Molecular Characterization of Annexin B2, B3 and B12 in Taenia multiceps

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Parasites and Animals
2.3. Bioinformatics Analysis
2.4. Cloning, Expression and Purification of Recombinant TmAnxB2, TmAnxB3 and TmAnxB12
2.5. Western Blotting
2.6. Fluorescence Immunohistochemistry
2.7. Quantitative Real-Time PCR
3. Results
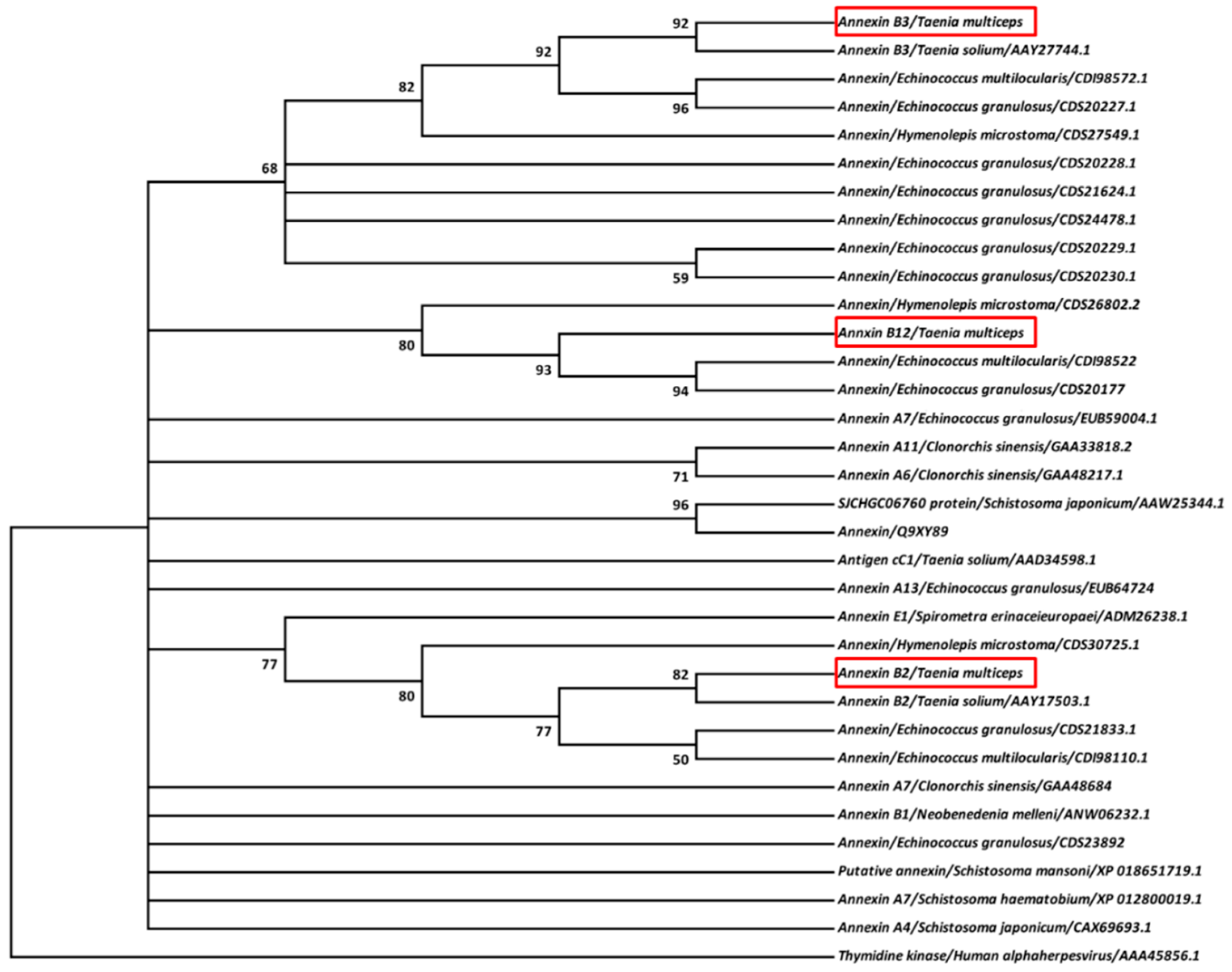
3.1. Molecular Features of Taenia multiceps Annexins
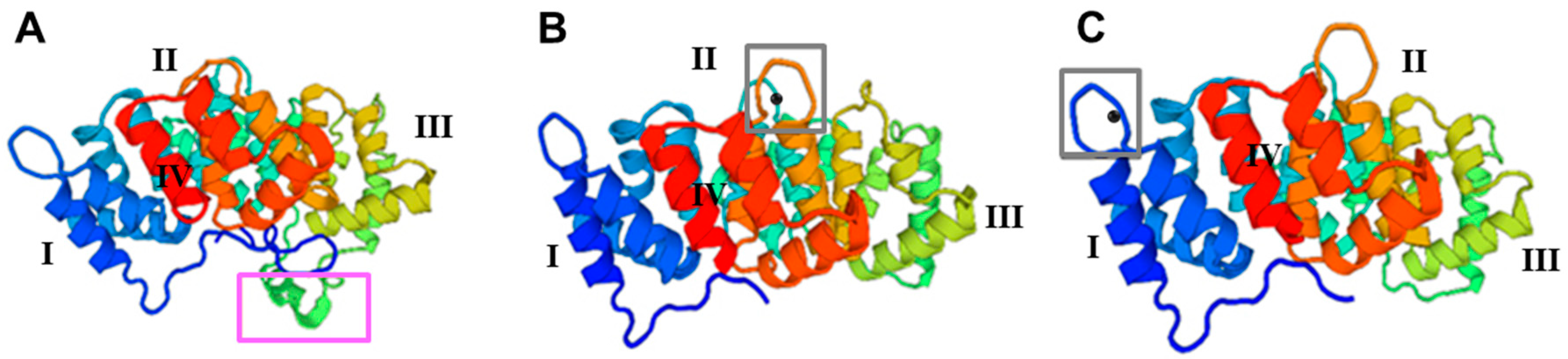
3.2. Structural Analysis of Taenia multiceps Annexins
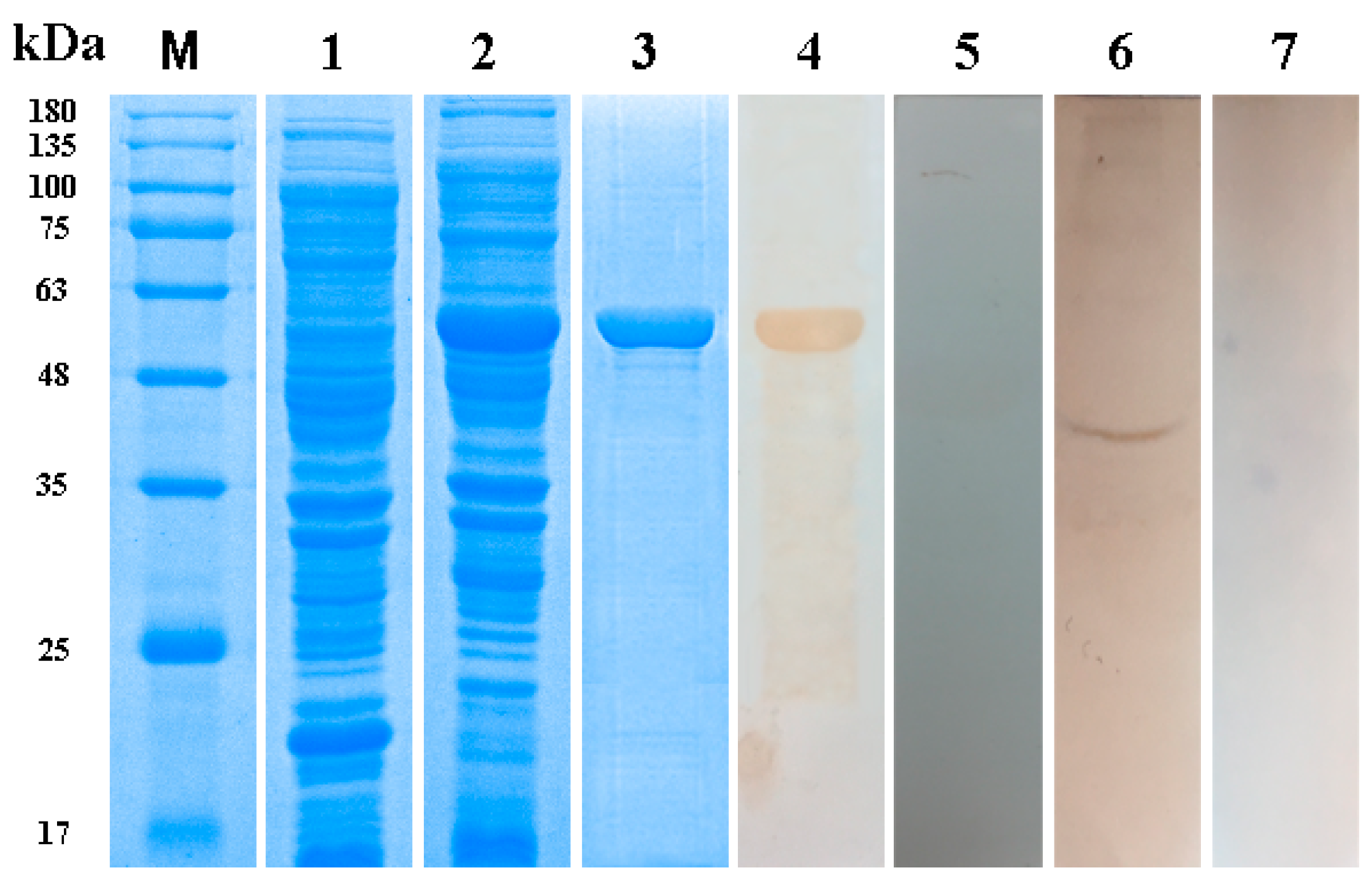
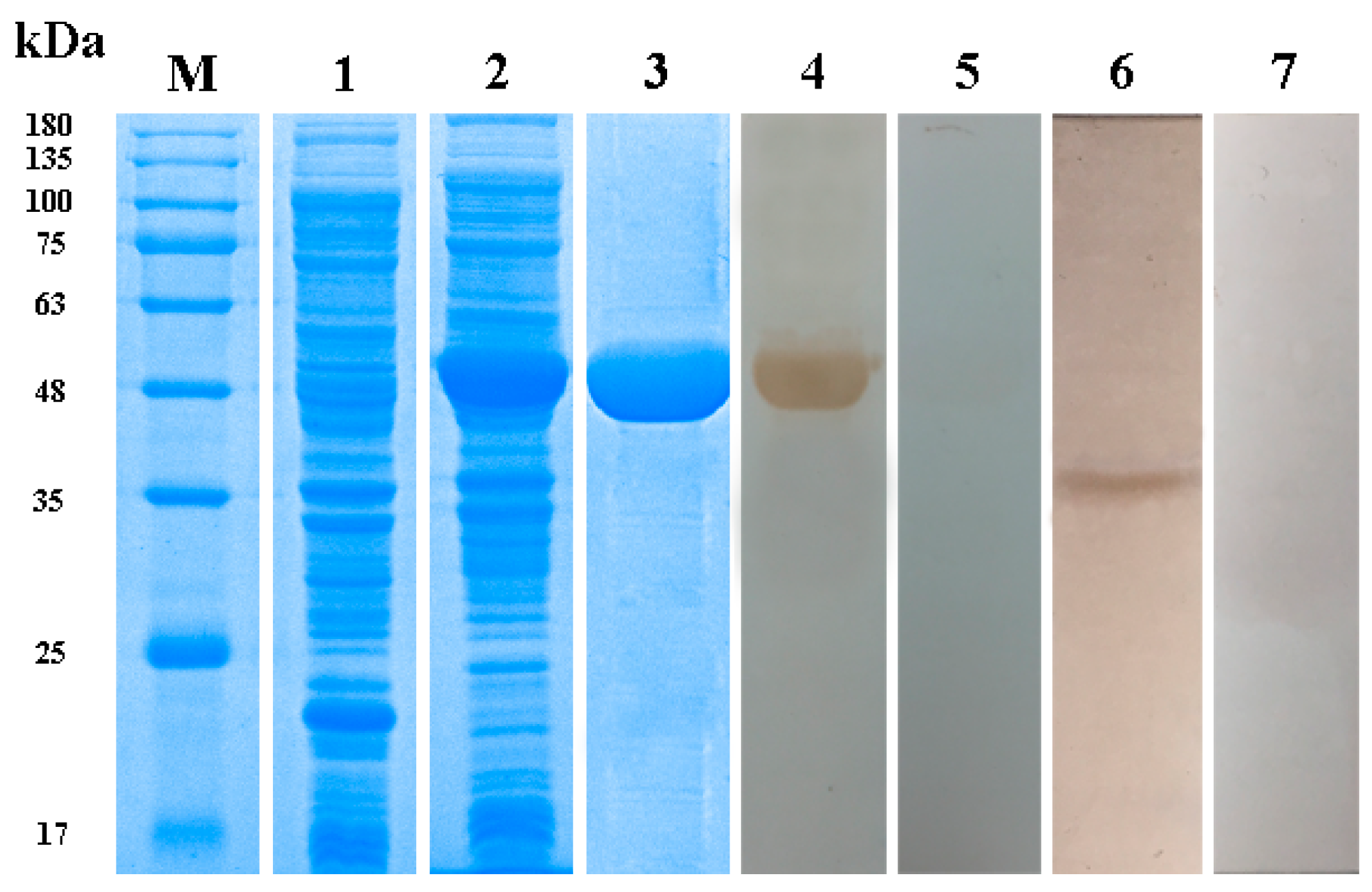
3.3. Expression, Purification and Western Blotting of Taenia multiceps Annexins
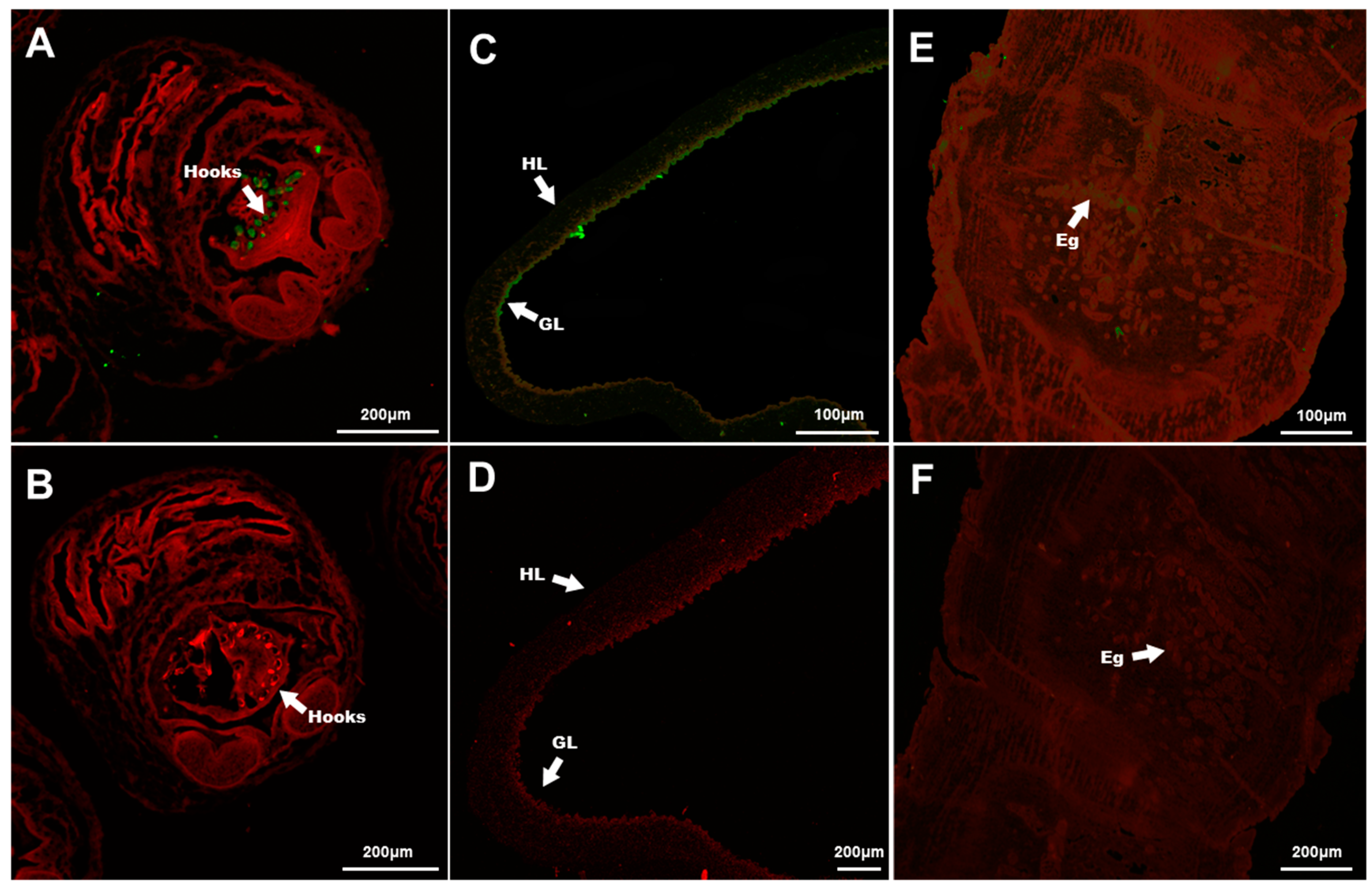
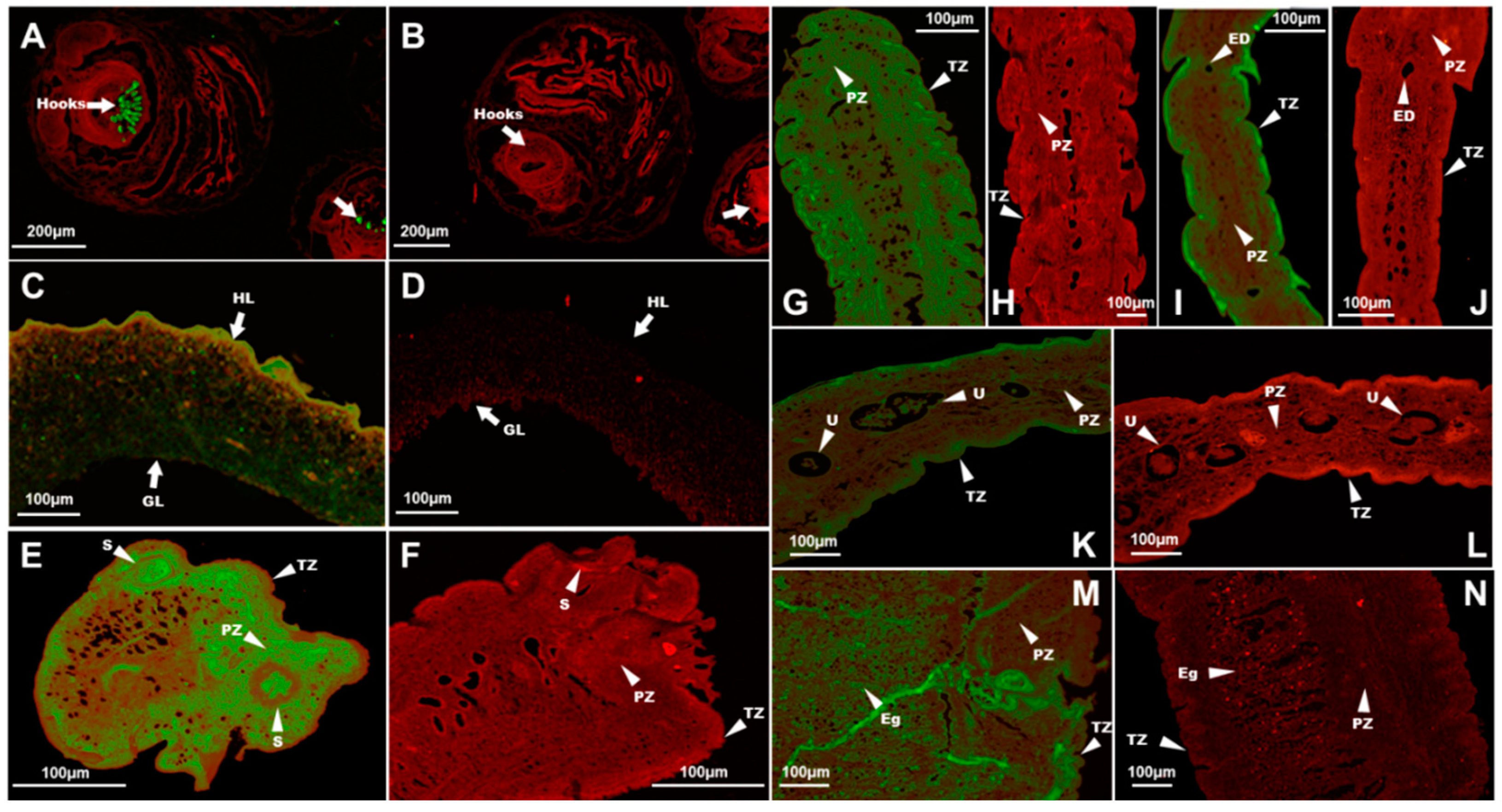
3.4. Immunolocalization of TmAnxB2 and TmAnxB3
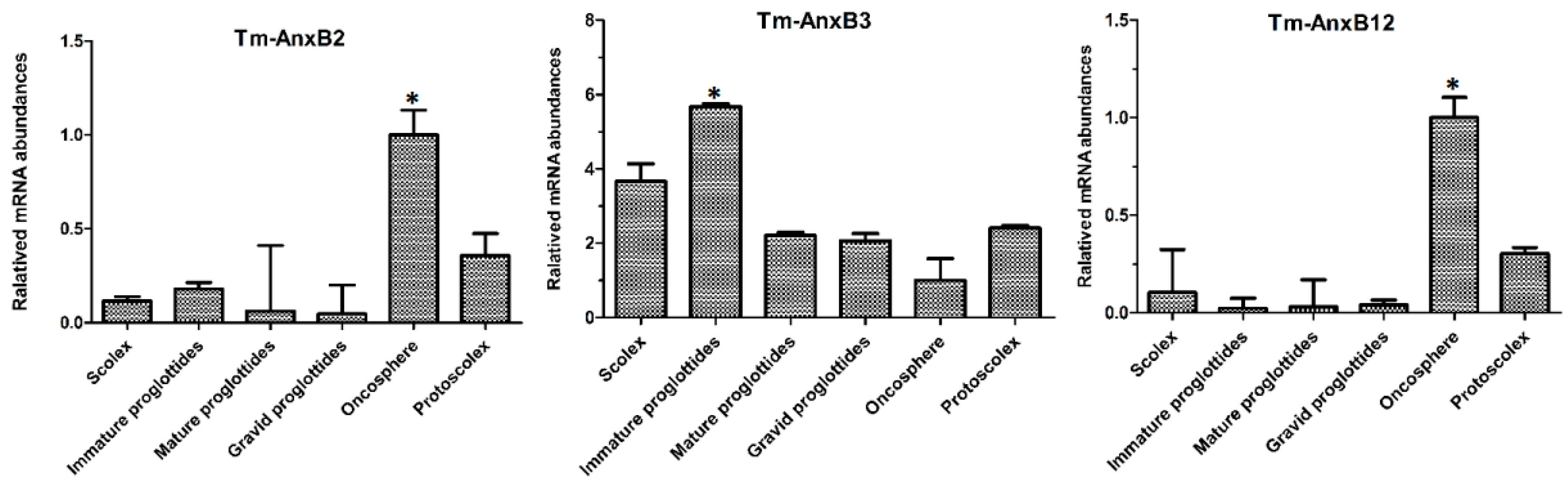
3.5. Transcriptional Profiles of Taenia multiceps Annexins
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Li, W.; Liu, B.; Yang, Y.; Ren, Y.; Wang, S.; Liu, C.; Zhang, N.; Qu, Z.; Yang, W.; Zhang, Y.; et al. The genome of tapeworm Taenia multiceps sheds light on understanding parasitic mechanism and control of coenurosis disease. DNA Res. 2018, in press. [Google Scholar] [CrossRef] [PubMed]
- Li, W.H.; Jia, W.Z.; Qu, Z.G.; Xie, Z.Z.; Luo, J.X.; Yin, H.; Sun, X.L.; Blaga, R.; Fu, B.Q. Molecular characterization of Taenia multiceps isolates from Gansu province, China by sequencing of mitochondrial cytochrome c oxidase subunit 1. Korean J. Parasitol. 2013, 51, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Xu, J.; Wang, Y.; Guo, C.; Chen, L.; Gu, X.; Lai, W.; Peng, X.; Yang, G. GP50 as a promising early diagnostic antigen for Taenia multiceps infection in goats by indirect ELISA. Parasit. Vectors 2016, 9, 618. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wang, Y.; Ye, Q.; Yang, Y.; Wan, J.; Guo, C.; Zhan, J.; Gu, X.; Lai, W.; Xie, Y.; et al. Development of a direct PCR assay to detect Taenia multiceps eggs isolated from dog feces. Vet. Parasitol. 2018, 251, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Oryan, A.; Moazeni, M.; Amrabadi, O.; Akbari, M.; Sharifiyazdi, H. Comparison of distribution pattern, pathogenesis and molecular characteristics of larval stages of Taenia multiceps in sheep and goats. Small Rumin. Res. 2015, 132, 44–49. [Google Scholar] [CrossRef]
- Christodoulopoulos, G.; Dinkel, A.; Romig, T.; Ebi, D.; Mackenstedt, U.; Loos-Frank, B. Cerebral and non-cerebral coenurosis: On the genotypic and phenotypic diversity of Taenia multiceps. Parasitol. Res. 2016, 115, 4543–4558. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Wang, Y.; Huang, X.; Wang, N.; Yan, M.; He, R.; Gu, X.; Xie, Y.; Lai, W.; Jing, B.; et al. Molecular cloing and bioinformatics analysis of lactate dehydrogenase from Taenia multiceps. Parasitol. Res. 2017, 116, 2845–2852. [Google Scholar] [CrossRef] [PubMed]
- Nie, H.M.; Xie, Y.; Fu, Y.; Yang, Y.D.; Gu, X.B.; Wang, S.X.; Peng, X.; Lai, W.M.; Peng, X.R.; Yang, G.Y. Cloning and characterization of the fatty acid-binding protein gene from the protoscolex of Taenia multiceps. Parasitol. Res. 2013, 112, 1833–1839. [Google Scholar] [CrossRef] [PubMed]
- Amer, S.; ElKhatam, A.; Fukuda, Y.; Bakr, LI.; Zidan, S.; Elsify, A.; Mohamed, M.A.; Tada, C.; Nakai, Y. Clinical, pathological, and molecular data concerning Coenurus cerebralis in sheep in Egypt. Data Brief 2018, 16, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dalimi, A.; Sattari, A.; Motamedi, G. A study on intestinal helminthes of dogs, foxes and jackals in the western part of Iran. Vet. Parasitol. 2006, 142, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Aliye, J.; Deressa, F.B. Prevalence and economic importance of Coenurus Cerebralis in sheep and goats in and around “Legahida” district of bale zone, south eastern Ethiopia. Comp. Clin. Pathol. 2017, 26, 483–492. [Google Scholar] [CrossRef]
- Wu, X.; Fu, Y.; Yang, D.; Xie, Y.; Zhang, R.; Zheng, W.; Nie, H.; Yan, N.; Wang, N.; Wang, J. Identification of neglected cestode Taenia multiceps microRNAs by illumina sequencing and bioinformatic analysis. BMC Vet. Res. 2013, 9, 162. [Google Scholar] [CrossRef] [PubMed]
- Collomb, J.; Machouart, M.; Biava, MF.; Brizion, M.; Montagne, K.; Plénat, F.; Fortier, B. Contribution of NADH dehydrogenase subunit I and cytochrome c oxidase subunit I sequences toward identifying a case of human coenuriasis in France. J. Parasitol. 2007, 93, 934–937. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nie, H.; Gu, X.; Wang, T.; Huang, X.; Chen, L.; Lai, W.; Peng, X.; Yang, G. An ELISA using recombinant TmHSP70 for the diagnosis of Taenia multiceps infections in goats. Vet. Parasitol. 2015, 212, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Rescher, U.; Gerke, V. Annexins—Unique membrane binding proteins with diverse functions. J. Cell Sci. 2004, 117, 2631–2639. [Google Scholar] [CrossRef] [PubMed]
- Chander, A.; Chen, X.L.; Naidu, D.G. A role for diacylglycerol in annexin a 7-mediated fusion of lung lamellar bodies. Biochim. Biophys. Acta 2007, 1771, 1308–1318. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, J.C.; Laohavisit, A.; Macpherson, N.; Webb, A.; Brownlee, C.; Battey, N.H.; Davies, J.M. Annexins: Multifunctional components of growth and adaptation. J. Exp. Bot. 2008, 59, 533. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, A.; Osman, A.; Leow, C.Y.; Driguez, P.; McManus, D.P.; Jones, M.K. Parasite annexins—New molecules with potential for drug and vaccine development. Bioessays 2010, 32, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.; Yang, Y.D.; Yang, G.Y.; Wang, Y.W.; An, X.X.; Gu, X.B.; Bian, Y.; Fu, Y.; Wang, S.X. Biological characteristics of Taenia multiceps Sichuan strain. Chin. Vet. Sci. 2010, 36, 327–330. [Google Scholar]
- Cantacessi, C.; Seddon, J.M.; Miller, T.L.; Leow, C.Y.; Thomas, L.; Mason, L.; Willis, C.; Walker, G.; Loukas, A.; Gasser, R.B. A genome-wide analysis of annexins from parasitic organisms and their vectors. Sci. Rep. 2013, 3, 2893. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Hu, D.; Zhong, X.; Wang, N.; Gu, X.; Wang, T.; Peng, X.; Yang, G. Characterization of a secretory annexin in Echinococcus granulosus. Am. J. Trop. Med. Hyg. 2016, 94, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Tararam, C.A.; Farias, L.P.; Wilson, R.A.; Leite, L.C. Schistosoma mansoni annexin 2: Molecular characterization and immunolocalization. Exp. Parasitol. 2010, 126, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Morgan, R.O.; Martin-Almedina, S.; Iglesias, J.M.; Gonzalez-Florez, M.I.; Fernandez, M.P. Evolutionary perspective on annexin calcium-binding domains. Biochim. Biophys. Acta 2004, 1742, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Dissous, C.; Khayath, N.; Vicogne, J.; Capron, M. Growth factor receptors in helminth parasites: Signalling and host-parasite relationships. FEBS Lett. 2006, 580, 2968–2975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahia, D.; Andrade, L.F.; Ludolf, F.; Mortara, R.A.; Oliveira, G. Protein tyrosine kinases in Schistosoma mansoni. Mem. Inst. Oswaldo Cruz 2006, 101, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Brehm, K. The role of evolutionarily conserved signalling systems in Echinococcus multilocularis development and host-parasite interaction. Med. Microbiol. Immunol. 2010, 199, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Cheetham, G.M. Novel protein kinases and molecular mechanisms of autoinhibition. Curr. Opin. Struct. Biol. 2004, 14, 700–705. [Google Scholar] [CrossRef] [PubMed]
- Weingärtner, A.; Kemmer, G.; Müller, F.D.; Zampieri, R.A.; Gonzaga dos Santos, M.; Schiller, J.; Pomorski, T.G. Leishmania promastigotes lack phosphatidylserine but bind annexin V upon permeabilization or miltefosine treatment. PLoS ONE 2012, 7, e42070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solano, S.; Cortés, I.M.; Copitin, N.I.; Tato, P.; Molinari, J.L. Lymphocyte apoptosis in the inflammatory reaction around Taenia solium metacestodes in porcine cysticercosis. Vet. Parasitol. 2006, 140, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.J.; Yan, H.L.; Ding, F.X.; Lu, Y.M.; Sun, S.H. Annexin B1 at the host-parasite interface of the Taenia solium cysticercus: Secreted and associated with inflammatory reaction. Acta Trop. 2007, 101, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Hamamouch, N.; Li, C.; Hewezi, T.; Hussey, R.S.; Baum, T.J.; Mitchum, M.G.; Davis, E.L. A nematode effector protein similar to annexins in host plants. J. Exp. Bot. 2010, 61, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Guo, Y.; Li, K.; Lu, Y.; Zhang, Y.; Sun, S.; Yan, H.; Zhang, S. Molecular characterization and anticoagulant activity of a novel annexin derived from the Taenia solium. Acta Trop. 2006, 99, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.L.; Xue, G.; Mei, Q.; Ding, F.X.; Wang, Y.Z.; Sun, S.H. Calcium-dependent proapoptotic effect of Taenia solium metacestodes annexin B1 on human eosinophils: A novel strategy to prevent host immune response. Int. J. Biochem. Cell. Biol. 2008, 40, 2151–2163. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Ren, M.; Chen, X.; Wang, X.; Li, S.; Lin, J.; Liang, C.; Liang, P.; Hu, Y.; Lei, H.; et al. Biochemical and immunological characterization of annexin B30 from Clonorchis sinensis excretory/secretory products. Parasitol. Res. 2014, 113, 2743–2755. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.X.; Li, T.L.; Suo, Y.L. Advances in annexin of Taeniidae. Contemp. Anim. Husb. 2015, 3–4. [Google Scholar] [CrossRef]
- Dai, J.L.; Huang, J.; Liao, X.J.; Li, B.; Wang, Y. The expression and purification of the annexins B2 gene in Taenia saginata asiatica and the immunologic analysis of the recombinant proteins. J. Xi’an Jiaotong Univ. 2010, 30, 696–698. [Google Scholar]
- Wang, Z.W.; Han, H.Y.; Huang, B. Four alctium signaling proteins and their preliminary studies in parasitology. Chin. J. Anim. Infect. Dis. 2015, 23, 78–86. [Google Scholar]
- Wang, Z.X.; Zhang, Y.Y.; Lu, P.P.; Ma, X.; Wang, Z.R.; Bo, X.W. The role of wnt signaling pathway in the flatworm. Chin. J. Anim. Vet. Sci. 2017, 48, 207–213. [Google Scholar]
- Wang, Z.R.; Zhang, Y.Y.; Bo, X.W.; Xu, X.P.; Xu, C.S. Differential expression of six wnt gene family members in Echinococcus granulosus protoscoleces and adult worms. Chin. J. Parasitol. Parasit. Dis. 2016, 34, 552–557. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primers | Restriction Enzyme Sites | Reference Sequences |
|---|---|---|---|
| TmAnxB2 | F: CGCGGATCCATGGCAAAAAATACTCGCTC | BamHI | Unigene18519 |
| R: CCGCTCGAGTTAGGATTCATTCAGTAGTGCG | XhoI | Ts AY998562.1 | |
| TmAnxB3 | F: CCGGAATTCATGGCGACTGTCAAGCTTT | EcoRI | Unigene19512 |
| R: CCGCTCGAGTCACTCCAGTATGGTGAGCA | XhoI | Ts DQ010543.1 | |
| TmAnxB12 | F: CGCGGATCCATGAATGGGCGTCCAACTA | BamHI | Unigene17682 |
| R: CCGGAATTCTTATGCATTCCCCTCTACAA | EcoRI | EgrG_000237700 |
| Gene | Primers | Size (bp) |
|---|---|---|
| TmAnxB2 | F: GGTTCAACACGCCGTAGACAGAC | 101 |
| R: TGAGGACTCGCATGAGGAGGAAG | ||
| TmAnxB3 | F: TGCACCGCCACCAACAACG | 114 |
| R: CACTCGCTTGAAGTCGCCAGAG | ||
| TmAnxB12 | F: AGGAGGTGACGGAGATGAAGGC | 94 |
| R: GCAACGGCGATGATCTGGATGAG | ||
| Actin | F: CTAAGGCGAACCGTGAGAAGATGAC | 188 |
| R: GGCATGAGGCAAGGCGTAACC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, C.; Xie, Y.; Liu, Y.; Wang, N.; Zhan, J.; Zhou, X.; Angel, C.; Gu, X.; Lai, W.; Peng, X.; et al. Molecular Characterization of Annexin B2, B3 and B12 in Taenia multiceps. Genes 2018, 9, 559. https://doi.org/10.3390/genes9110559
Guo C, Xie Y, Liu Y, Wang N, Zhan J, Zhou X, Angel C, Gu X, Lai W, Peng X, et al. Molecular Characterization of Annexin B2, B3 and B12 in Taenia multiceps. Genes. 2018; 9(11):559. https://doi.org/10.3390/genes9110559
Chicago/Turabian StyleGuo, Cheng, Yue Xie, Yuchen Liu, Ning Wang, Jiafei Zhan, Xuan Zhou, Christiana Angel, Xiaobin Gu, Weimin Lai, Xuerong Peng, and et al. 2018. "Molecular Characterization of Annexin B2, B3 and B12 in Taenia multiceps" Genes 9, no. 11: 559. https://doi.org/10.3390/genes9110559
APA StyleGuo, C., Xie, Y., Liu, Y., Wang, N., Zhan, J., Zhou, X., Angel, C., Gu, X., Lai, W., Peng, X., & Yang, G. (2018). Molecular Characterization of Annexin B2, B3 and B12 in Taenia multiceps. Genes, 9(11), 559. https://doi.org/10.3390/genes9110559