Genetic Polymorphisms and In Silico Mutagenesis Analyses of CYP2C9, CYP2D6, and CYPOR Genes in the Pakistani Population


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants, DNA Extraction and Quantification
2.2. Polymerase Chain Reaction and Sequence Analysis
2.3. Statistical Analysis
2.4. In Silico Mutagenesis Analysis: Modeling of CYP2C9, CYP2D6, and CYPOR Variant Protein Structures
3. Results and Discussion
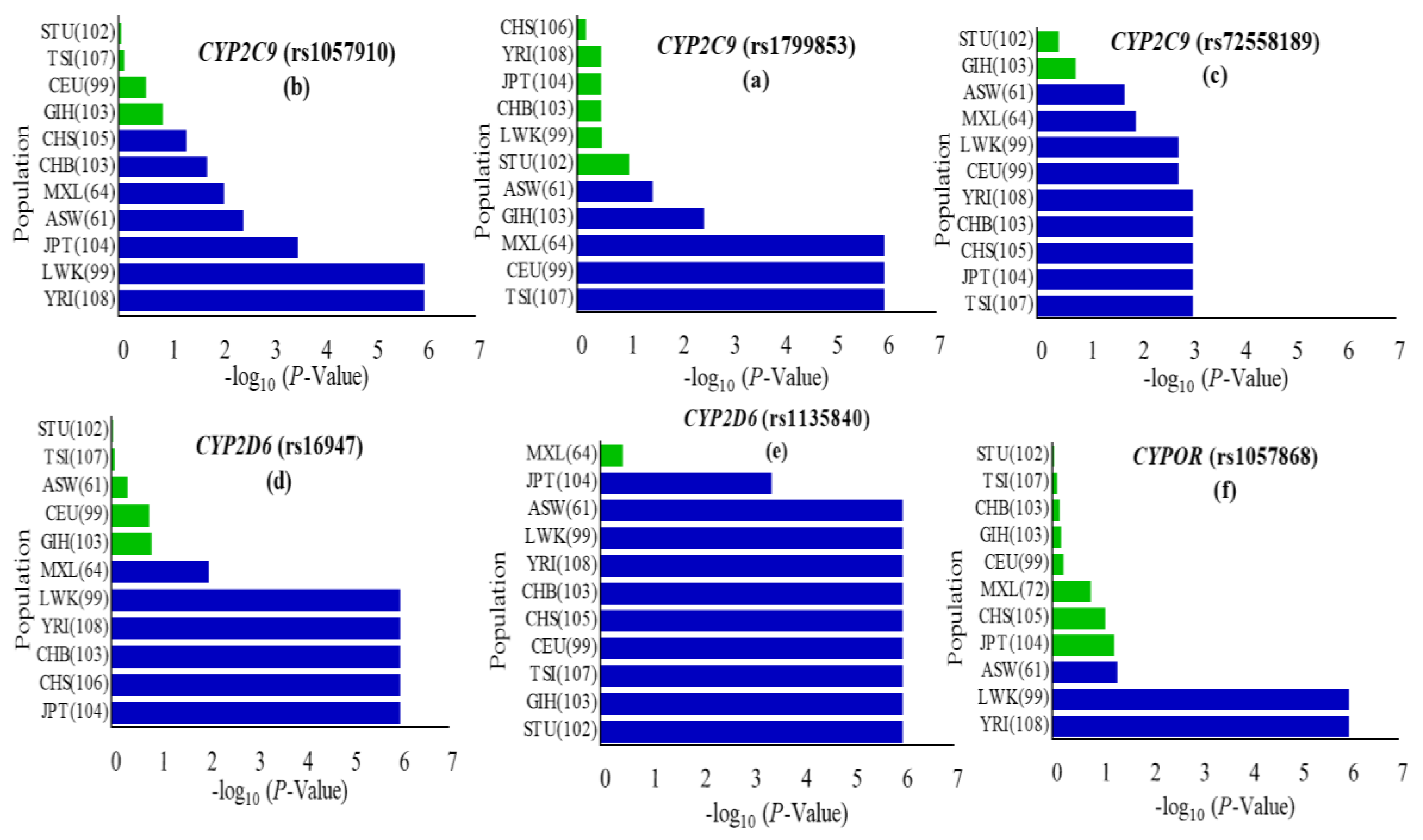
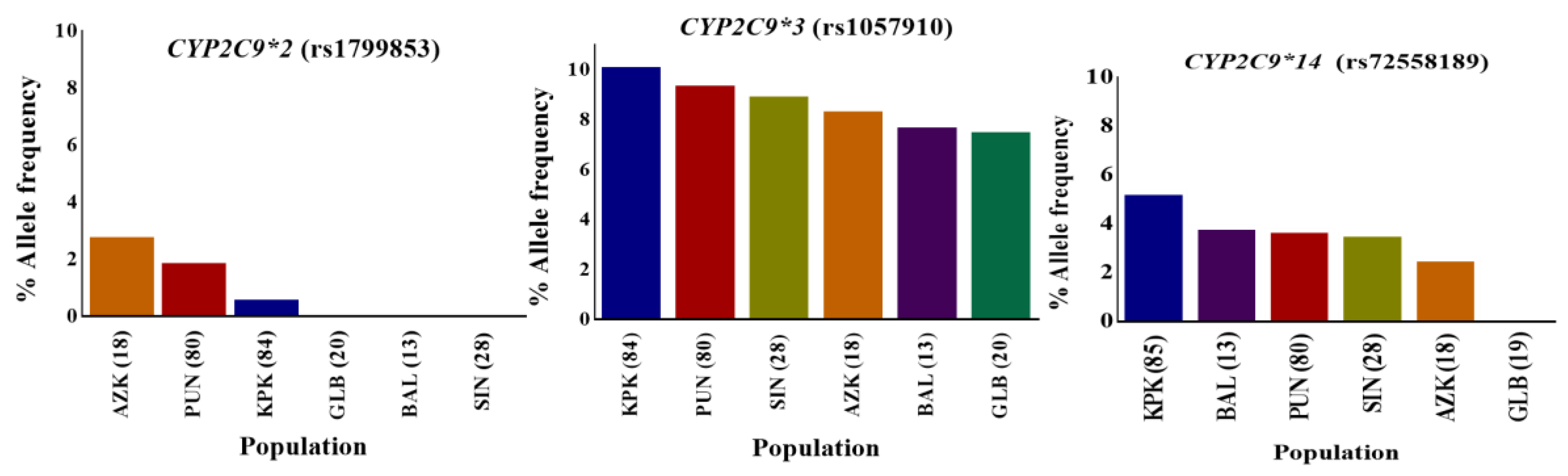
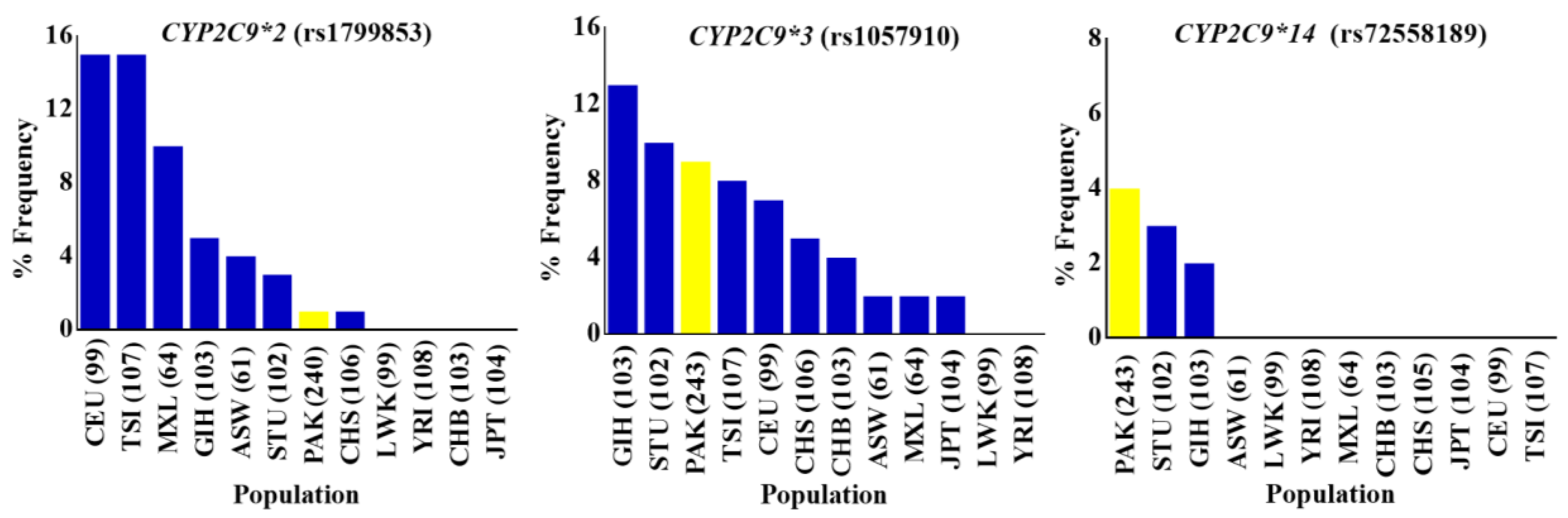
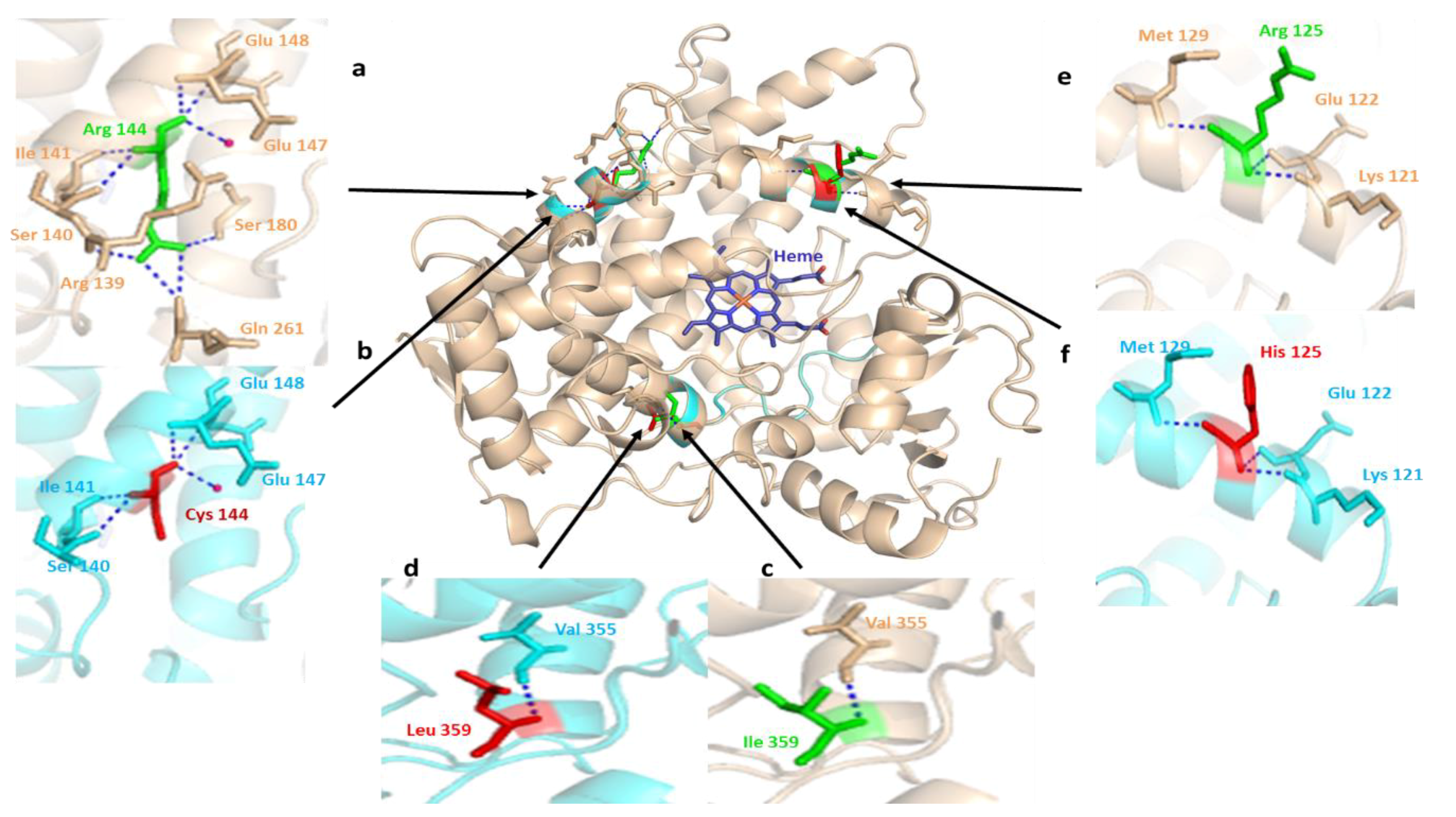
3.1. CYP2C9 Polymorphisms
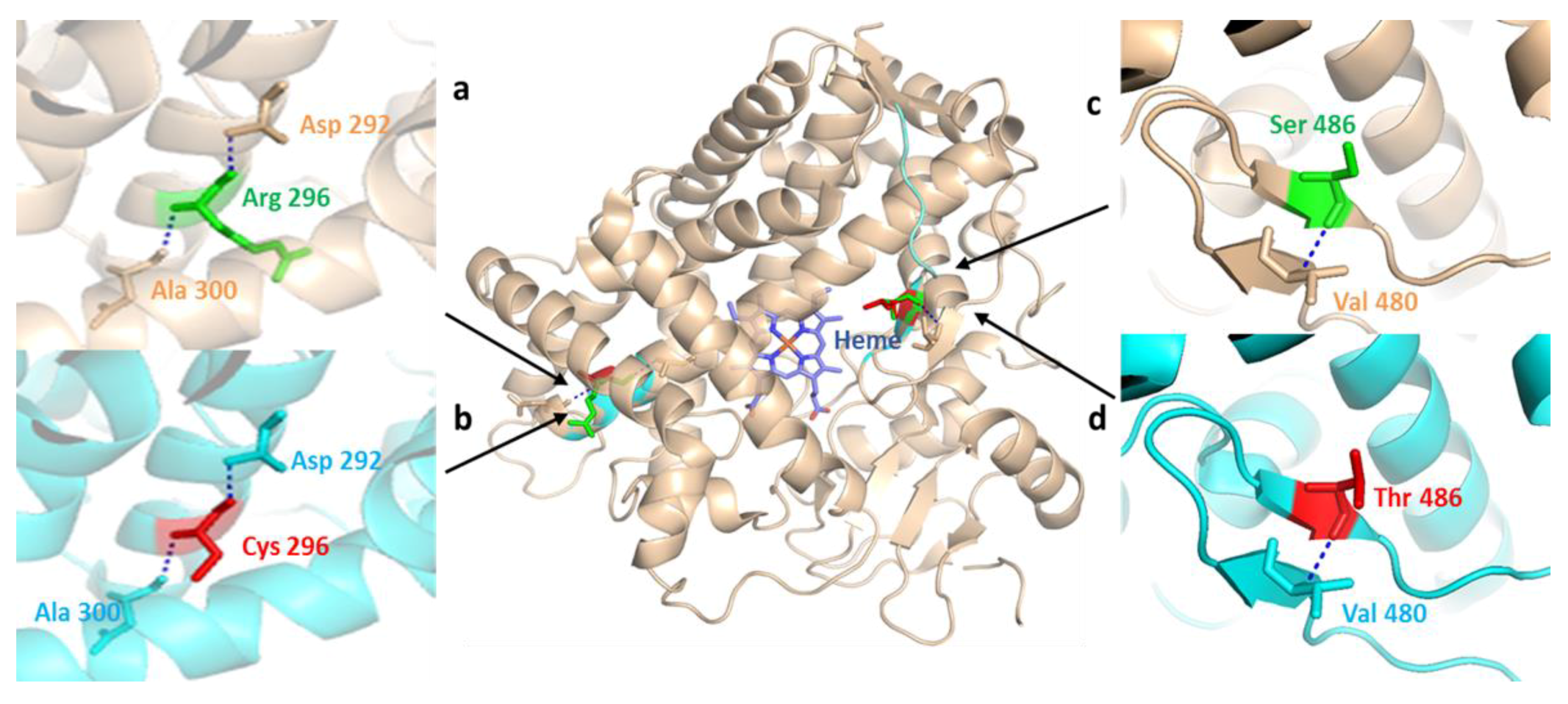
3.2. CYP2D6 Polymorphisms
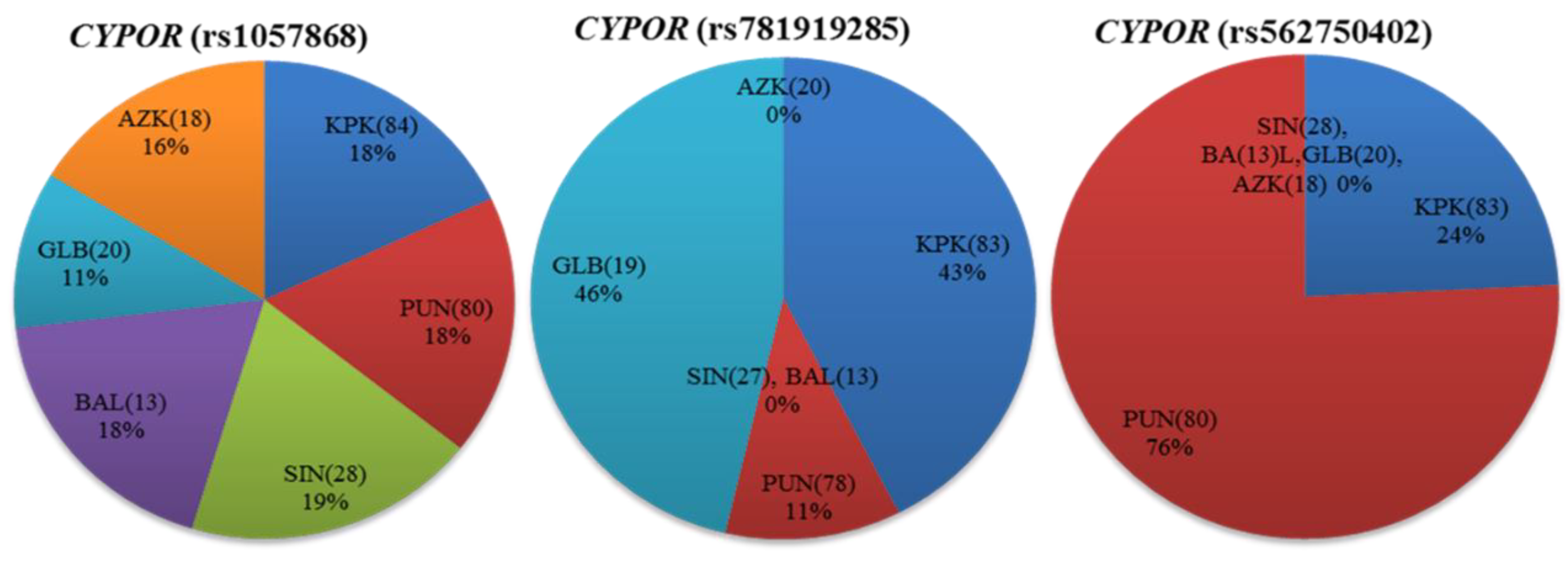
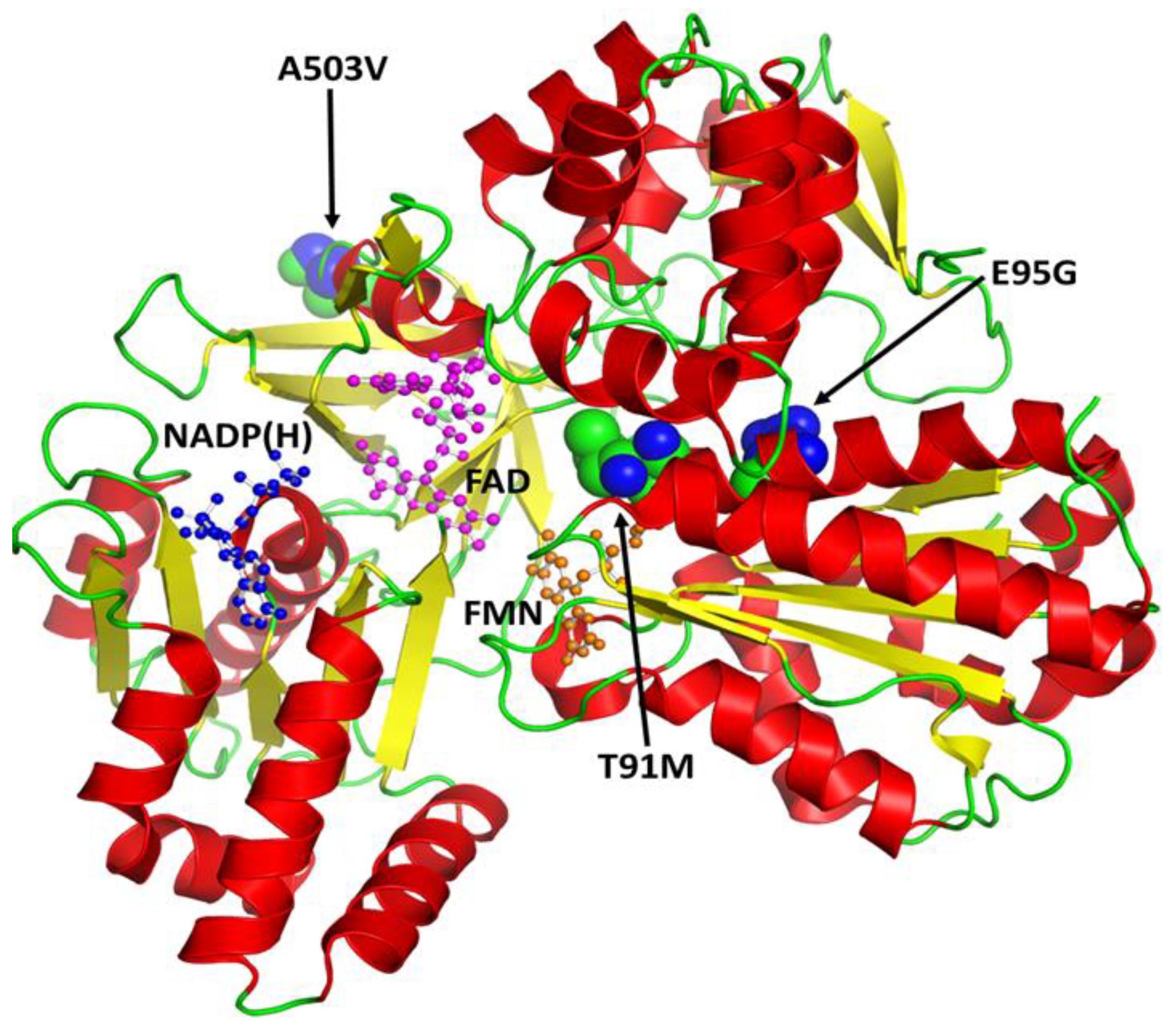
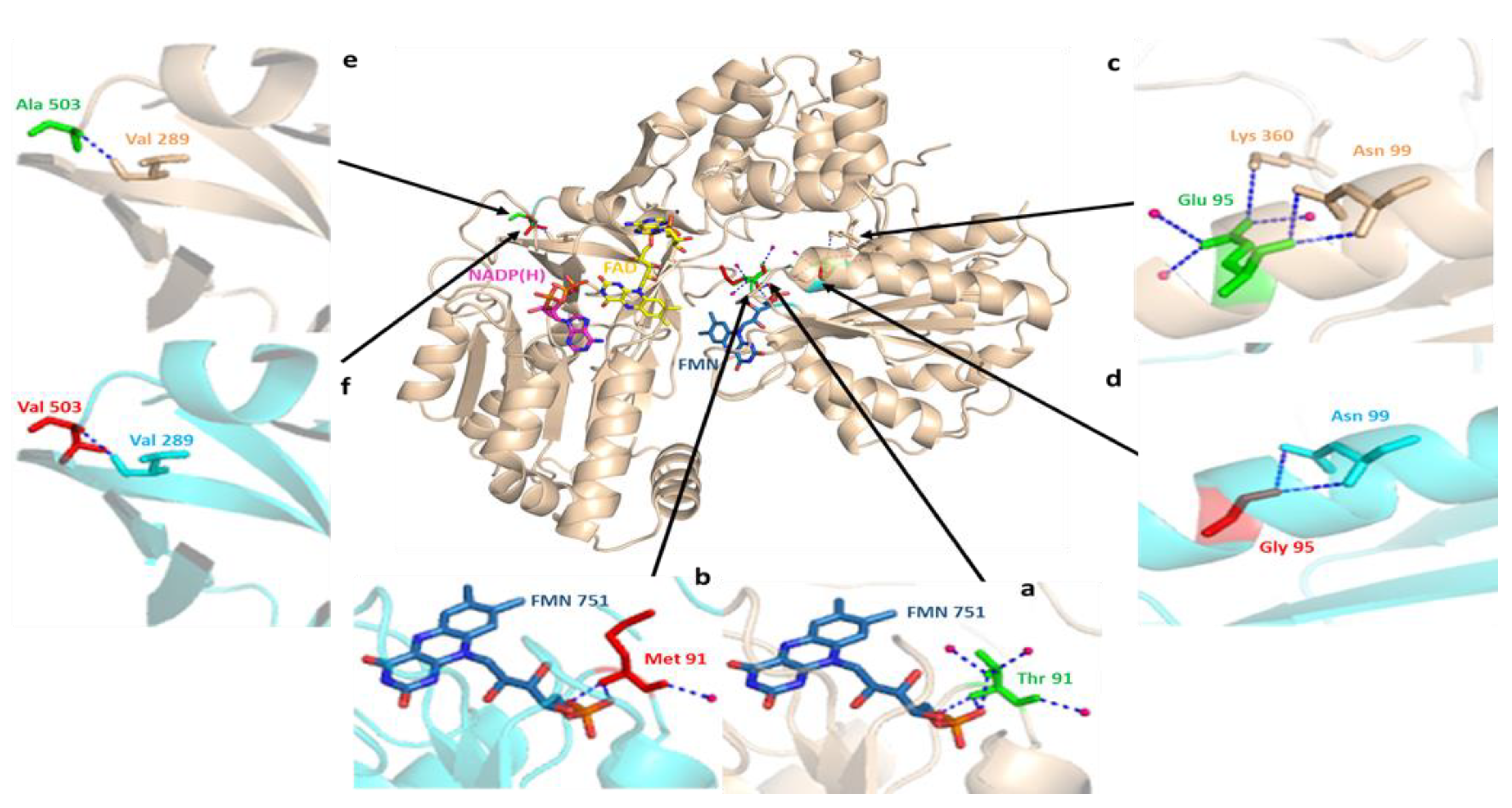
3.3. CYPOR Polymorphisms
3.4. In Silico Influences of SNPs on the Structure and Functions of CYP2C9, CYP2D6, and CYPOR Proteins
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bernard, S.; Neville, K.A.; Nguyen, A.T.; Flockhart, D.A. Interethnic differences in genetic polymorphisms of CYP2D6 in the U.S. Population: Clinical implications. Oncologist 2006, 11, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.K.; Woo, M.H.; Mcleod, H.L. Genetic basis of drug metabolism. Am. J. Health-Syst. Pharm. 2002, 59, 2061–2069. [Google Scholar] [PubMed]
- Zhang, H.F.; Li, Z.H.; Liu, J.Y.; Liu, T.T.; Wang, P.; Fang, Y.; Zhou, J.; Cui, M.Z.; Gao, N.; Tian, X.; et al. Correlation of cytochrome P450 oxidoreductase expression with the expression of 10 isoforms of cytochrome P450 in human liver. Drug Metab. Dispos. 2016, 44, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, S.J.; Begg, E.J. Pharmacogenetics, drug-metabolizing enzymes, and clinical practice. Pharmacol. Rev. 2006, 58, 521–590. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.R.; Goldstein, J.A.; Pieper, J.A. Cytochrome P450 2C9 polymorphisms: A comprehensive review of the in-vitro and human data. Pharmacogenetics 2002, 12, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.A.; Faletto, M.B.; Romkes-Sparks, M.; Sullivan, T.; Kitareewan, S.; Raucy, J.L.; Lasker, J.M.; Ghanayem, B.I. Evidence that CYP2C19 is the major (S)-mephenytoin 4’-hydroxylase in humans. Biochemistry 1994, 33, 1743–1752. [Google Scholar] [CrossRef] [PubMed]
- PharmVar, Pharmacogene Variation Consortium, The Human Cytochrome P450 (CYP) Allele Nomenclature Database. Available online: https://www.pharmvar.org/htdocs/archive/cyp2c9.htm (accessed on 23 March 2018).
- Zhao, F.; Loke, C.; Rankin, S.C.; Guo, J.Y.; Lee, H.S.; Wu, T.S.; Tan, T.; Liu, T.C.; Lu, W.L.; Lim, Y.T.; et al. Novel CYP2C9 genetic variants in Asian subjects and their influence on maintenance warfarin dose. Clin. Pharmacol. Ther. 2004, 76, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Crespi, C.L.; Miller, V.P. The r144c change in the CYP2C9*2 allele alters interaction of the cytochrome p450 with NADPH: Cytochrome P450 oxidoreductase. Pharmacogenetics 1997, 7, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Sullivan-Klose, T.H.; Ghanayem, B.I.; Bell, D.A.; Zhang, Z.Y.; Kaminsky, L.S.; Shenfield, G.M.; Miners, J.O.; Birkett, D.J.; Goldstein, J.A. The role of the CYP2C9-LEU359 allelic variant in the tolbutamide polymorphism. Pharmacogenetics 1996, 6, 341–349. [Google Scholar] [CrossRef] [PubMed]
- DeLozier, T.C.; Lee, S.C.; Coulter, S.J.; Goh, B.C.; Goldstein, J.A. Functional characterization of novel allelic variants of CYP2C9 recently discovered in southeast Asians. J. Pharmacol. Exp. Ther. 2005, 315, 1085–1090. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R.; Koymans, L.; Kamataki, T.; Stegeman, J.J.; Feyereisen, R.; Waxman, D.J.; Waterman, M.R.; Gotoh, O.; Coon, M.J.; Estabrook, R.W.; et al. P450 superfamily: Update on new sequences, gene mapping, accession numbers and nomenclature. Pharmacogenetics 1996, 6, 1–42. [Google Scholar] [CrossRef] [PubMed]
- Ingelman-Sundberg, M.; Evans, W.E. Unravelling the functional genomics of the human CYP2D6 gene locus. Pharmacogenetics 2001, 11, 553–554. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.F. Polymorphism of human cytochrome P450 2D6 and its clinical significance: Part I. Clin. Pharmacokinet. 2009, 48, 689–723. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.F. Polymorphism of human cytochrome P450 2D6 and its clinical significance: Part ii. Clin. Pharmacokinet. 2009, 48, 761–804. [Google Scholar] [CrossRef] [PubMed]
- PharmVar, Pharmacogene Variation Consortium, The Human Cytochrome P450 (CYP) Allele Nomenclature Database. Available online: https://www.pharmvar.org/htdocs/archive/cyp2d6.htm (accessed on 27 March 2018).
- Kitada, M. Genetic polymorphism of cytochrome P450 enzymes in Asian populations: Focus on CYP2D6. Int. J. Clin. Pharmacol. Res. 2003, 23, 31–35. [Google Scholar] [PubMed]
- Ahmed, S.; Zhou, Z.; Zhou, J.; Chen, S.Q. Pharmacogenomics of drug metabolizing enzymes and transporters: Relevance to precision medicine. Genom. Proteom. Bioinform. 2016, 14, 298–313. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.V.; Fluck, C.E. Nadph P450 oxidoreductase: Structure, function, and pathology of diseases. Pharmacol. Ther. 2013, 138, 229–254. [Google Scholar] [CrossRef] [PubMed]
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, S.C.; Miller, W.L.; Zhong, X.B.; Arlt, W.; Ogata, T.; Ding, X.; Wolf, C.R.; Fluck, C.E.; Pandey, A.V.; Henderson, C.J.; et al. Nomenclature for alleles of the cytochrome P450 oxidoreductase gene. Pharmacogenet. Genom. 2009, 19, 565–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- PharmVar, Pharmacogene Variation Consortium, The Human Cytochrome P450 (CYP) Allele Nomenclature Database. Available online: https://www.pharmvar.org/htdocs/archive/por.htm (accessed on 30 March 2018).
- Dhir, V.; Ivison, H.E.; Krone, N.; Shackleton, C.H.; Doherty, A.J.; Stewart, P.M.; Arlt, W. Differential inhibition of CYP17A1 and CYP21A2 activities by the P450 oxidoreductase mutant a287p. Mol. Endocrinol. 2007, 21, 1958–1968. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.V.; Kempna, P.; Hofer, G.; Mullis, P.E.; Fluck, C.E. Modulation of human CYP19A1 activity by mutant NADPH P450 oxidoreductase. Mol. Endocrinol. 2007, 21, 2579–2595. [Google Scholar] [CrossRef] [PubMed]
- NIH. Genome Reference Consortium, The International Genome Sample Resource. Available online: http://www.1000genomes.org/ (accessed on 28 April 2018).
- Rajput, T.A.; Naveed, A.K.; Khan, S.; Farooqi, Z.U. Frequencies of two functionally significant SNPs and their haplotypes of organic anion transporting polypeptide 1b1 slco1b1 gene in six ethnic groups of Pakistani population. Iran. J. Basic Med. Sci. 2014, 17, 441–447. [Google Scholar] [PubMed]
- Rodriguez-Flores, J.L.; Fuller, J.; Hackett, N.R.; Salit, J.; Malek, J.A.; Al-Dous, E.; Chouchane, L.; Zirie, M.; Jayoussi, A.; Mahmoud, M.A.; et al. Exome sequencing of only seven qataris identifies potentially deleterious variants in the qatari population. PLoS ONE 2012, 7, e47614. [Google Scholar] [CrossRef] [PubMed]
- NIH. Primer3 (v. 0.4.0) Pick Primers from a DNA sequence. Available online: http://bioinfo.ut.ee/primer3-0.4.0/primer3/ (accessed on 15 May 2017).
- Ratajewski, M.; Slomka, M.; Karas, K.; Sobalska-Kwapis, M.; Korycka-Machala, M.; Salkowska, A.; Dziadek, J.; Strapagiel, D.; Dastych, J. Functional analysis of the rs774872314, rs116171003, rs200231898 and rs201107751 polymorphisms in the human RORγt gene promoter region. Genes 2017, 8, 126. [Google Scholar] [CrossRef] [PubMed]
- Pearson, W.R. Finding protein and nucleotide similarities with fasta. Curr. Protoc. Bioinform. 2016, 53, 3–9. [Google Scholar]
- Madden, T. The NCBI Handbook Internet, 2nd ed.; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2013. [Google Scholar]
- NCBI Resource Coordinators. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2015, 43, D6–D17. [Google Scholar] [CrossRef] [PubMed]
- Rooryck, C.; Roudaut, C.; Robine, E.; Musebeck, J.; Arveiler, B. Oculocutaneous albinism with tyrp1 gene mutations in a caucasian patient. Pigment Cell Res. 2006, 19, 239–242. [Google Scholar] [CrossRef] [PubMed]
- OEGE. Online Encyclopedia for Genetic Epidemiology Studies, Chi-Square Test for Deviation from Hardy-Weinberg Equilibrium. Available online: http://www.genes.org.uk/software/hardy-weinberg.html (accessed on 25 April 2018).
- R Development Core Team. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org (accessed on 20 January 2018).
- RCSB. Protein Data Bank, A Structural View of Biology. Available online: https://www.rcsb.org/ (accessed on 25 April 2017).
- Maekawa, K.; Adachi, M.; Matsuzawa, Y.; Zhang, Q.; Kuroki, R.; Saito, Y.; Shah, M.B. Structural basis of single-nucleotide polymorphisms in cytochrome P450 2c9. Biochemistry 2017, 56, 5476–5480. [Google Scholar] [CrossRef] [PubMed]
- Sundermann, A.; Oostenbrink, C. Molecular dynamics simulations give insight into the conformational change, complex formation, and electron transfer pathway for cytochrome p450 reductase. Protein Sci. 2013, 22, 1183–1195. [Google Scholar] [CrossRef] [PubMed]
- Fluck, C.E.; Mullis, P.E.; Pandey, A.V. Modeling of human P450 oxidoreductase structure by in silico mutagenesis and md simulation. Mol. Cell. Endocrinol. 2009, 313, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Saunders, M.A.; Hammer, M.F.; Nachman, M.W. Nucleotide variability at G6PD and the signature of malarial selection in humans. Genetics 2002, 162, 1849–1861. [Google Scholar] [PubMed]
- Hamblin, M.T.; Thompson, E.E.; Di Rienzo, A. Complex signatures of natural selection at the duffy blood group locus. Am. J. Hum. Genet. 2002, 70, 369–383. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, S.; Aslamkhan, M.; Attimonelli, M.; Abbas, S.; Aydin, H.H. Mitochondrial DNA variation in the Sindh population of Pakistan. Aust. J. Forensic Sci. 2017, 49, 201–216. [Google Scholar] [CrossRef]
- Rosenberg, N.A.; Pritchard, J.K.; Weber, J.L.; Cann, H.M.; Kidd, K.K.; Zhivotovsky, L.A.; Feldman, M.W. Genetic structure of human populations. Science 2002, 298, 2381–2385. [Google Scholar] [CrossRef] [PubMed]
- De Morais, S.M.; Wilkinson, G.R.; Blaisdell, J.; Nakamura, K.; Meyer, U.A.; Goldstein, J.A. The major genetic defect responsible for the polymorphism of s-mephenytoin metabolism in humans. J. Biol. Chem. 1994, 269, 15419–15422. [Google Scholar] [PubMed]
- Dai, D.P.; Xu, R.A.; Hu, L.M.; Wang, S.H.; Geng, P.W.; Yang, J.F.; Yang, L.P.; Qian, J.C.; Wang, Z.S.; Zhu, G.H.; et al. Cyp2c9 polymorphism analysis in Han Chinese populations: Building the largest allele frequency database. Pharmacogenom. J. 2014, 14, 85–92. [Google Scholar] [CrossRef] [PubMed]
- London, S.J.; Daly, A.K.; Leathart, J.B.; Navidi, W.C.; Idle, J.R. Lung cancer risk in relation to the cyp2c9*1/cyp2c9*2 genetic polymorphism among African-Americans and Caucasians in Los Angeles County, California. Pharmacogenetics 1996, 6, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.C.; Xu, X.M.; Hu, G.X.; Dai, D.P.; Xu, R.A.; Hu, L.M.; Li, F.H.; Zhang, X.H.; Yang, J.F.; Cai, J.P. Genetic variations of human CYP2D6 in the Chinese Han population. Pharmacogenomics 2013, 14, 1731–1743. [Google Scholar] [CrossRef] [PubMed]
- Johansson, I.; Lundqvist, E.; Bertilsson, L.; Dahl, M.L.; Sjoqvist, F.; Ingelman-Sundberg, M. Inherited amplification of an active gene in the cytochrome P450 CYP2D locus as a cause of ultrarapid metabolism of debrisoquine. Proc. Natl. Acad. Sci. USA 1993, 90, 11825–11829. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Kang, C.; Yuan, J.; Zhang, Y.; Wei, Y.; Xu, L.; Zhou, F.; Yang, J. CYP2D6 polymorphisms are associated with effects of risperidone on neurocognitive performance in schizophrenia. Schizophr. Res. 2017, 188, 50–51. [Google Scholar] [CrossRef] [PubMed]
- Sakuyama, K.; Sasaki, T.; Ujiie, S.; Obata, K.; Mizugaki, M.; Ishikawa, M.; Hiratsuka, M. Functional characterization of 17 CYP2D6 allelic variants (CYP2D6.2, 10, 14a-b, 18, 27, 36, 39, 47–51, 53–55, and 57). Drug Metab. Dispos. 2008, 36, 2460–2467. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.P.; Luan, H.; Dong, X.H.; Jin, G.J.; Man, D.L.; Shang, H. Genetic variants of CYP2D6 gene and cancer risk: A huge systematic review and meta-analysis. Asian Pac. J. Cancer Prev. 2012, 13, 3165–3172. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Han, J.L.; Mi, Y.C.; Xiao, Z.J.; Feng, S.Z.; Zhou, Y.L.; Wang, J.X.; Han, M.Z. Relationships between the gene polymorphisms of drug metabolizing enzymes and the outcome of the first induction chemotherapy in patients with de novo acute myeloid leukemia. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2011, 19, 327–331. [Google Scholar] [PubMed]
- Yuan, H.; Yu, M.; Yang, Y.; Wu, K.; Lin, X.; Li, J. Association of CYP2D6 single-nucleotide polymorphism with response to ophthalmic timolol in primary open-angle glaucoma—A pilot study. J. Ocul. Pharmacol. Ther. 2010, 26, 497–501. [Google Scholar] [CrossRef] [PubMed]
- McCammon, K.M.; Panda, S.P.; Xia, C.; Kim, J.J.; Moutinho, D.; Kranendonk, M.; Auchus, R.J.; Lafer, E.M.; Ghosh, D.; Martasek, P.; et al. Instability of the human cytochrome P450 reductase A287P variant is the major contributor to its Antley-Bixler syndrome-like phenotype. J. Biol. Chem. 2016, 291, 20487–20502. [Google Scholar] [CrossRef] [PubMed]
- Koika, V.; Armeni, A.K.; Georgopoulos, N.A. Delayed diagnosis of disorder of sex development (DSD) due to P450 oxidoreductase (POR) deficiency. Hormones 2016, 15, 277–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marohnic, C.C.; Panda, S.P.; Martasek, P.; Masters, B.S. Diminished FAD binding in the Y459H and V492E antley-bixler syndrome mutants of human cytochrome p450 reductase. J. Biol. Chem. 2006, 281, 35975–35982. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, V.; Choi, J.H.; Giacomini, K.M.; Miller, W.L. Substrate-specific modulation of CYP3A4 activity by genetic variants of cytochrome P450 oxidoreductase. Pharmacogenet. Genom. 2010, 20, 611–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elens, L.; Nieuweboer, A.J.; Clarke, S.J.; Charles, K.A.; de Graan, A.J.; Haufroid, V.; van Gelder, T.; Mathijssen, R.H.; van Schaik, R.H. Impact of POR*28 on the clinical pharmacokinetics of CYP3A phenotyping probes midazolam and erythromycin. Pharmacogenet. Genom. 2013, 23, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Chenoweth, M.J.; Zhu, A.Z.; Sanderson Cox, L.; Ahluwalia, J.S.; Benowitz, N.L.; Tyndale, R.F. Variation in p450 oxidoreductase (POR) A503V and flavin-containing monooxygenase (FMO)-3 E158K is associated with minor alterations in nicotine metabolism, but does not alter cigarette consumption. Pharmacogenet. Genom. 2014, 24, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Kuypers, D.R.; de Loor, H.; Naesens, M.; Coopmans, T.; de Jonge, H. Combined effects of CYP3A5*1, POR*28, and CYP3A4*22 single nucleotide polymorphisms on early concentration-controlled tacrolimus exposure in de-novo renal recipients. Pharmacogenet. Genom. 2014, 24, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Woillard, J.B.; Kamar, N.; Coste, S.; Rostaing, L.; Marquet, P.; Picard, N. Effect of CYP3A4*22, POR*28, and PPARA rs4253728 on sirolimus in vitro metabolism and trough concentrations in kidney transplant recipients. Clin. Chem. 2013, 59, 1761–1769. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Ma, G.; Li, S.; Wang, M.; Liu, N.; Ma, L.; Zhang, Z.; Chu, H.; Zhang, Z.; Wang, S.L. Functional por a503v is associated with the risk of bladder cancer in a Chinese population. Sci. Rep. 2015, 5, 11751. [Google Scholar] [CrossRef] [PubMed]
- Nair, P.C.; McKinnon, R.A.; Miners, J.O. Cytochrome P450 structure-function: Insights from molecular dynamics simulations. Drug Metab. Rev. 2016, 48, 434–452. [Google Scholar] [CrossRef] [PubMed]
- Isvoran, A.; Louet, M.; Vladoiu, D.L.; Craciun, D.; Loriot, M.A.; Villoutreix, B.O.; Miteva, M.A. Pharmacogenomics of the cytochrome P450 2c family: Impacts of amino acid variations on drug metabolism. Drug Discov. Today 2017, 22, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Nzabonimpa, G.S.; Rasmussen, H.B.; Brunak, S.; Taboureau, O. Investigating the impact of missense mutations in hCES1 by in silico structure-based approaches. Drug Metab. Pers. Ther. 2016. [Google Scholar] [CrossRef] [PubMed]
- Khalil, S.I.; Mahmoud, T.A.A.A.; Mohammed, W.S.A.; Elbager, S.G.; Elmoselhy, A.M.; Alfaki, M.A.I. In silico genetic variation analysis of cytochrome P450 2c19 and their effect in certain drugs metabolism. Am. J. Bioinform. Res. 2017, 7, 59–65. [Google Scholar]
- Nishita, N.; Suvagiya, P.; Georrge, J.J. Modeling mutations, docking, primer and probe designing of cytochrome P450 2d6, a drug metabolizing enzyme. In Proceedings of the 9th National Level Science Symposium; Christ Publications: Rajkot, India, 2016; Volume 3, pp. 328–333. [Google Scholar]
- Blaisdell, J.; Jorge-Nebert, L.F.; Coulter, S.; Ferguson, S.S.; Lee, S.J.; Chanas, B.; Xi, T.; Mohrenweiser, H.; Ghanayem, B.; Goldstein, J.A. Discovery of new potentially defective alleles of human CYP2C9. Pharmacogenetics 2004, 14, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Sano, E.; Li, W.; Yuki, H.; Liu, X.; Furihata, T.; Kobayashi, K.; Chiba, K.; Neya, S.; Hoshino, T. Mechanism of the decrease in catalytic activity of human cytochrome P450 2c9 polymorphic variants investigated by computational analysis. J. Comput. Chem. 2010, 31, 2746–2758. [Google Scholar] [CrossRef] [PubMed]
- Takanashi, K.; Tainaka, H.; Kobayashi, K.; Yasumori, T.; Hosakawa, M.; Chiba, K. CYP2C9 Ile359 and Leu359 variants: Enzyme kinetic study with seven substrates. Pharmacogenetics 2000, 10, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Bridges, A.; Gruenke, L.; Chang, Y.T.; Vakser, I.A.; Loew, G.; Waskell, L. Identification of the binding site on cytochrome p450 2b4 for cytochrome b5 and cytochrome p450 reductase. J. Biol. Chem. 1998, 273, 17036–17049. [Google Scholar] [CrossRef] [PubMed]
- de Waal, P.W.; Sunden, K.F.; Furge, L.L. Molecular dynamics of CYP2D6 polymorphisms in the absence and presence of a mechanism-based inactivator reveals changes in local flexibility and dominant substrate access channels. PLoS ONE 2014, 9, e108607. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lim, Y.R.; Han, S.; Han, J.S.; Chun, Y.J.; Yun, C.H.; Lee, C.H.; Kim, D. Functional influence of human CYP2D6 allelic variations: P34s, e418k, s486t, and r296c. Arch. Pharm. Res. 2013, 36, 1500–1506. [Google Scholar] [CrossRef] [PubMed]
- Martiny, V.Y.; Miteva, M.A. Advances in molecular modeling of human cytochrome P450 polymorphism. J. Mol. Biol. 2013, 425, 3978–3992. [Google Scholar] [CrossRef] [PubMed]
- Niinuma, Y.; Saito, T.; Takahashi, M.; Tsukada, C.; Ito, M.; Hirasawa, N.; Hiratsuka, M. Functional characterization of 32 CYP2C9 allelic variants. Pharmacogenom. J. 2014, 14, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Ciccacci, C.; Falconi, M.; Paolillo, N.; Oteri, F.; Forte, V.; Novelli, G.; Desideri, A.; Borgiani, P. Characterization of a novel CYP2C9 gene mutation and structural bioinformatic protein analysis in a warfarin hypersensitive patient. Pharmacogenet. Genom. 2011, 21, 344–346. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Pandey, A.V.; Agrawal, V.; Reardon, W.; Lapunzina, P.D.; Mowat, D.; Jabs, E.W.; Van Vliet, G.; Sack, J.; Fluck, C.E.; et al. Diversity and function of mutations in P450 oxidoreductase in patients with antley-bixler syndrome and disordered steroidogenesis. Am. J. Hum. Genet. 2005, 76, 729–749. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Roberts, D.L.; Paschke, R.; Shea, T.M.; Masters, B.S.; Kim, J.J. Three-dimensional structure of NADPH-cytochrome P450 reductase: Prototype for FMN- and FAD-containing enzymes. Proc. Natl. Acad. Sci. USA 1997, 94, 8411–8416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandee, D.; Morrissey, K.; Agrawal, V.; Tam, H.K.; Kramer, M.A.; Tracy, T.S.; Giacomini, K.M.; Miller, W.L. Effects of genetic variants of human P450 oxidoreductase on catalysis by CYP2D6 in vitro. Pharmacogenet. Genom. 2010, 20, 677–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene. | SNP ID | Amino Acid Substitution | a Chromosomal Position | b SNP Function | c MAF (Pak) |
|---|---|---|---|---|---|
| CYP2C9 | rs1799853 | R144C | chr10:94942290 | missense | 0.0479 (0.0104) |
| CYP2C9 | rs1057910 | I359L | chr10:94981296 | missense | 0.0485 (0.0926 |
| CYP2C9 | rs72558189 | R125H | chr10:94942234 | missense | 0.0042 (0.0412) |
| CYP2D6 | rs16947 | R296C | chr22:42127941 | missense | 0.3592 (0.3817) |
| CYP2D6 | rs1135840 | S486T | chr22:42126611 | missense | 0.4012 (0.3521) |
| CYP2D6 | rs566383351 | chr22:42131222 | intron variant | 0.0004 (0.1459) | |
| CYP2D6 | rs530422334 | chr22:42131145 | intron variant | 0.0002 (0.0207) | |
| CYP2D6 | rs35023634 | chr22:42131023 | intron variant | 0.0004 (0.0014) | |
| CYP2D6 | rs35481113 | chr22:42131016 | intron variant | 0.0006 (0.0166) | |
| CYP2D6 | rs543939200 | chr22:42130994 | intron variant | 0.0002 (0) | |
| CYP2D6 | rs372204775 | chr22:42130993 | intron variant | 0.0084 (0.09) | |
| CYP2D6 | rs267608272 | chr22:42130929 | intron variant | 0.0022 (0) | |
| CYP2D6 | rs28371725 | chr22:42127803 | intron variant | 0.0635 (0.1958) | |
| CYP2D6 | rs28371738 | chr22:42126390 | intron variant | 0.2368 (0.1033) | |
| CYP2D6 | rs374672076 | chr22:42131189 | intron variant | NA (0.127) | |
| CYP2D6 | rs17002852 | H232H | chr22:42128321 | synonymous | 0.0072 (0.0271) |
| CYP2D6 | rs34167214 | chr22:42131122 | intron variant | NA (0.5) | |
| CYP2D6 | rs35046171 | chr22:42131063 | intron variant | NA (0.1339) | |
| CYPOR | rs1057868 | A503V | chr7:75985688 | missense | 0.2861 (0.3251) |
| CYPOR | rs1057870 | S572S | chr7:75985969 | synonymous | 0.1953 (0.2851) |
| CYPOR | rs781919285 | E95G | chr7:75979497 | missense | 0.000009 (0.0042) |
| CYPOR | rs562750402 | T91M | chr7:75979485 | missense | 0.00003 (0.0062) |
| CYPOR | rs71526806 | chr7:75985545 | intron variant | NA (0.095) | |
| CYPOR | rs4732514 | chr7:75984680 | intron variant | 0.4952 (0.5729) | |
| CYPOR | rs4732515 | chr7:75984711 | intron variant | 0.1881 (0.0612) | |
| CYPOR | rs4732516 | chr7:75984764 | intron variant | 0.1881 (0.0599) | |
| CYPOR | rs2228104 | A485A | chr7:75985635 | synonymous | 0.1947 (0.0702) |
| CYPOR | rs2302433 | chr7:75985882 | intron variant | 0.0246 (0.0166) | |
| CYPOR | rs1135612 | P129P | chr7:75980359 | synonymous | 0.2494 (0.2303) |
| CYPOR | rs13223707 | chr7:75983452 | intron variant | 0.1887 (0.0622) | |
| CYPOR | rs13240147 | chr7:75983465 | intron variant | 0.1889 (0.1889) | |
| CYPOR | rs41301394 | chr7:75983485 | intron variant | 0.275 (0.3229) | |
| CYPOR | rs10239977 | chr7:75979668 | intron variant | 0.1963 (0.2063) | |
| CYPOR | rs533975609 | chr7:75979781 | intron variant | 0.0016 (0.0232) | |
| CYPOR | rs72554000 | chr7:75979826 | intron variant | 0.0106 (0.0167) | |
| CYPOR | rs369026313 | D163D | chr7:75980461 | synonymous | 0.00003 (0) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, S.; Zhou, J.; Zhou, Z.; Chen, S.-Q. Genetic Polymorphisms and In Silico Mutagenesis Analyses of CYP2C9, CYP2D6, and CYPOR Genes in the Pakistani Population. Genes 2018, 9, 514. https://doi.org/10.3390/genes9100514
Ahmed S, Zhou J, Zhou Z, Chen S-Q. Genetic Polymorphisms and In Silico Mutagenesis Analyses of CYP2C9, CYP2D6, and CYPOR Genes in the Pakistani Population. Genes. 2018; 9(10):514. https://doi.org/10.3390/genes9100514
Chicago/Turabian StyleAhmed, Shabbir, Jie Zhou, Zhan Zhou, and Shu-Qing Chen. 2018. "Genetic Polymorphisms and In Silico Mutagenesis Analyses of CYP2C9, CYP2D6, and CYPOR Genes in the Pakistani Population" Genes 9, no. 10: 514. https://doi.org/10.3390/genes9100514
APA StyleAhmed, S., Zhou, J., Zhou, Z., & Chen, S. -Q. (2018). Genetic Polymorphisms and In Silico Mutagenesis Analyses of CYP2C9, CYP2D6, and CYPOR Genes in the Pakistani Population. Genes, 9(10), 514. https://doi.org/10.3390/genes9100514