Genome-Wide Identification and Characterization of wALOG Family Genes Involved in Branch Meristem Development of Branching Head Wheat

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Scanning Electron Microscopy
2.3. RNA Isolation and RNA Sequencing
2.4. RNA-seq Data Processing
2.5. Isolation of ALOG Family Genes
2.6. Multiple Sequence Alignment, Motif Identification, and Phylogenetic Analysis
2.7. Cis-Acting Regulatory Elements Prediction in the Promoter Regions of wALOG Genes
2.8. qRT-PCR Analysis
2.9. Cloning of BM Development-Associated TtALOG Genes and its Promoters
3. Results
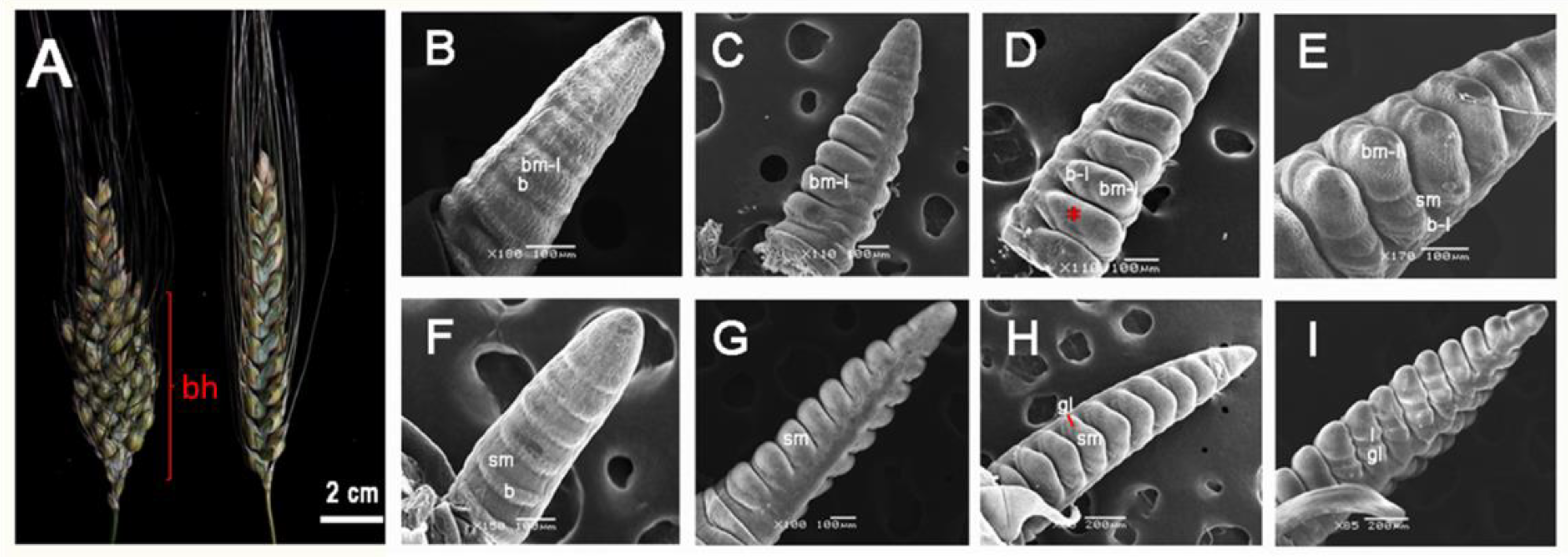
3.1. Morphological Differences and Lateral BM Initiation of Branching Head Wheat
3.2. RNA-Seq Analysis of Early Stages of Inflorescence Development in GAN-A631
3.3. Expansion of the wALOG Family Genes During Wheat Polyploidization
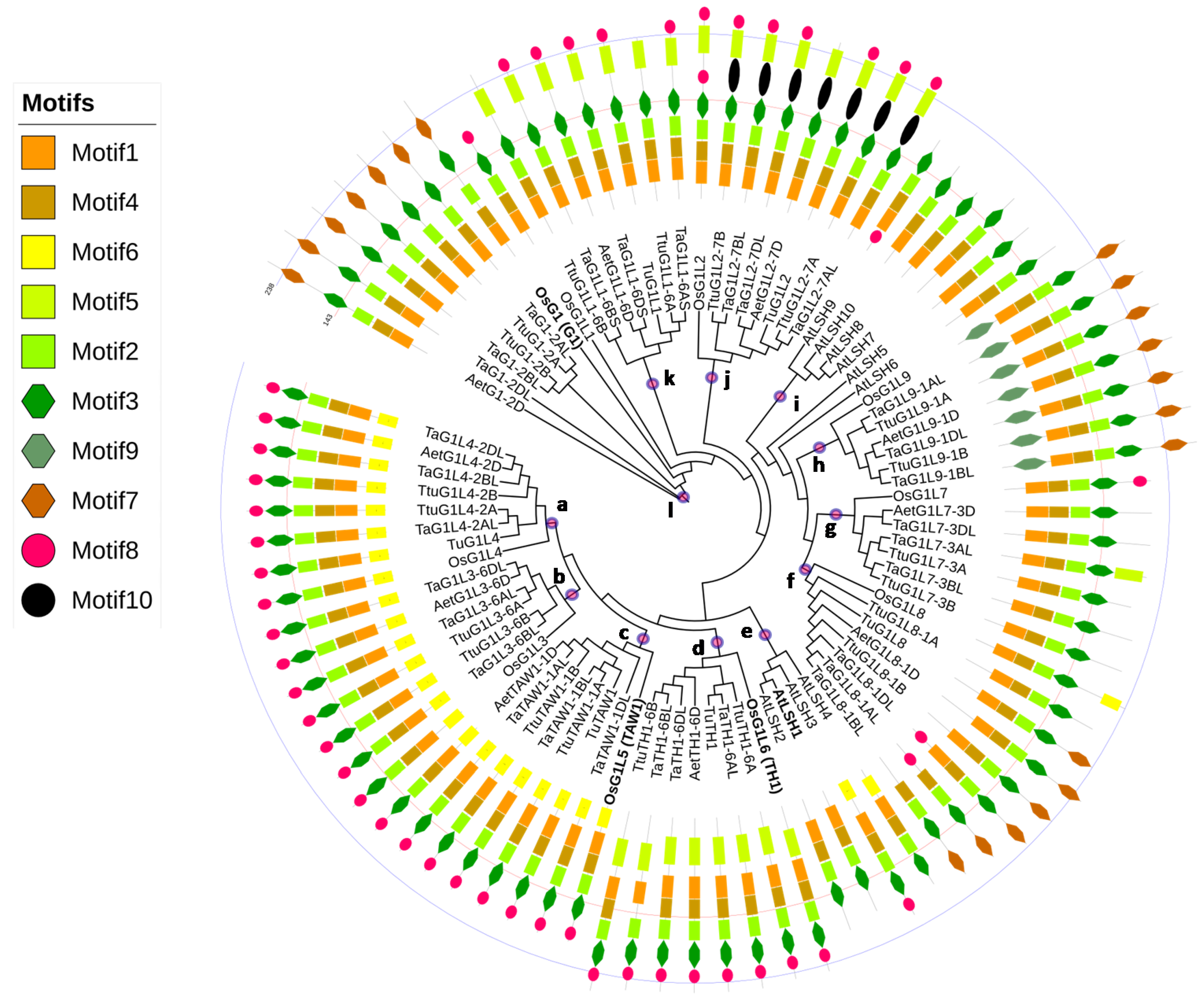
3.4. Phylogenetic Analysis and Classification of the wALOG Genes
3.5. Prediction of Cis-Acting Regulatory Elements in the Promoter Regions of wALOGs
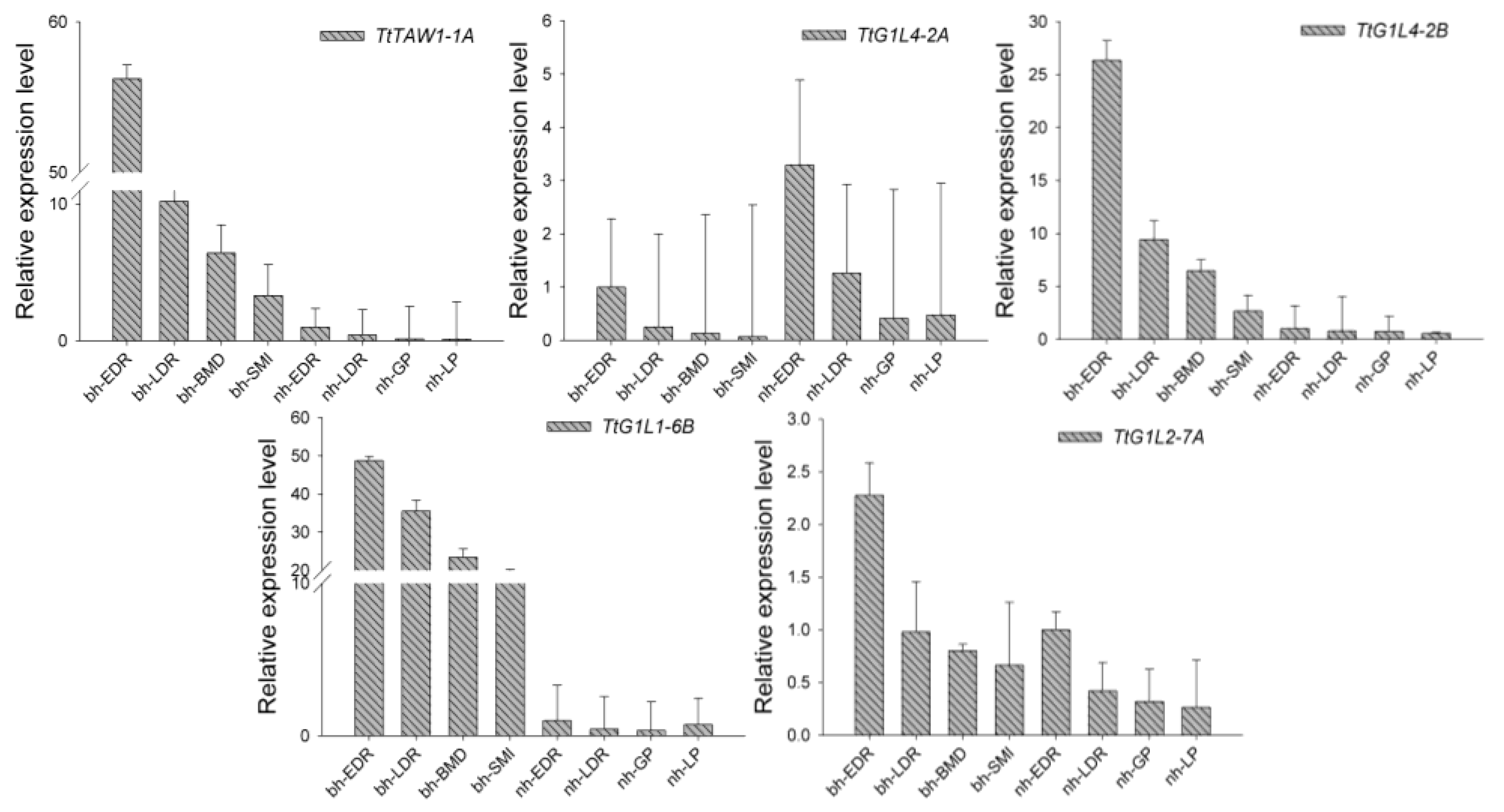
3.6. Expression of the TtALOG Genes and Possibly their Downstream Genes Relate to BM Development
3.7. Characterization of Protein Sequences and Promoters of TtALOG Genes Involved in BM Development
4. Discussion
4.1. The Development Pattern of the Branch-Like Meristems of GAN-A631 is Similar to the Indeterminate BMs of Rice
4.2. Copy Number Variation and Functional Diversification of wALOGs During Polyploidization
4.3. TtALOGs are Attractive Candidates for Promoting Indeterminate BM Fate
4.4. Light and Phytohormones May Participate in the Regulation of wALOGs
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ashikari, M.; Sakakibara, H.; Lin, S.; Yamamoto, T.; Takashi, T.; Nishimura, A.; Angeles, E.R.; Qian, Q.; Kitano, H.; Matsuoka, M. Cytokinin oxidase regulates rice grain production. Science 2005, 309, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Hake, S. Inflorescence architecture: The transition from branches to flowers. Curr. Biol. 2008, 18, R1106–R1108. [Google Scholar] [CrossRef] [PubMed]
- Hirano, H.-Y.; Tanaka, W.; Toriba, T. Grass flower development. In Flower Development: Methods and Protocols; Humana Press: New York, NY, USA, 2014; pp. 57–84. [Google Scholar] [CrossRef]
- Bommert, P.; Whipple, C. Grass inflorescence architecture and meristem determinacy. Semin. Cell Dev. Biol. 2017, 79, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, A.; Sasao, M.; Yasuno, N.; Takagi, K.; Daimon, Y.; Chen, R.; Yamazaki, R.; Tokunaga, H.; Kitaguchi, Y.; Sato, Y. TAWAWA1, a regulator of rice inflorescence architecture, functions through the suppression of meristem phase transition. Proc. Natl. Acad. Sci. USA 2013, 110, 767–772. [Google Scholar] [CrossRef] [PubMed]
- Kyozuka, J.; Tokunaga, H.; Yoshida, A. Control of grass inflorescence form by the fine-tuning of meristem phase change. Curr. Opin. Plant Biol. 2014, 17, 110. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, M.; Chujo, A.; Nagato, Y.; Shimamoto, K.; Kyozuka, J. FRIZZY PANICLE is required to prevent the formation of axillary meristems and to establish floral meristem identity in rice spikelets. Development 2003, 130, 3841–3850. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Nagasawa, N.; Nagato, Y. ABERRANT PANICLE ORGANIZATION 1 temporally regulates meristem identity in rice. Dev. Biol. 2005, 282, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Ikeda-Kawakatsu, K.; Maekawa, M.; Izawa, T.; Itoh, J.I.; Nagato, Y. ABERRANT PANICLE ORGANIZATION 2/RFL, the rice ortholog of Arabidopsis LEAFY, suppresses the transition from inflorescence meristem to floral meristem through interaction with APO1. Plant J. 2012, 69, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Murai, K.; Takumi, S.; Koga, H.; Ogihara, Y. Pistillody, homeotic transformation of stamens into pistil-like structures, caused by nuclear-cytoplasm interaction in wheat. Plant J. 2002, 29, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Shitsukawa, N.; Takagishi, A.; Ikari, C.; Takumi, S.; Murai, K. WFL, a wheat FLORICAULA/LEAFY ortholog, is associated with spikelet formation as lateral branch of the inflorescence meristem. Genes Genet. Syst. 2006, 81, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Boden, S.A.; Cavanagh, C.; Cullis, B.R.; Ramm, K.; Greenwood, J.; Finnegan, E.J.; Trevaskis, B.; Swain, S.M. Ppd-1 is a key regulator of inflorescence architecture and paired spikelet development in wheat. Nat. Plants 2015, 1, 14016. [Google Scholar] [CrossRef] [PubMed]
- Dobrovolskaya, O.; Pont, C.; Sibout, R.; Martinek, P.; Badaeva, E.; Murat, F.; Chosson, A.; Watanabe, N.; Prat, E.; Gautier, N. FRIZZY PANICLE drives supernumerary spikelets in bread wheat. Plant Physiol. 2015, 167, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Poursarebani, N.; Seidensticker, T.; Koppolu, R.; Trautewig, C.; Gawroński, P.; Bini, F.; Govind, G.; Rutten, T.; Sakuma, S.; Tagiri, A. The genetic basis of composite spike form in barley and ‘Miracle-Wheat’. Genetics 2015, 201, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Nakazawa, M.; Takase, T.; Manabe, K.; Kobayashi, M.; Seki, M.; Shinozaki, K.; Matsui, M. Overexpression of LSH1, a member of an uncharacterised gene family, causes enhanced light regulation of seedling development. Plant J. 2004, 37, 694–706. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, A.; Suzaki, T.; Tanaka, W.; Hirano, H.-Y. The homeotic gene long sterile lemma (G1) specifies sterile lemma identity in the rice spikelet. Proc. Natl. Acad. Sci. USA 2009, 106, 20103–20108. [Google Scholar] [CrossRef] [PubMed]
- Zhi, W.N.T.; Song, S.; Wang, Y.Q.; Liu, J.; Yu, H. New insights into the regulation of inflorescence architecture. Trends Plant Sci. 2014, 19, 158–165. [Google Scholar] [CrossRef]
- Peng, P.; Liu, L.; Fang, J.; Zhao, J.; Yuan, S.; Li, X. The rice TRIANGULAR HULL1 protein acts as a transcriptional repressor in regulating lateral development of spikelet. Sci. Rep. 2017, 7, 13712. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Hanano, K.; Kariya, A.; Shimizu, S.; Zhao, L.; Matsui, M.; Tasaka, M.; Aida, M. CUP-SHAPED COTYLEDON1 transcription factor activates the expression of LSH4 and LSH3, two members of the ALOG gene family, in shoot organ boundary cells. Plant J. 2011, 66, 1066–1077. [Google Scholar] [CrossRef] [PubMed]
- Hong, L.; Qian, Q.; Zhu, K.; Tang, D.; Huang, Z.; Gao, L.; Li, M.; Gu, M.; Cheng, Z. ELE restrains empty glumes from developing into lemmas. J. Genet. Genom. 2010, 37, 101–115. [Google Scholar] [CrossRef]
- Yoshida, H.; Nagato, Y. Flower development in rice. J. Exp. Bot. 2011, 62, 4719–4730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Li, H.; Su, Y.; Li, W.; Shi, C. G1/ELE functions in the development of rice lemmas in addition to determining identities of empty glumes. Front. Plant Sci. 2016, 7, 1006. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Cheng, Z.; Wu, F.; Jin, M.; Zhang, L.; Zhou, F.; Wang, J.; Zhou, K.; Ma, J.; Lin, Q. BEAK LIKE SPIKELET1 is required for lateral development of lemma and Palea in rice. Plant Mol. Biol. Rep. 2013, 31, 98–108. [Google Scholar] [CrossRef]
- Yan, D.; Zhou, Y.; Ye, S.; Zeng, L.; Zhang, X.; He, Z. BEAK-SHAPED GRAIN 1/TRIANGULAR HULL 1, a DUF640 gene, is associated with grain shape, size and weight in rice. Sci. China Life Sci. 2013, 56, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Rao, Y.; Wu, L.; Xu, Q.; Li, Z.; Yu, H.; Zhang, Y.; Leng, Y.; Hu, J.; Zhu, L. The pleiotropic ABNORMAL FLOWER AND DWARF1 affects plant height, floral development and grain yield in rice. J. Integr. Plant Biol. 2016, 58, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A.; Coggill, P.; Finn, R.D. DUFs: Families in search of function. Acta Crystallogr. 2010, 66, 1148–1152. [Google Scholar] [CrossRef] [PubMed]
- Iyer, L.M.; Aravind, L. ALOG domains: Provenance of plant homeotic and developmental regulators from the DNA-binding domain of a novel class of DIRS1-type retroposons. Biol. Direct 2012, 7, 39. [Google Scholar] [CrossRef] [PubMed]
- MacAlister, C.A.; Park, S.J.; Jiang, K.; Marcel, F.; Bendahmane, A.; Izkovich, Y.; Eshed, Y.; Lippman, Z.B. Synchronization of the flowering transition by the tomato TERMINATING FLOWER gene. Nat. Genet. 2012, 44, 1393–1398. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Lee, J.; Nou, I.-S.; Hur, Y. Expression Characteristics of LSH Genes in Brassica Suggest their Applicability for Modification of Leaf Morphology and the Use of their Promoter for Transgenesis. Plant Breed. Biotechnol. 2014, 2, 126–138. [Google Scholar] [CrossRef]
- Marcussen, T.; Sandve, S.R.; Heier, L.; Spannagl, M.; Pfeifer, M.; Jakobsen, K.S.; Wulff, B.B.; Steuernagel, B.; Mayer, K.F.; Olsen, O.A. Ancient hybridizations among the ancestral genomes of bread wheat. Science 2014, 345, 1250092. [Google Scholar] [CrossRef] [PubMed]
- Mayer, K.F.X.; Rogers, J.; Doležel, J.; Pozniak, C.; Eversole, K.; Feuillet, C.; Gill, B.; Friebe, B.; Lukaszewski, A.J.; Sourdille, P. A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum) genome. Science 2014, 345, 1251788. [Google Scholar] [CrossRef]
- Soltis, P.S.; Soltis, D.E. The role of genetic and genomic attributes in the success of polyploids. Proc. Natl. Acad. Sci. USA 2000, 97, 7051–7057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendel, J.F. Genome evolution in polyploids. Plant Mol. Biol. 2000, 42, 225–249. [Google Scholar] [CrossRef] [PubMed]
- Gautier, M.F.; Cosson, P.; Guirao, A.; Alary, R.; Joudrier, P. Puroindoline genes are highly conserved in diploid ancestor wheats and related species but absent in tetraploid Triticum species. Plant Sci. 2000, 153, 81–91. [Google Scholar] [CrossRef]
- Feuillet, C.; Penger, A.; Gellner, K.; Mast, A.; Keller, B. Molecular evolution of receptor-like kinase genes in hexaploid wheat. Independent evolution of orthologs after polyploidization and mechanisms of local rearrangements at paralogous loci. Plant Physiol. 2001, 125, 1304–1313. [Google Scholar] [CrossRef] [PubMed]
- Kashkush, K.; Feldman, M.; Levy, A.A. Gene loss, silencing and activation in a newly synthesized wheat allotetraploid. Genetics 2002, 160, 1651–1659. [Google Scholar] [PubMed]
- Ling, H.Q.; Ma, B.; Shi, X.; Liu, H.; Dong, L.; Sun, H.; Cao, Y.; Gao, Q.; Zheng, S.; Li, Y. Genome sequence of the progenitor of wheat A subgenome Triticum urartu. Nature 2018, 557, 424–428. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.-C.; Gu, Y.Q.; Puiu, D.; Wang, H.; Twardziok, S.O.; Deal, K.R.; Huo, N.; Zhu, T.; Wang, L.; Wang, Y. Genome sequence of the progenitor of the wheat D genome Aegilops tauschii. Nature 2017, 551, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Avni, R.; Nave, M.; Barad, O.; Baruch, K.; Twardziok, S.O.; Gundlach, H.; Hale, I.; Mascher, M.; Spannagl, M.; Wiebe, K. Wild emmer genome architecture and diversity elucidate wheat evolution and domestication. Science 2017, 357, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Clavijo, B.J.; Venturini, L.; Schudoma, C.; Accinelli, G.G.; Kaithakottil, G.; Wright, J.; Borrill, P.; Kettleborough, G.; Heavens, D.; Chapman, H. An improved assembly and annotation of the allohexaploid wheat genome identifies complete families of agronomic genes and provides genomic evidence for chromosomal translocations. Genome Res. 2017, 27, 885–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, A.; Zhong, H.; Wang, W.; Sticklen, M.B. Shoot apical meristem: In vitro regeneration and morphogenesis in wheat (Triticum aestivum L.). In Vitro Cell. Dev. Biol. Plant 2002, 38, 163–167. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- FastQC Software. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 15 November 2017).
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Cufflinks Software. Available online: http://cole-trapnell-lab.github.io/cufflinks/ (accessed on 19 November 2017).
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Heatmap. Available online: https://github.com/jokergoo/ComplexHeatmap/ (accessed on 20 December 2017).
- The Ensembl Plants. Available online: http://plants.ensembl.org/index.html/ (accessed on 28 February 2018).
- The MBKBASE Database. Available online: http://www.mbkbase.org/Tu/ (accessed on 28 February 2018).
- The WEWseq Database. Available online: http://wewseq.wixsite.com/consortium/ (accessed on 28 February 2018).
- Cho, E.; Zambryski, P.C. Organ boundary1 defines a gene expressed at the junction between the shoot apical meristem and lateral organs. Proc. Natl. Acad. Sci. USA 2011, 108, 2154–2159. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Mistry, J.; Schuster-Böckler, B.; Griffiths-Jones, S.; Hollich, V.; Lassmann, T.; Moxon, S.; Marshall, M.; Khanna, A.; Durbin, R.; et al. PFAM: Clans, web tools and services. Nucleic Acids Res. 2006, 34, D247–D251. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, T.J.; Eddy, S.R. NHMMER: DNA homology search with profile HMMs. Bioinformatics 2013, 29, 2487–2489. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- The Conserved Domain Database. Available online: https://www.ncbi.nlm.nih.gov/cdd/ (accessed on 4 March 2018).
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- The MEME Suite. Available online: http://meme-suite.org/tools/meme/ (accessed on 5 March 2018).
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Xia, R.; Chen, H.; He, Y. TBtools, a Toolkit for Biologists integrating various HTS-data handling tools with a user-friendly interface. bioRxiv 2018. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Primer Premier: A Comprehensive PCR Primer Design Software. Available online: http://www. premierbiosoft.com/primerdesign/ (accessed on 2 May 2018).
- Doyle, J.; Doyle, J. Genomic plant DNA preparation from fresh tissue-CTAB method. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- ProtParam Tool. Available online: https://web.expasy.org/protparam/ (accessed on 5 March 2018).
- Hernandez-Garcia, C.M.; Finer, J.J. Identification and validation of promoters and cis-acting regulatory elements. Plant Sci. 2014, 217–218, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.Q.; Oelke, E.A.; Porter, R.A.; Reuter, R.; Liu, Q.Q. Formation of panicles and hermaphroditic florets in wild-rice. Int. J. Plant Sci. 2015, 159, 550–558. [Google Scholar] [CrossRef]
- Bonnett, O.T. The development of the wheat spike. J. Agric. Res. 1936, 53, 445–451. [Google Scholar]
- Adams, K.L.; Wendel, J.F. Polyploidy and genome evolution in plants. Curr. Opin. Plant Biol. 2005, 8, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, R.; Ma, C.; Xue, S.; Liu, Z.; Wang, Z.; Sun, Q.; Cao, J.; Xu, S. Massive expansion and differential evolution of small heat shock proteins with wheat (Triticum aestivum L.) polyploidization. Sci. Rep. 2017, 7, 2581. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Shu, D.T.; Wang, M.; Xing, G.W.; Zhan, H.S.; Du, X.H.; Song, W.N.; Nie, X.J. Genome-Wide Identification and Expression Analysis of the HD-Zip Gene Family in Wheat (Triticum aestivum L.). Genes 2018, 9, 70. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Dhugga, K.S.; Beech, R.; Singh, J. Genome-wide analysis of the cellulose synthase-like (Csl) gene family in bread wheat (Triticum aestivum L.). BMC Plant Biol. 2017, 17, 193. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sun, L.; Tan, L.; Liu, F.; Zhu, Z.; Fu, Y.; Sun, X.; Sun, X.; Xie, D.; Sun, C. TH1, a DUF640 domain-like gene controls lemma and palea development in rice. Plant Mol. Biol. 2012, 78, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Veit, B. Hormone mediated regulation of the shoot apical meristem. Plant Mol. Biol. 2008, 69, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Cao, W.; Dai, T.; Zhou, Q. Effects of exogenous hormones on floret development and grain setin wheat. Plant Growth Regul. 2001, 35, 225–231. [Google Scholar] [CrossRef]
- Cai, Q.; Yuan, Z.; Chen, M.; Yin, C.; Luo, Z.; Zhao, X.; Liang, W.; Hu, J.; Zhang, D. Jasmonic acid regulates spikelet development in rice. Nat. Commun. 2014, 5, 3476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Jiang, X.; He, F.; Zhang, W.; Gao, Q. Comparison of Dynamic Changes of Endogenous Plant Hormones and Hormone-Related Gene Expression between Near Isogenic Lines with Normal Spike and Branched Spike Wheat. J. Agric. Sci. 2015, 7. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, F.; Gao, Q.; Zhang, Y.; Wang, N.; Zhang, W. Auxins Regulations of Branched Spike Development and Expression of TFL, a LEAFY-Like Gene in Branched Spike Wheat (Triticum aestivum). J. Agric. Sci. 2017, 9, 27. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Total Reads | Clean Reads | GC (%) | Q20 (%) | Total Mapped Reads | Ratio (%) |
|---|---|---|---|---|---|---|
| EDR_1 | 72,894,272 | 70,725,372 | 54.97 | 97.31 | 64,059,309 | 90.57% |
| EDR_2 | 67,545,684 | 65,579,864 | 54.88 | 96.23 | 58,973,520 | 89.93% |
| EDR_3 | 51,128,478 | 50,309,938 | 52.31 | 96.19 | 46,677,546 | 92.78% |
| LDR_1 | 66,259,258 | 64,664,774 | 55.30 | 97.24 | 58,684,655 | 90.75% |
| LDR_2 | 68,195,516 | 66,651,562 | 55.49 | 96.09 | 59,891,457 | 89.86% |
| LDR_3 | 47,152,170 | 46,527,636 | 52.47 | 96.45 | 43,373,007 | 93.22% |
| BMD_1 | 66,756,912 | 65,234,314 | 55.94 | 96.03 | 58,293,385 | 89.36% |
| BMD_2 | 63,734,100 | 63,575,894 | 52.47 | 98.21 | 59,187,411 | 93.10% |
| BMD_3 | 44,215,840 | 42,328,284 | 54.78 | 94.24 | 36,970,389 | 87.34% |
| SMI_1 | 59,302,160 | 57,670,038 | 55.70 | 96.05 | 51,464,811 | 89.24% |
| SMI_2 | 63,615,448 | 62,426,934 | 54.75 | 96.01 | 56,088,433 | 89.85% |
| SMI_3 | 48,891,176 | 48,768,792 | 52.50 | 98.21 | 45,299,165 | 92.89% |
| No. | Phylogenetic Groups | Triticum turgidum ssp. Dicoccoides Genome ID | Gene Name | Gene Name | Ensembl ID | Gene Name | Triticum urartu Genome ID | Gene Name | Aegilops tauschii Genome ID | Gene Name |
|---|---|---|---|---|---|---|---|---|---|---|
| Triticum turgidum ssp. Dicoccoides | Corresponding Gene in Triticum turgidum ssp. Dicoccoides | Corresponding Gene in Triticum turgidum L. | Triticum aestivum | Corresponding Gene in Triticum aestivum | Triticum urartu | Corresponding Gene in Triticum urartu | Aegilops tauschii | Corresponding gene in Aegilops tauschii | ||
| 1 | f | TRIDC1AG044060 | TtuG1L8-1A | TtG1L8-1A | TRIAE_CS42_1AL_TGACv1_002086_AA0038570 | TaG1L8-1AL | TuG1812G0100003368.01 | TuG1L8 | ||
| 2 | TRIDC1BG049870 | TtuG1L8-1B | TtG1L8-1B | TRIAE_CS42_1BL_TGACv1_030408_AA0089770 | TaG1L8-1BL | |||||
| 3 | TRIAE_CS42_1DL_TGACv1_062292_AA0211930 | TaG1L8-1DL | AET1Gv20705800 | AetG1L8-1D | ||||||
| 4 | c | TRIDC1AG023480 | TtuTAW1-1A | TtTAW1-1A | TRIAE_CS42_1AL_TGACv1_000217_AA0006430 | TaTAW1-1AL | TuG1812G0100001884.01 | TuTAW1 | ||
| 5 | TRIDC1BG028470 | TtuTAW1-1B | TtTAW1-1B | TRIAE_CS42_1BL_TGACv1_033389_AA0139150 | TaTAW1-1BL | |||||
| 6 | TRIAE_CS42_1DL_TGACv1_061798_AA0203560 | TaTAW1-1DL | AET1Gv20402400 | AetTAW1-1D | ||||||
| 7 | h | TRIDC1AG033590 | TtuG1L9-1A | TtG1L9-1A | TRIAE_CS42_1AL_TGACv1_002992_AA0046990 | TaG1L9-1AL | ||||
| 8 | TRIDC1BG038970 | TtuG1L9-1B | TtG1L9-1B | TRIAE_CS42_1BL_TGACv1_031074_AA0107090 | TaG1L9-1BL | |||||
| 9 | TRIAE_CS42_1DL_TGACv1_061913_AA0205560 | TaG1L9-1DL | AET1Gv20554400 | AetG1L9-1D | ||||||
| 10 | a | TRIDC2AG055200 | TtuG1L4-2A | TtG1L4-2A | TRIAE_CS42_2AL_TGACv1_097378_AA0324030 | TaG1L4-2AL | TuG1812G0200004320.01 | TuG1L4 | ||
| 11 | TRIDC2BG057970 | TtuG1L4-2B | TtG1L4-2B | TRIAE_CS42_2BL_TGACv1_129460_AA0384820 | TaG1L4-2BL | |||||
| 12 | TRIAE_CS42_2DL_TGACv1_159566_AA0539920 | TaG1L4-2DL | AET2Gv20853200 | AetG1L4-2D | ||||||
| 13 | l | TRIDC2AG035710 | TtuG1-2A | TtG1-2A | TRIAE_CS42_2AL_TGACv1_092935_AA0267510 | TaG1-2AL | ||||
| 14 | TRIDC2BG039660 | TtuG1-2B | TtG1-2B | TRIAE_CS42_2BL_TGACv1_129607_AA0390240 | TaG1-2BL | |||||
| 15 | TRIAE_CS42_2DL_TGACv1_159706_AA0541770 | TaG1-2DL | AET2Gv20574300 | AetG1-2D | ||||||
| 16 | g | TRIDC3AG050280 | TtuG1L7-3A | TtG1L7-3A | TRIAE_CS42_3AL_TGACv1_193753_AA0619150 | TaG1L7-3AL | ||||
| 17 | TRIDC3BG056500 | TtuG1L7-3B | TtG1L7-3B | TRIAE_CS42_3B_TGACv1_220926_AA0724140 | TaG1L7-3BL | |||||
| 18 | TRIAE_CS42_3DL_TGACv1_249281_AA0844060 | TaG1L7-3DL | AET3Gv20786800 | AetG1L7-3D | ||||||
| 19 | k | TRIDC6AG019090 | TtuG1L1-6A | TtG1L1-6A | TRIAE_CS42_6AS_TGACv1_485429_AA1545270 | TaG1L1-6AS | TuG1812G0600001461.01 | TuG1L1 | ||
| 20 | TRIDC6BG025090 | TtuG1L1-6B | TtG1L1-6B | TRIAE_CS42_6BS_TGACv1_513152_AA1632150 | TaG1L1-6BS | |||||
| 21 | TRIAE_CS42_6DS_TGACv1_542463_AA1721150 | TaG1L1-6DS | AET6Gv20360000 | AetG1L1-6D | ||||||
| 22 | b | TRIDC6AG035110 | TtuG1L3-6A | TtG1L3-6A | TRIAE_CS42_6AL_TGACv1_471514_AA1510200 | TaG1L3-6AL | ||||
| 23 | TRIDC6BG041790 | TtuG1L3-6B | TtG1L3-6B | TRIAE_CS42_6BL_TGACv1_499492_AA1584200 | TaG1L3-6BL | |||||
| 24 | TRIAE_CS42_U_TGACv1_642735_AA2122410 | TaG1L3-6DL | AET6Gv20599400 | AetG1L3-6D | ||||||
| 25 | d | TRIDC6AG056190 | TtuTH1-6A | TtTH1-6A | TRIAE_CS42_6AL_TGACv1_472779_AA1525890 | TaTH1-6AL | TuG1812G0600004039.01 | TuTH1 | ||
| 26 | TRIDC6BG065670 | TtuTH1-6B | TtTH1-6B | TRIAE_CS42_6BL_TGACv1_500779_AA1609580 | TaTH1-6BL | |||||
| 27 | TRIAE_CS42_6DL_TGACv1_528245_AA1712740 | TaTH1-6DL | AET6Gv20927400 | AetTH1-6D | ||||||
| 28 | j | TRIDC7AG065080 | TtuG1L2-7A | TtG1L2-7A | TRIAE_CS42_7AL_TGACv1_556874_AA1772610 | TaG1L2-7AL | TuG1812G0700004994.01 | TuG1L2 | ||
| 29 | TRIDC7BG058610 | TtuG1L2-7B | TtG1L2-7B | TRIAE_CS42_7BL_TGACv1_576798_AA1855280 | TaG1L2-7BL | |||||
| 30 | TRIAE_CS42_7DL_TGACv1_604927_AA2003130 | TaG1L2-7DL | AET7Gv21141300 | AetG1L2-7D |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nan, W.; Shi, S.; Jeewani, D.C.; Quan, L.; Shi, X.; Wang, Z. Genome-Wide Identification and Characterization of wALOG Family Genes Involved in Branch Meristem Development of Branching Head Wheat. Genes 2018, 9, 510. https://doi.org/10.3390/genes9100510
Nan W, Shi S, Jeewani DC, Quan L, Shi X, Wang Z. Genome-Wide Identification and Characterization of wALOG Family Genes Involved in Branch Meristem Development of Branching Head Wheat. Genes. 2018; 9(10):510. https://doi.org/10.3390/genes9100510
Chicago/Turabian StyleNan, Wenzhi, Shandang Shi, Diddugodage Chamila Jeewani, Li Quan, Xue Shi, and Zhonghua Wang. 2018. "Genome-Wide Identification and Characterization of wALOG Family Genes Involved in Branch Meristem Development of Branching Head Wheat" Genes 9, no. 10: 510. https://doi.org/10.3390/genes9100510
APA StyleNan, W., Shi, S., Jeewani, D. C., Quan, L., Shi, X., & Wang, Z. (2018). Genome-Wide Identification and Characterization of wALOG Family Genes Involved in Branch Meristem Development of Branching Head Wheat. Genes, 9(10), 510. https://doi.org/10.3390/genes9100510