D.M.; Alfaro, R.; King, J.; Bohlmann, J.; Plant, A.L. Wound-induced terpene synthase gene expression in Sitka spruce that exhibit resistance or susceptibility to attack by the white pine weevil. Plant Physiol. 2006, 140, 1009–1021. [Google Scholar] [CrossRef] [PubMed]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [PubMed]
- Sorek, R.; Lawrence, C.M.; Wiedenheft, B. CRISPR-mediated adaptive immune systems in bacteria and archaea. Annu. Rev. Biochem. 2013, 82, 237–266. [Google Scholar] [CrossRef] [PubMed]
- Grissa, I.; Vergnaud, G.; Pourcel, C. CRISPRFinder: A web tool to identify clustered regularly interspaced short palindromic repeats. Nucleic Acids Res. 2007, 35, W52–W57. [Google Scholar] [CrossRef] [PubMed]
- Biswas, A.; Staals, R.H.; Morales, S.E.; Fineran, P.C.; Brown, C.M. CRISPRDetect: A flexible algorithm to define CRISPR arrays. BMC Genom. 2016, 17, 356. [Google Scholar] [CrossRef] [PubMed]
- Biswas, A.; Fineran, P.C.; Brown, C.M. Computational Detection of CRISPR/crRNA Targets. Methods Mol. Biol. 2015, 1311, 77–89. [Google Scholar] [PubMed]
- Monsieurs, P.; Moors, H.; Van Houdt, R.; Janssen, P.J.; Janssen, A.; Coninx, I.; Mergeay, M.; Leys, N. Heavy metal resistance in Cupriavidus metallidurans CH34 is governed by an intricate transcriptional network. BioMetals 2011, 24, 1133–1151. [Google Scholar] [CrossRef] [PubMed]
- Henne, K.L.; Nakatsu, C.H.; Thompson, D.K.; Konopka, A.E. High-level chromate resistance in Arthrobacter sp. strain FB24 requires previously uncharacterized accessory genes. BMC Microbiol. 2009, 9, 199. [Google Scholar] [CrossRef] [PubMed]
- Henne, K.L.; Turse, J.E.; Nicora, C.D.; Lipton, M.S.; Tollaksen, S.L.; Lindberg, C.; Babnigg, G.; Giometti, C.S.; Nakatsu, C.H.; Thompson, D.K.; et al. Global proteomic analysis of the chromate response in Arthrobacter sp. strain FB24. J. Proteome Res. 2009, 8, 1704–1716. [Google Scholar] [CrossRef] [PubMed]
- Nies, D.H.; Rehbein, G.; Hoffmann, T.; Baumann, C.; Grosse, C. Paralogs of genes encoding metal resistance proteins in Cupriavidus metallidurans strain CH34. J. Mol. Microbiol. Biotechnol. 2006, 11, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Hlozkova, K.; Suman, J.; Strnad, H.; Ruml, T.; Paces, V.; Kotrba, P. Characterization of pbt genes conferring increased Pb2+ and Cd2+ tolerance upon Achromobacter xylosoxidans A8. Res. Microbiol. 2013, 164, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Wiesemann, N.; Mohr, J.; Grosse, C.; Herzberg, M.; Hause, G.; Reith, F.; Nies, D.H. Influence of copper resistance determinants on gold transformation by Cupriavidus metallidurans strain CH34. J. Bacteriol. 2013, 195, 2298–2308. [Google Scholar] [CrossRef] [PubMed]
- Reith, F.; Etschmann, B.; Grosse, C.; Moors, H.; Benotmane, M.A.; Monsieurs, P.; Grass, G.; Doonan, C.; Vogt, S.; Lai, B.; et al. Mechanisms of gold biomineralization in the bacterium Cupriavidus metallidurans. Proc. Natl. Acad. Sci. USA 2009, 106, 17757–17762. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.; Schlegel, H.G. Combined nickel-cobalt-cadmium resistance encoded by the ncc locus of Alcaligenes xylosoxidans 31A. J. Bacteriol. 1994, 176, 7045–7054. [Google Scholar] [CrossRef] [PubMed]
- Orita, I.; Iwazawa, R.; Nakamura, S.; Fukui, T. Identification of mutation points in Cupriavidus necator NCIMB 11599 and genetic reconstitution of glucose-utilization ability in wild strain H16 for polyhydroxyalkanoate production. J. Biosci. Bioeng. 2012, 113, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Raberg, M.; Peplinski, K.; Heiss, S.; Ehrenreich, A.; Voigt, B.; Doring, C.; Bomeke, M.; Hecker, M.; Steinbuchel, A. Proteomic and transcriptomic elucidation of the mutant Ralstonia eutropha G+1 with regard to glucose utilization. Appl. Environ. Microbiol. 2011, 77, 2058–2070. [Google Scholar] [CrossRef] [PubMed]
- Kurnasov, O.; Jablonski, L.; Polanuyer, B.; Dorrestein, P.; Begley, T.; Osterman, A. Aerobic tryptophan degradation pathway in bacteria: Novel kynurenine formamidase. FEMS Microbiol. Lett. 2003, 227, 219–227. [Google Scholar] [CrossRef]
- Weimann, A.; Mooren, K.; Frank, J.; Pope, P.B.; Bremges, A.; McHardy, A.C. From genomes to phenotypes: Traitar, the microbial trait analyzer. mSystems 2016, 1, e00101-16. [Google Scholar] [CrossRef] [PubMed]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Plasmid | Genotype/Relevant Characteristics | Reference |
|---|---|---|
| STRAIN | ||
| Cupriavidus metallidurans | ||
| CH34T | Type strain | [31] |
| NA1 | Isolated from a water sample, ISS | [12] |
| NA4 | Isolated from a water sample, ISS | [12] |
| NA4 ΔCRISPR | ΔCRISPR::tet, TcR | This study |
| H1130 | Isolated from invasive human infection | [15] |
| Escherichia coli | ||
| DG1 | mcrA Δmrr-hsdRMS-mcrBC (rB−mB−) Φ80lacZΔM15 ΔlacX74 recA1 araD139 Δ(ara-leu)7697 galU galK rpsL endA1 nupG | Eurogentec |
| MFDpir | MG1655 RP4-2-Tc::[ΔMu1::aac(3)IV-ΔaphA-Δnic35-ΔMu2::zeo] ΔdapA::(erm-pir) ΔrecA | [32] |
| PLASMID | ||
| pK18mob | pMB1 ori, mob+, lacZ, KmR | [33] |
| pK18mob-CRISPR | CRISPR region of NA4 in pK18mob, KmR | This study |
| pK18mob-CRISPR::tet | pK18mob-CRISPR derivative, CRISPR::tet, KmR, TcR | This study |
| pACYC184 | p15A ori, CmR, TcR | [34] |
| pJB3kan1 | RK2 minimal replicon; ApR, KmR | [35] |
| pJB3kan1_Rmet2825 | Rmet_2825 of CH34 in pJB3kan1; KmR | This study |
| Strain | Size a | Completeness b | Score c | # d | Position | Most Common Phage e | GC % |
|---|---|---|---|---|---|---|---|
| NA1 | 27.9 | questionable | 90 | 32 | 528,474–556,451 | Ralsto_RS138 (NC_029107; 7) | 65.35 |
| 17.7 | incomplete | 20 | 21 | 554,542–572,263 | Pseudo_NP1 (NC_031058; 3) | 64.23 | |
| NA4 | 43.6 | intact | 100 | 50 | 1,706,628–1,750,233 | Bordet_BPP_1 (NC_005357; 18) | 64.94 |
| 6.1 | intact | 100 | 10 | 1,941,664–1,947,835 | Ralsto_PE226 (NC_015297; 6) | 60.08 | |
| 45.2 | intact | 150 | 41 | 2,145,854–2,191,126 | Burkho_Bcep176 (NC_007497; 11) | 61.83 | |
| 8 | incomplete | 30 | 10 | 2,181,367–2,189,450 | Gordon_Nymphadora (NC_031061; 2) | 62.44 | |
| 120.5 | intact | 130 | 125 | 2,248,504–2,369,042 | Salmon_118970_sal3 (NC_031940; 14) | 61.84 | |
| 44.5 | incomplete | 30 | 39 | 2,545,435–2,589,959 | Pseudo_JBD44 (NC_030929; 5) | 63.89 | |
| H1130 | 19.3 | incomplete | 30 | 21 | 1,470,543–1,489,908 | Burkho_phiE125 (NC_003309; 3) | 61.27 |
| 12.7 | incomplete | 40 | 19 | 1,505,965–1,518,710 | Bacill_SP_15 (NC_031245; 5) | 63.39 | |
| 7.9 | incomplete | 30 | 9 | 2,763,200–2,771,108 | Entero_phi92 (NC_023693; 4) | 58.60 | |
| 48.4 | intact | 110 | 73 | 6,748,679–6,797,173 | Salmon_SEN34 (NC_028699; 22) | 62.09 | |
| 16 | incomplete | 50 | 29 | 7,110,429–7,126,525 | Clostr_phiCT453B (NC_029004; 4) | 61.04 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Houdt, R.; Provoost, A.; Van Assche, A.; Leys, N.; Lievens, B.; Mijnendonckx, K.; Monsieurs, P. Cupriavidus metallidurans Strains with Different Mobilomes and from Distinct Environments Have Comparable Phenomes. Genes 2018, 9, 507. https://doi.org/10.3390/genes9100507
Van Houdt R, Provoost A, Van Assche A, Leys N, Lievens B, Mijnendonckx K, Monsieurs P. Cupriavidus metallidurans Strains with Different Mobilomes and from Distinct Environments Have Comparable Phenomes. Genes. 2018; 9(10):507. https://doi.org/10.3390/genes9100507
Chicago/Turabian StyleVan Houdt, Rob, Ann Provoost, Ado Van Assche, Natalie Leys, Bart Lievens, Kristel Mijnendonckx, and Pieter Monsieurs. 2018. "Cupriavidus metallidurans Strains with Different Mobilomes and from Distinct Environments Have Comparable Phenomes" Genes 9, no. 10: 507. https://doi.org/10.3390/genes9100507
APA StyleVan Houdt, R., Provoost, A., Van Assche, A., Leys, N., Lievens, B., Mijnendonckx, K., & Monsieurs, P. (2018). Cupriavidus metallidurans Strains with Different Mobilomes and from Distinct Environments Have Comparable Phenomes. Genes, 9(10), 507. https://doi.org/10.3390/genes9100507