Molecular Factors of Hypochlorite Tolerance in the Hypersaline Archaeon Haloferax volcanii


 ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Strains and Media
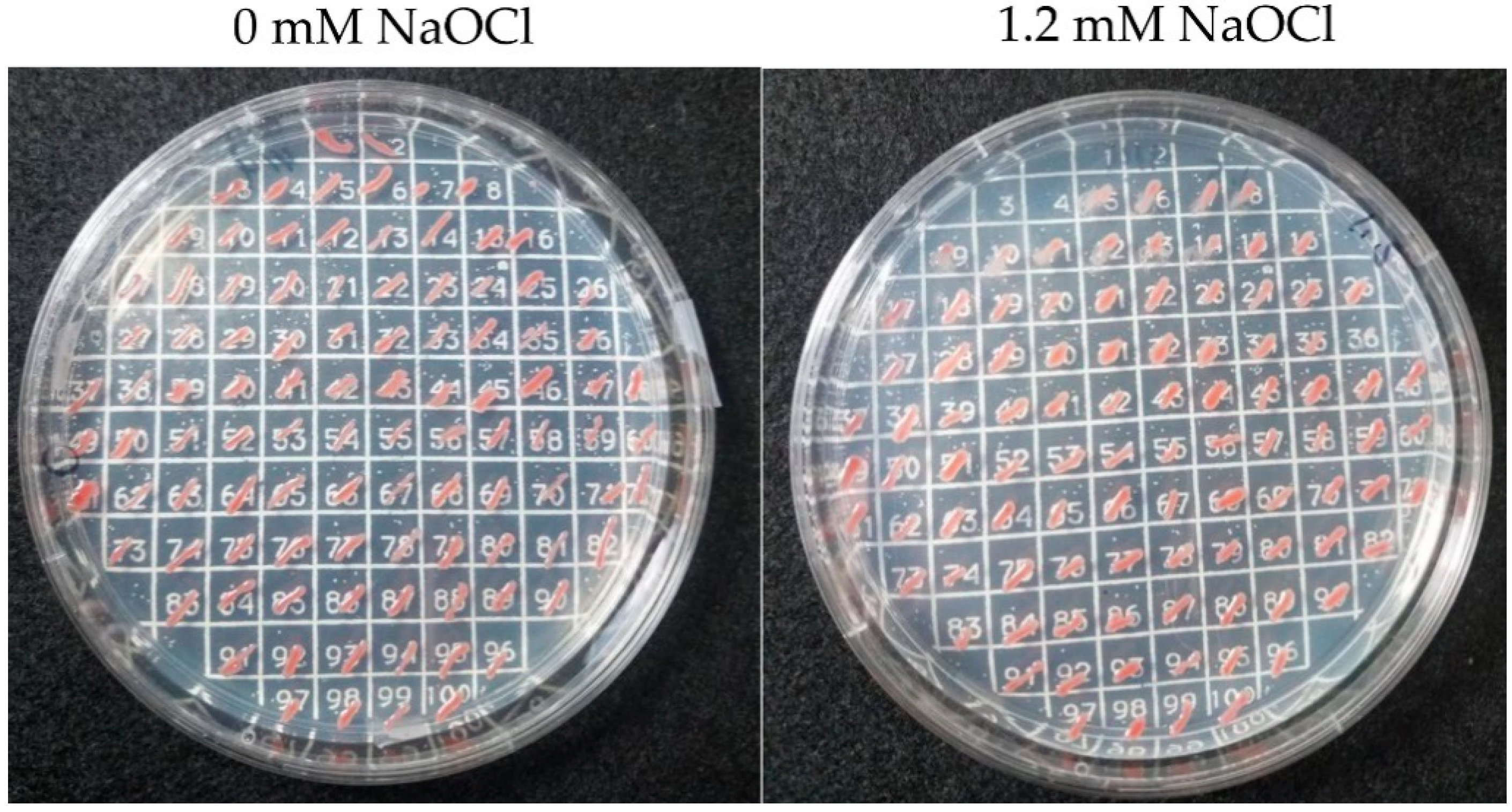
2.3. Isolation of Mutants with Enhanced Tolerance to Hypochlorite Stress
2.4. Genomic DNA Isolation
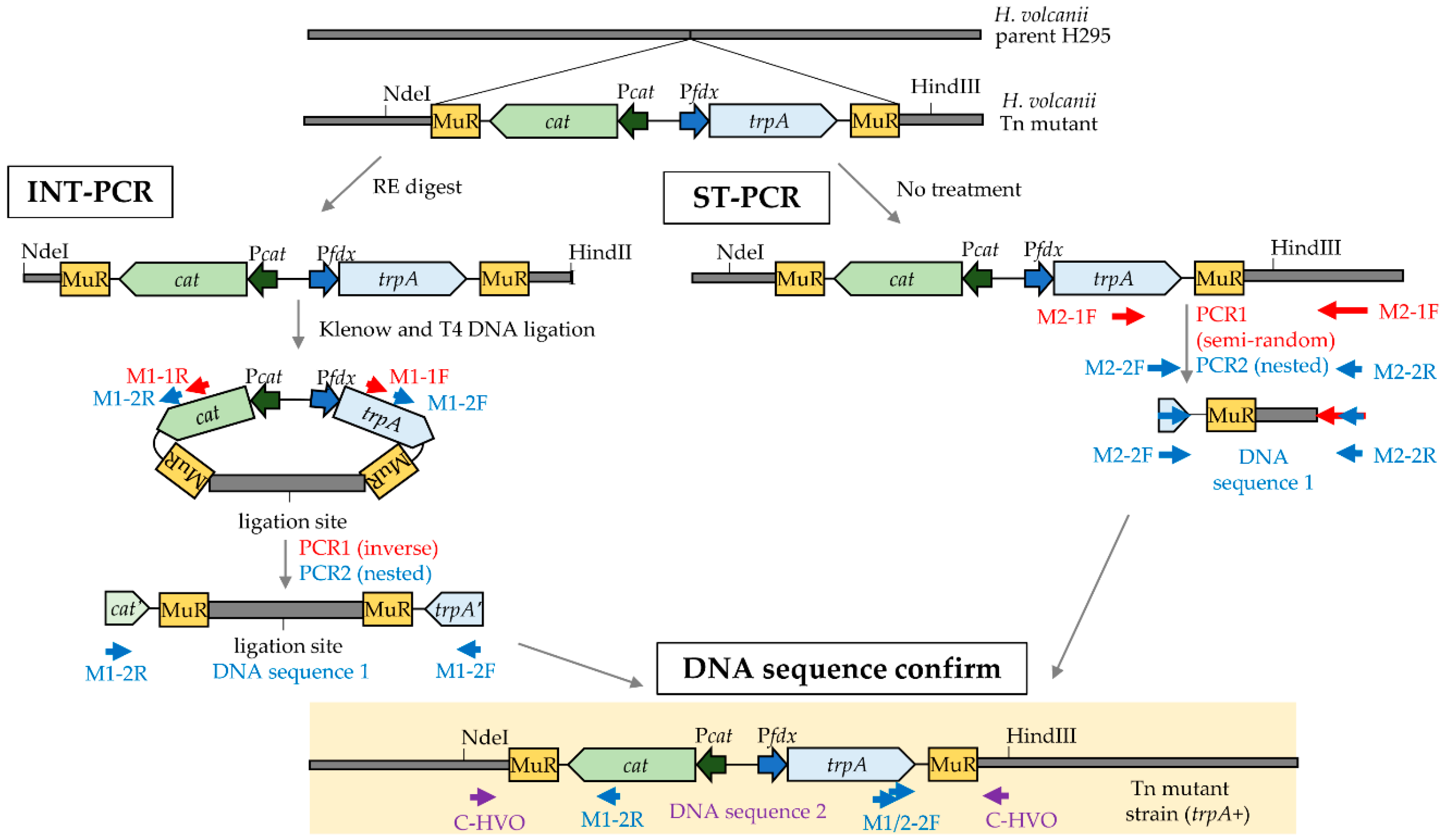
2.5. Identification of Transposon Insertion Sites
2.5.1. Inversed Nested Two-Step PCR
2.5.2. Semi-Random Two-Step PCR
2.5.3. DNA Sequencing to Identify the Transposon Insertion Sites
2.6. PCR Conditions
2.7. RNA Isolation and Real-time Quantitative Reverse Transcription PCR
2.8. Hypochlorite Stress Plate Assay
2.9. Hypochlorite Stress Liquid Assay
2.10. SDS-PAGE and Immunoblotting Analysis
2.11. Prediction of Protein Structure and Function
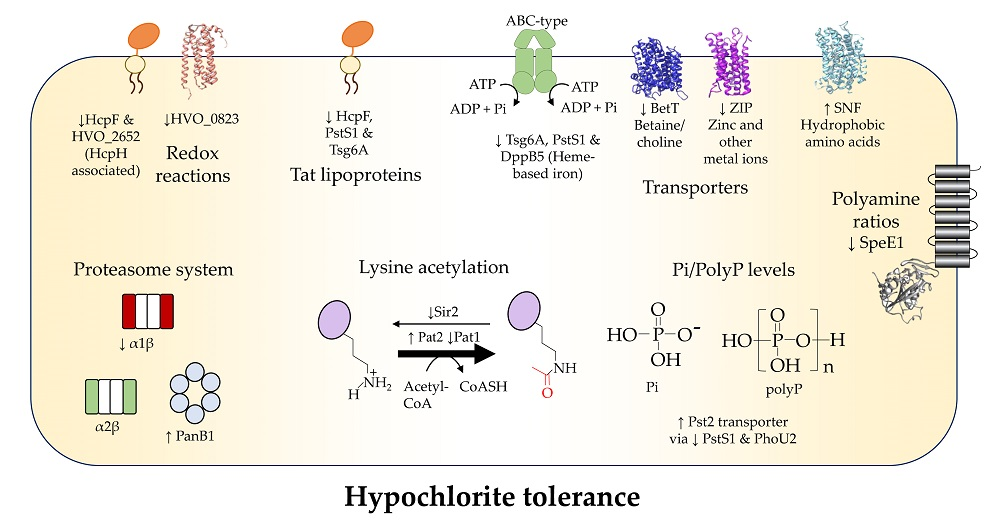
3. Results and Discussion
3.1. Haloferax volcanii Mutants of Enhanced Tolerance to Hypochlorite Stress
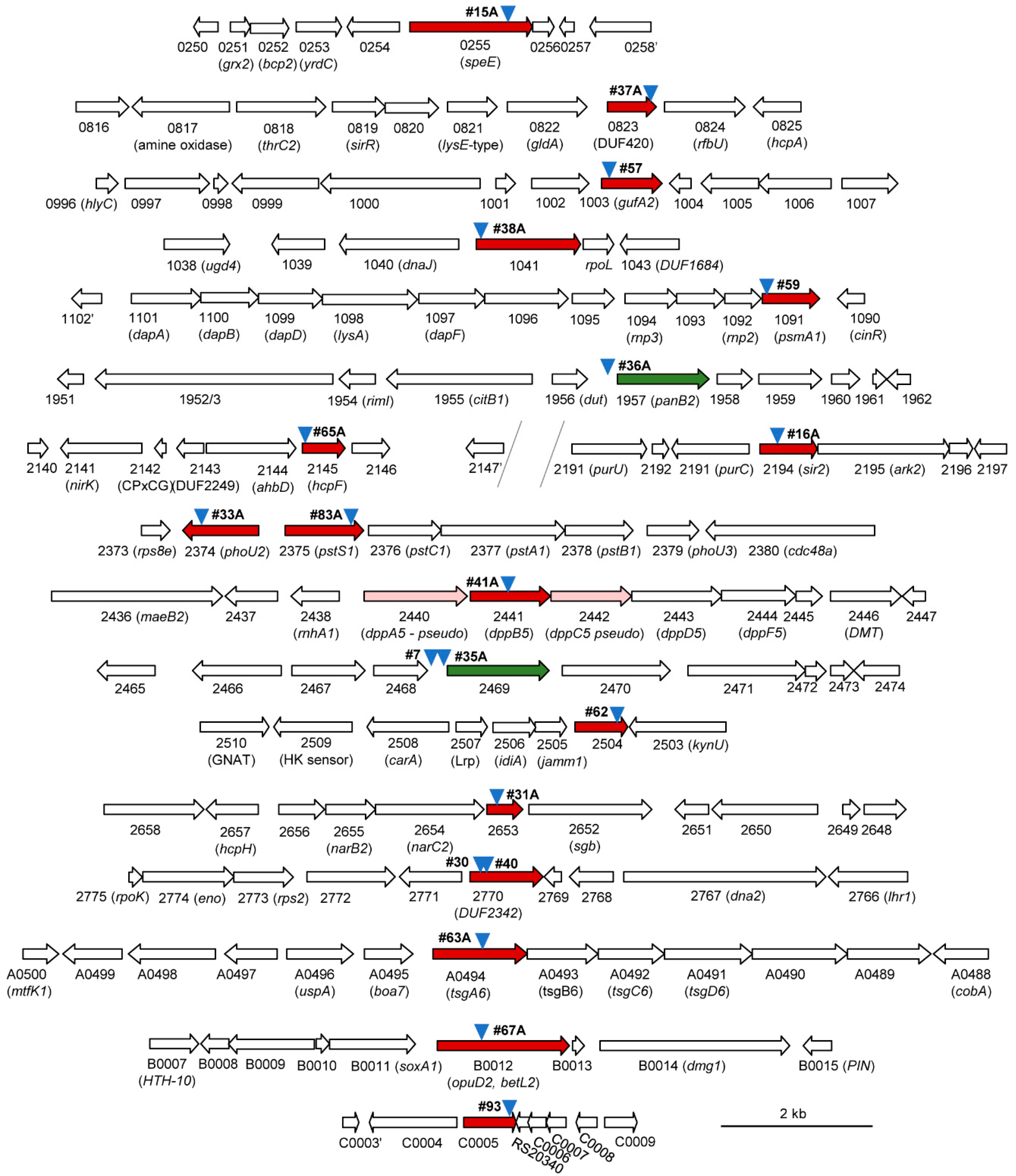
3.2. Transposon Insertion Sites Mapped on the Genome of Haloferax volcanii Mutant Strains were Found to be Hypertolerant to Hypochlorite
3.3. Membrane versus Intracellular Functions
3.4. Metal Ion Transport
3.5. Inorganic Phosphate Transport
3.6. Organic Molecule Transport
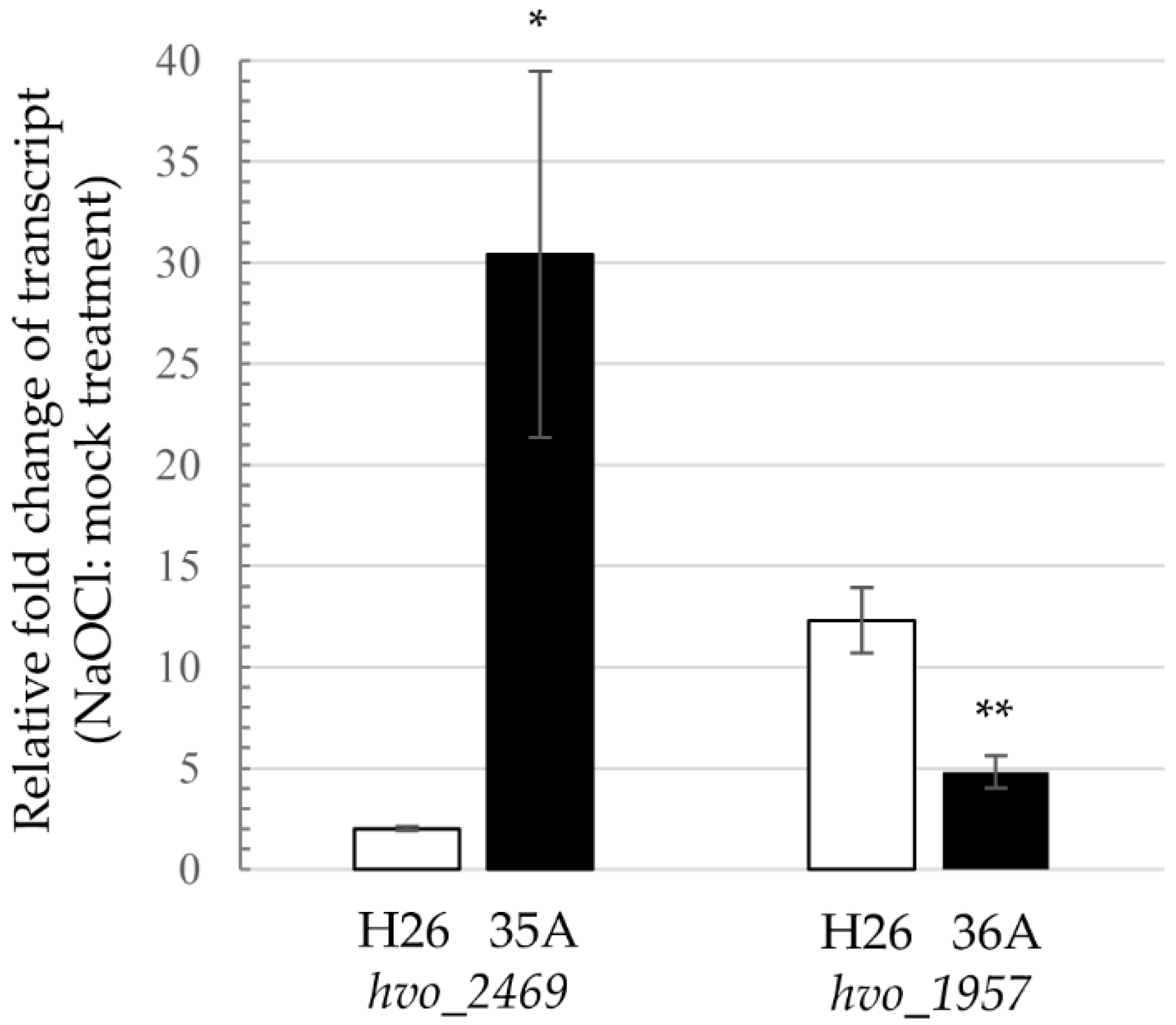
3.7. Polyamine Synthesis
3.8. Membrane Associated Redox Reactions
3.9. Oxidoreductase and Hydrolase Enzymes
3.10. Protein Lysine Deacetylation
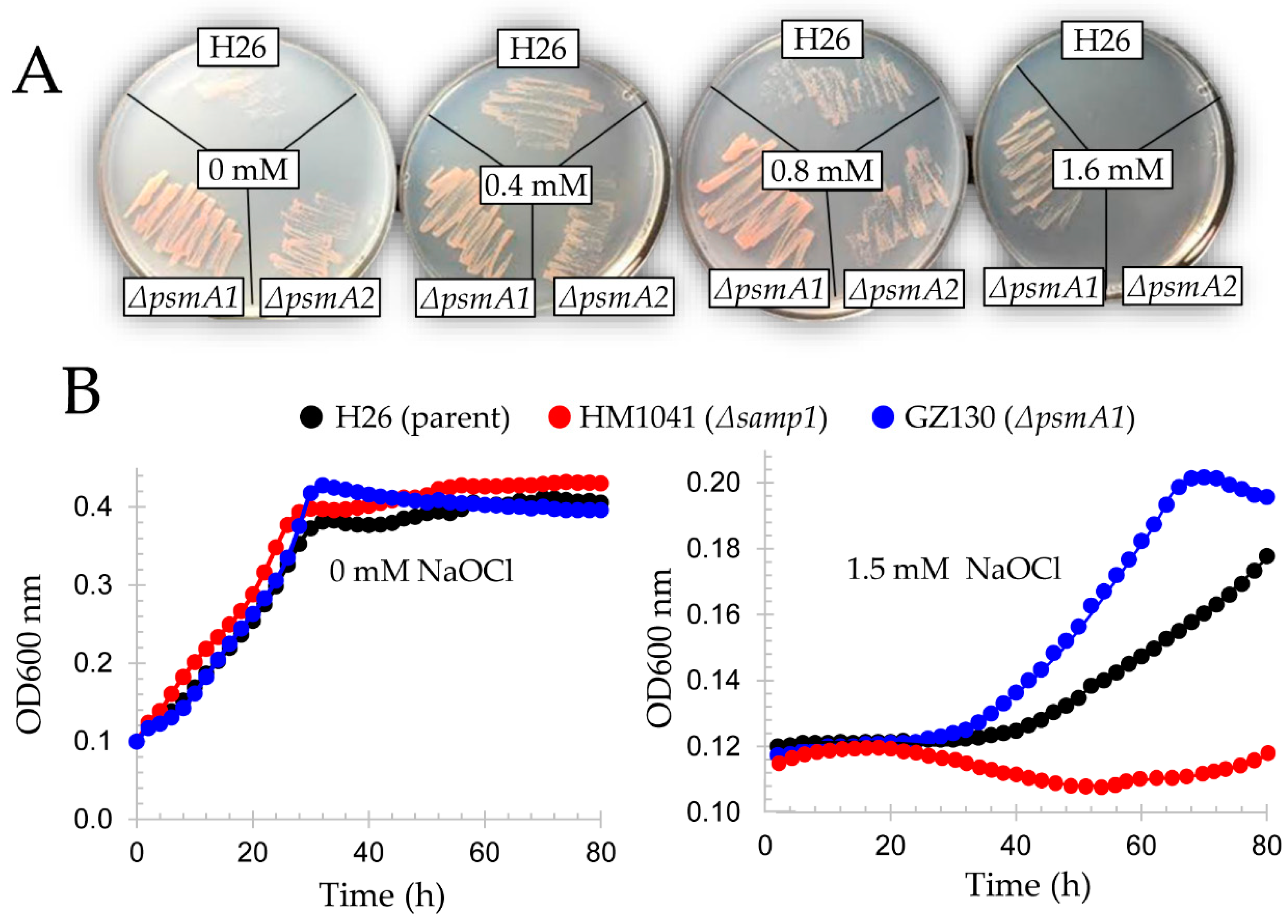
3.11. Proteasome Components
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Riley, P.A. Free radicals in biology: Oxidative stress and the effects of ionizing radiation. Int. J. Radiat. Biol. 1994, 65, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Panasenko, O.M.; Gorudko, I.V.; Sokolov, A.V. Hypochlorous acid as a precursor of free radicals in living systems. Biochemistry (Mosc.) 2013, 78, 1466–1489. [Google Scholar] [CrossRef] [PubMed]
- Cabiscol, E.; Tamarit, J.; Ros, J. Oxidative stress in bacteria and protein damage by reactive oxygen species. Int. Microbiol. 2000, 3, 3–8. [Google Scholar] [PubMed]
- Pratt, D.A.; Tallman, K.A.; Porter, N.A. Free radical oxidation of polyunsaturated lipids: New mechanistic insights and the development of peroxyl radical clocks. Acc. Chem. Res. 2011, 44, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Reichmann, D.; Voth, W.; Jakob, U. Maintaining a healthy proteome during oxidative stress. Mol. Cell. 2018, 69, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.L.; Baxter, B.K. DNA repair and photoprotection: Mechanisms of overcoming environmental ultraviolet radiation exposure in halophilic archaea. Front. Microbiol. 2017, 8, 1882. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Bermudez, P.; Srebotnik, E.; Hammel, K.E. Chlorination and cleavage of lignin structures by fungal chloroperoxidases. Appl. Environ. Microbiol. 2003, 69, 5015–5018. [Google Scholar] [CrossRef] [PubMed]
- Wang, G. Chloride flux in phagocytes. Immunol. Rev. 2016, 273, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Loi, V.V.; Rossius, M.; Antelmann, H. Redox regulation by reversible protein S-thiolation in bacteria. Front. Microbiol. 2015, 6, 187. [Google Scholar] [CrossRef] [PubMed]
- Wardman, P.; Candeias, L.P. Fenton chemistry: An introduction. Radiat. Res. 1996, 145, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Kiljunen, S.; Pajunen, M.I.; Dilks, K.; Storf, S.; Pohlschroder, M.; Savilahti, H. Generation of comprehensive transposon insertion mutant library for the model archaeon, Haloferax volcanii, and its use for gene discovery. BMC Biol. 2014, 12, 103. [Google Scholar] [CrossRef]
- Mullakhanbhai, M.F.; Larsen, H. Halobacterium volcanii spec. nov., a Dead Sea halobacterium with a moderate salt requirement. Arch. Microbiol. 1975, 104, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Haapa, S.; Taira, S.; Heikkinen, E.; Savilahti, H. An efficient and accurate integration of mini-Mu transposons in vitro: A general methodology for functional genetic analysis and molecular biology applications. Nucleic Acids Res. 1999, 27, 2777–2784. [Google Scholar] [CrossRef] [PubMed]
- Kiljunen, S.; Pajunen, M.I.; Savilahti, H. Transposon insertion mutagenesis for archaeal gene discovery. Methods Mol. Biol. 2017, 1498, 309–320. [Google Scholar] [CrossRef] [PubMed]
- McMillan, L.J.; Hwang, S.; Farah, R.E.; Koh, J.; Chen, S.; Maupin-Furlow, J.A. Multiplex quantitative SILAC for analysis of archaeal proteomes: A case study of oxidative stress responses. Environ. Microbiol. 2018, 20, 385–401. [Google Scholar] [CrossRef] [PubMed]
- Nissenbaum, A. The microbiology and biogeochemistry of the Dead Sea. Microb. Ecol. 1975, 2, 139–161. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, K.; Cano, D.; Maupin-Furlow, J. Glycerol-mediated repression of glucose metabolism and glycerol kinase as the sole route of glycerol catabolism in the haloarchaeon Haloferax volcanii. J. Bacteriol. 2009, 191, 4307–4315. [Google Scholar] [CrossRef] [PubMed]
- Legerme, G.; Yang, E.; Esquivel, R.N.; Kiljunen, S.; Savilahti, H.; Pohlschroder, M. Screening of a Haloferax volcanii transposon library reveals novel motility and adhesion mutants. Life (Basel) 2016, 6, E41. [Google Scholar] [CrossRef] [PubMed]
- Dyall-Smith, M. The Halohandbook: Protocols for Halobacterial Genetics v.7.2. 2009. Available online: http://www.haloarchaea.com/resources/halohandbook/Halohandbook_2009_v7.2mds.pdf (accessed on 19 November 2018).
- Chun, K.T.; Edenberg, H.J.; Kelley, M.R.; Goebl, M.G. Rapid amplification of uncharacterized transposon-tagged DNA sequences from genomic DNA. Yeast 1997, 13, 233–240. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kaczowka, S.J.; Maupin-Furlow, J.A. Subunit topology of two 20S proteasomes from Haloferax volcanii. J. Bacteriol. 2003, 185, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Attwood, T.K.; Babbitt, P.C.; Bateman, A.; Bork, P.; Bridge, A.J.; Chang, H.Y.; Dosztanyi, Z.; El-Gebali, S.; Fraser, M.; et al. InterPro in 2017-beyond protein family and domain annotations. Nucleic Acids Res. 2017, 45, D190–D199. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H. Predicting secretory proteins with SignalP. Methods Mol. Biol. 2017, 1611, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Bendtsen, J.D.; Nielsen, H.; Widdick, D.; Palmer, T.; Brunak, S. Prediction of twin-arginine signal peptides. BMC Bioinform. 2005, 6, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, R.W.; Bruser, T.; Kissinger, J.C.; Pohlschroder, M. Adaptation of protein secretion to extremely high-salt conditions by extensive use of the twin-arginine translocation pathway. Mol. Microbiol. 2002, 45, 943–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, N.Y.; Wagner, J.R.; Laird, M.R.; Melli, G.; Rey, S.; Lo, R.; Dao, P.; Sahinalp, S.C.; Ester, M.; Foster, L.J.; et al. PSORTb 3.0: Improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 2010, 26, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Schneider, K.L.; Pollard, K.S.; Baertsch, R.; Pohl, A.; Lowe, T.M. The UCSC Archaeal Genome Browser. Nucleic Acids Res. 2006, 34, D407–D410. [Google Scholar] [CrossRef] [PubMed]
- Oberto, J. SyntTax: A web server linking synteny to prokaryotic taxonomy. BMC Bioinform. 2013, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, P.; Humbard, M.; Daniels, C.; Maupin-Furlow, J. Shotgun proteomics of the haloarchaeon Haloferax volcanii. J. Proteome Res. 2008, 7, 5033–5039. [Google Scholar] [CrossRef] [PubMed]
- Gimenez, M.I.; Dilks, K.; Pohlschroder, M. Haloferax volcanii twin-arginine translocation substates include secreted soluble, C-terminally anchored and lipoproteins. Mol. Microbiol. 2007, 66, 1597–1606. [Google Scholar] [CrossRef] [PubMed]
- Porcheron, G.; Garenaux, A.; Proulx, J.; Sabri, M.; Dozois, C.M. Iron, copper, zinc, and manganese transport and regulation in pathogenic Enterobacteria: Correlations between strains, site of infection and the relative importance of the different metal transport systems for virulence. Front. Cell. Infect. Microbiol. 2013, 3, 90. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. The mismetallation of enzymes during oxidative stress. J. Biol. Chem. 2014, 289, 28121–28128. [Google Scholar] [CrossRef] [PubMed]
- Nieboer, E.; Richardson, D.H.S. The replacement of the nondescript term ‘heavy metal’ by a biologically significant and chemically significant classification of metal ions. Environ. Pollut. B 1980, 1, 3–26. [Google Scholar] [CrossRef]
- Letoffe, S.; Delepelaire, P.; Wandersman, C. The housekeeping dipeptide permease is the Escherichia coli heme transporter and functions with two optional peptide binding proteins. Proc. Natl. Acad. Sci. USA 2006, 103, 12891–12896. [Google Scholar] [CrossRef] [PubMed]
- Frawley, E.R.; Fang, F.C. The ins and outs of bacterial iron metabolism. Mol. Microbiol. 2014, 93, 609–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartman, A.; Norais, C.; Badger, J.; Delmas, S.; Haldenby, S.; Madupu, R.; Robinson, J.; Khouri, H.; Ren, Q.; Lowe, T.; et al. The complete genome sequence of Haloferax volcanii DS2, a model archaeon. PLoS ONE 2010, 5, e9605. [Google Scholar] [CrossRef] [PubMed]
- Furtwangler, K.; Tarasov, V.; Wende, A.; Schwarz, C.; Oesterhelt, D. Regulation of phosphate uptake via Pst transporters in Halobacterium salinarum R1. Mol. Microbiol. 2010, 76, 378–392. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lou, Y.; Yokota, H.; Adams, P.D.; Kim, R.; Kim, S.H. Crystal structure of a PhoU protein homologue: A new class of metalloprotein containing multinuclear iron clusters. J. Biol. Chem. 2005, 280, 15960–15966. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Han, H.; Lv, Z.; Lin, Z.; Shang, Y.; Xu, T.; Wu, Y.; Zhang, Y.; Qu, D. PhoU2 but not PhoU1 as an important regulator of biofilm formation and tolerance to multiple stresses by participating in various fundamental metabolic processes in Staphylococcus epidermidis. J. Bacteriol. 2017, 199, e00219-17. [Google Scholar] [CrossRef] [PubMed]
- Tam, R.; Saier, M.H., Jr. Structural, functional, and evolutionary relationships among extracellular solute-binding receptors of bacteria. Microbiol Rev. 1993, 57, 320–346. [Google Scholar] [PubMed]
- Ziegler, C.; Bremer, E.; Kramer, R. The BCCT family of carriers: From physiology to crystal structure. Mol. Microbiol. 2010, 78, 13–34. [Google Scholar] [CrossRef] [PubMed]
- Schweikhard, E.S.; Ziegler, C.M. Amino acid secondary transporters: Toward a common transport mechanism. Curr. Top. Membr. 2012, 70, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Lamark, T.; Styrvold, O.B.; Strom, A.R. Efflux of choline and glycine betaine from osmoregulating cells of Escherichia coli. FEMS Microbiol. Lett. 1992, 75, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Malinauskaite, L.; Quick, M.; Reinhard, L.; Lyons, J.A.; Yano, H.; Javitch, J.A.; Nissen, P. A mechanism for intracellular release of Na+ by neurotransmitter/sodium symporters. Nat. Struct. Mol. Biol. 2014, 21, 1006–1012. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Xing, L.; Fu, Q.; Zhou, G.H.; Zhang, W.G. A review of antioxidant peptides derived from meat muscle and by-products. Antioxidants (Basel) 2016, 5, E32. [Google Scholar] [CrossRef] [PubMed]
- Agostinelli, E.; Marques, M.P.; Calheiros, R.; Gil, F.P.; Tempera, G.; Viceconte, N.; Battaglia, V.; Grancara, S.; Toninello, A. Polyamines: Fundamental characters in chemistry and biology. Amino Acids 2010, 38, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Rhee, H.J.; Kim, E.J.; Lee, J.K. Physiological polyamines: Simple primordial stress molecules. J. Cell. Mol. Med. 2007, 11, 685–703. [Google Scholar] [CrossRef] [PubMed]
- Hidese, R.; Im, K.H.; Kobayashi, M.; Niitsu, M.; Furuchi, T.; Fujiwara, S. Identification of a novel acetylated form of branched-chain polyamine from a hyperthermophilic archaeon Thermococcus kodakarensis. Biosci. Biotechnol. Biochem. 2017, 81, 1845–1849. [Google Scholar] [CrossRef] [PubMed]
- Prunetti, L.; Graf, M.; Blaby, I.K.; Peil, L.; Makkay, A.M.; Starosta, A.L.; Papke, R.T.; Oshima, T.; Wilson, D.N.; de Crecy-Lagard, V. Deciphering the translation initiation factor 5A modification pathway in halophilic archaea. Archaea 2016, 2016, 7316725. [Google Scholar] [CrossRef] [PubMed]
- Hamana, K.; Hamana, H.; Itoh, T. Ubiquitous occurrence of agmatine as the major polyamine within extremely halophilic archaebacteria. J. Gen. Appl. Microbiol. 1995, 41, 153–158. [Google Scholar] [CrossRef]
- Ohnuma, M.; Ganbe, T.; Terui, Y.; Niitsu, M.; Sato, T.; Tanaka, N.; Tamakoshi, M.; Samejima, K.; Kumasaka, T.; Oshima, T. Crystal structures and enzymatic properties of a triamine/agmatine aminopropyltransferase from Thermus thermophilus. J. Mol. Biol. 2011, 408, 971–986. [Google Scholar] [CrossRef] [PubMed]
- Mattar, S.; Scharf, B.; Kent, S.B.; Rodewald, K.; Oesterhelt, D.; Engelhard, M. The primary structure of halocyanin, an archaeal blue copper protein, predicts a lipid anchor for membrane fixation. J. Biol. Chem. 1994, 269, 14939–14945. [Google Scholar] [PubMed]
- Scharf, B.; Wittenberg, R.; Engelhard, M. Electron transfer proteins from the haloalkaliphilic archaeon Natronobacterium pharaonis: Possible components of the respiratory chain include cytochrome bc and a terminal oxidase cytochrome ba3. Biochemistry 1997, 36, 4471–4479. [Google Scholar] [CrossRef] [PubMed]
- Giro, M.; Ceccoli, R.D.; Poli, H.O.; Carrillo, N.; Lodeyro, A.F. An in vivo system involving co-expression of cyanobacterial flavodoxin and ferredoxin-NADP+ reductase confers increased tolerance to oxidative stress in plants. FEBS Open Bio 2011, 1, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant. Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Ogawa, N.; Ihara, K.; Sugiyama, Y.; Mukohata, Y. Cytochrome aa3 in Haloferax volcanii. J. Bacteriol. 2002, 184, 840–845. [Google Scholar] [CrossRef] [PubMed]
- Lenart, A.; Dudkiewicz, M.; Grynberg, M.; Pawlowski, K. CLCAs—A family of metalloproteases of intriguing phylogenetic distribution and with cases of substituted catalytic sites. PLoS ONE 2013, 8, e62272. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.J.; Wholey, W.Y.; Parker, B.W.; Kim, M.; Jakob, U. NemR is a bleach-sensing transcription factor. J. Biol. Chem. 2013, 288, 13789–13798. [Google Scholar] [CrossRef] [PubMed]
- Jo-Watanabe, A.; Ohse, T.; Nishimatsu, H.; Takahashi, M.; Ikeda, Y.; Wada, T.; Shirakawa, J.; Nagai, R.; Miyata, T.; Nagano, T.; et al. Glyoxalase I reduces glycative and oxidative stress and prevents age-related endothelial dysfunction through modulation of endothelial nitric oxide synthase phosphorylation. Aging Cell. 2014, 13, 519–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamura, A.; Ohtsuka, J.; Kubota, K.; Agari, Y.; Ebihara, A.; Nakagawa, N.; Nagata, K.; Tanokura, M. Crystal structure of TTHA1429, a novel metallo-beta-lactamase superfamily protein from Thermus thermophilus HB8. Proteins 2008, 73, 1053–1057. [Google Scholar] [CrossRef] [PubMed]
- Altman-Price, N.; Mevarech, M. Genetic evidence for the importance of protein acetylation and protein deacetylation in the halophilic archaeon Haloferax volcanii. J. Bacteriol. 2009, 191, 1610–1617. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.D.; Botting, C.H.; Wardleworth, B.N.; Jackson, S.P.; White, M.F. The interaction of Alba, a conserved archaeal chromatin protein, with Sir2 and its regulation by acetylation. Science 2002, 296, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, Q.; Jiang, X.; Yang, H.; Zhao, D.; Han, J.; Luo, Y.; Xiang, H. Systematic analysis of lysine acetylation in the halophilic archaeon Haloferax mediterranei. J. Proteome Res. 2017, 16, 3229–3241. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Liu, R.; Sanchez, I.; Silva-Sanchez, C.; Hepowit, N.L.; Cao, S.; Chen, S.; Maupin-Furlow, J. Ubiquitin-like proteasome system represents a eukaryotic-like pathway for targeted proteolysis in archaea. mBio 2016, 7, e00379-16. [Google Scholar] [CrossRef] [PubMed]
- Maupin-Furlow, J.A. Archaeal proteasomes and sampylation. Subcell. Biochem. 2013, 66, 297–327. [Google Scholar] [CrossRef] [PubMed]
- Wilson, H.; Aldrich, H.; Maupin-Furlow, J. Halophilic 20S proteasomes of the archaeon Haloferax volcanii: Purification, characterization, and gene sequence analysis. J. Bacteriol. 1999, 181, 5814–5824. [Google Scholar] [PubMed]
- Reuter, C.; Kaczowka, S.; Maupin-Furlow, J. Differential regulation of the PanA and PanB proteasome-activating nucleotidase and 20S proteasomal proteins of the haloarchaeon Haloferax volcanii. J. Bacteriol. 2004, 186, 7763–7772. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Kowalczyk, D.; Humbard, M.; Rohatgi, S.; Maupin-Furlow, J. Proteasomal components required for cell growth and stress responses in the haloarchaeon Haloferax volcanii. J. Bacteriol. 2008, 190, 8096–8105. [Google Scholar] [CrossRef] [PubMed]
- Hanson, A.D.; Pribat, A.; Waller, J.C.; de Crecy-Lagard, V. ‘Unknown’ proteins and ‘orphan’ enzymes: The missing half of the engineering parts list—And how to find it. Biochem. J. 2009, 425, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ortiz de Orue Lucana, D.; Bogel, G.; Zou, P.; Groves, M.R. The oligomeric assembly of the novel haem-degrading protein HbpS is essential for interaction with its cognate two-component sensor kinase. J. Mol. Biol. 2009, 386, 1108–1122. [Google Scholar] [CrossRef] [PubMed]
- Omasits, U.; Ahrens, C.H.; Muller, S.; Wollscheid, B. Protter: Interactive protein feature visualization and integration with experimental proteomic data. Bioinformatics 2014, 30, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Delmas, S.; Shunburne, L.; Ngo, H.P.; Allers, T. Mre11-Rad50 promotes rapid repair of DNA damage in the polyploid archaeon Haloferax volcanii by restraining homologous recombination. PLoS Genet. 2009, 5, e1000552. [Google Scholar] [CrossRef] [PubMed]
- Wendoloski, D.; Ferrer, C.; Dyall-Smith, M.L. A new simvastatin (mevinolin)-resistance marker from Haloarcula hispanica and a new Haloferax volcanii strain cured of plasmid pHV2. Microbiology 2001, 147, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Allers, T.; Ngo, H.P.; Mevarech, M.; Lloyd, R.G. Development of additional selectable markers for the halophilic archaeon Haloferax volcanii based on the leuB and trpA genes. Appl. Environ. Microbiol. 2004, 70, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Miranda, H.; Nembhard, N.; Su, D.; Hepowit, N.; Krause, D.; Pritz, J.; Phillips, C.; Söll, D.; Maupin-Furlow, J. E1- and ubiquitin-like proteins provide a direct link between protein conjugation and sulfur transfer in archaea. Proc. Natl. Acad. Sci. USA 2011, 108, 4417–4422. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomez, M.; Leung, W.; Dantuluri, S.; Pillai, A.; Gani, Z.; Hwang, S.; McMillan, L.J.; Kiljunen, S.; Savilahti, H.; Maupin-Furlow, J.A. Molecular Factors of Hypochlorite Tolerance in the Hypersaline Archaeon Haloferax volcanii. Genes 2018, 9, 562. https://doi.org/10.3390/genes9110562
Gomez M, Leung W, Dantuluri S, Pillai A, Gani Z, Hwang S, McMillan LJ, Kiljunen S, Savilahti H, Maupin-Furlow JA. Molecular Factors of Hypochlorite Tolerance in the Hypersaline Archaeon Haloferax volcanii. Genes. 2018; 9(11):562. https://doi.org/10.3390/genes9110562
Chicago/Turabian StyleGomez, Miguel, Whinkie Leung, Swathi Dantuluri, Alexander Pillai, Zyan Gani, Sungmin Hwang, Lana J. McMillan, Saija Kiljunen, Harri Savilahti, and Julie A. Maupin-Furlow. 2018. "Molecular Factors of Hypochlorite Tolerance in the Hypersaline Archaeon Haloferax volcanii" Genes 9, no. 11: 562. https://doi.org/10.3390/genes9110562
APA StyleGomez, M., Leung, W., Dantuluri, S., Pillai, A., Gani, Z., Hwang, S., McMillan, L. J., Kiljunen, S., Savilahti, H., & Maupin-Furlow, J. A. (2018). Molecular Factors of Hypochlorite Tolerance in the Hypersaline Archaeon Haloferax volcanii. Genes, 9(11), 562. https://doi.org/10.3390/genes9110562