Role of the Extremolytes Ectoine and Hydroxyectoine as Stress Protectants and Nutrients: Genetics, Phylogenomics, Biochemistry, and Structural Analysis

 ,
, 
Abstract
:
1. Introduction
2. The Salt-In and Salt-Out Strategies for Coping with High Osmolarity Environments
3. Compatible Solutes
Stress-Relieving Cytoprotectants Used in All Three Domains of Life
4. Ectoine and Hydroxyectoine
4.1. Discovery
4.2. Physico-Chemical Attributes
4.3. Stress-Protective Properties
4.4. Biotechnological Production and Practical Applications of Ectoines
5. Ectoine/5-Hydroxyectoine Biosynthetic Routes and Crystal Structures of Selected Enzymes
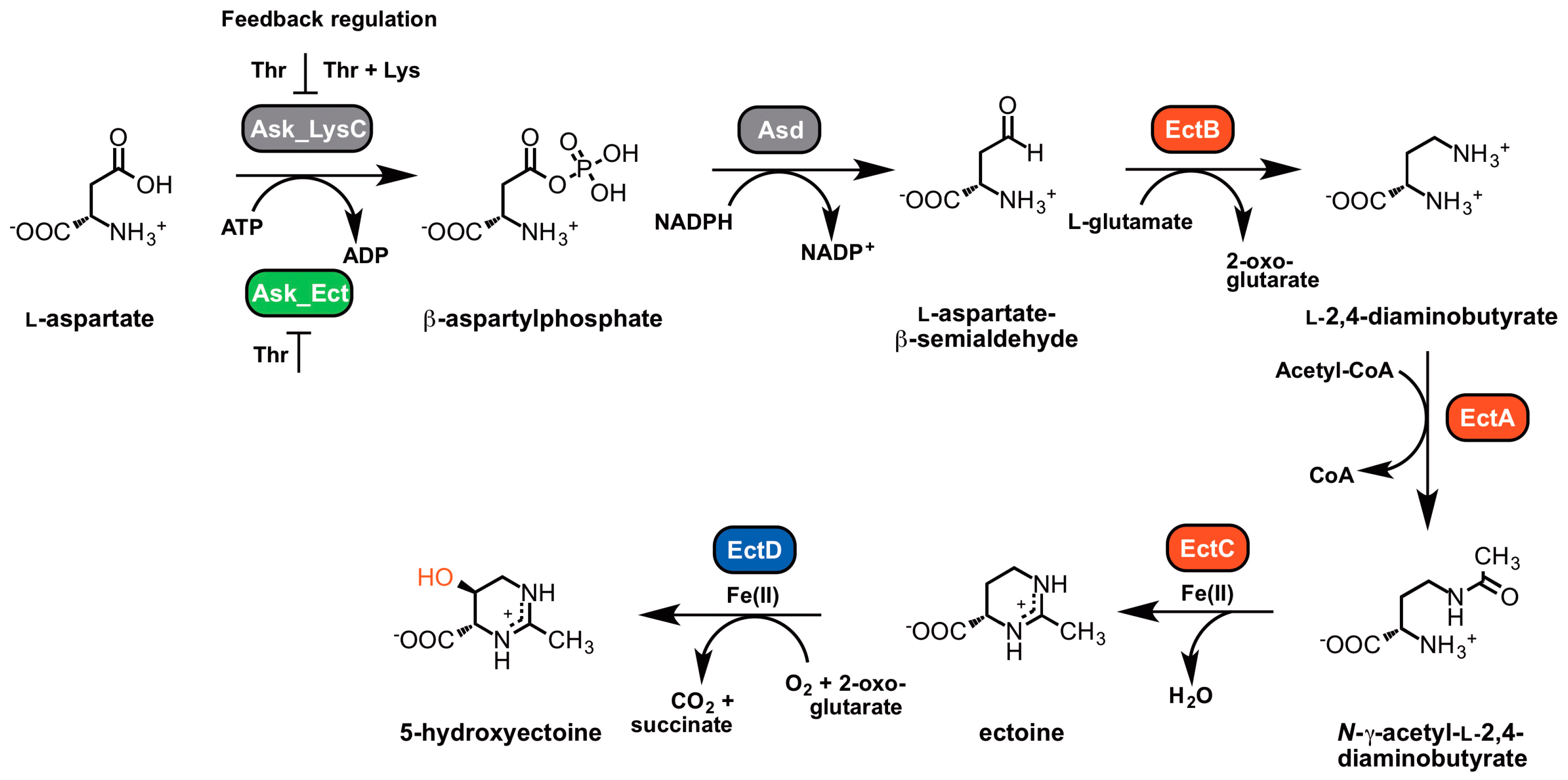
5.1. Biosynthetic Pathway: An Overview
5.2. Characteristics of the Ectoine/5-Hydroxyectoine Biosynthetic Enzymes
5.2.1. l-2,4-Diaminobutyrate Transaminase EctB
5.2.2. l-2,4-Diaminobutyrate Acetyltransferase EctA
5.2.3. Ectoine Synthase EctC
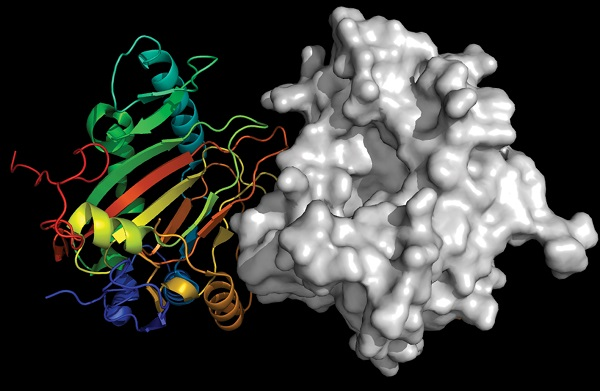
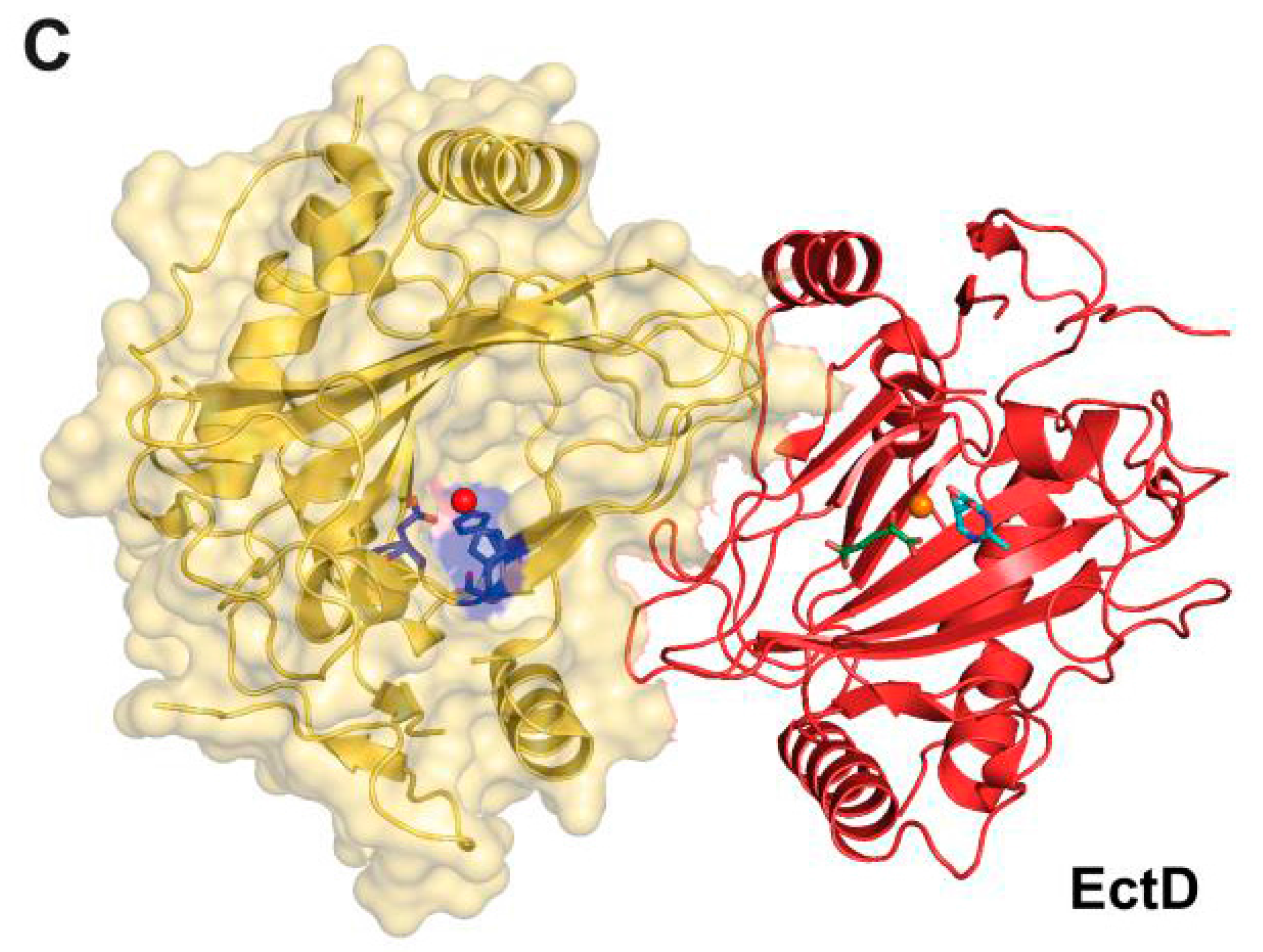
5.2.4. Ectoine Hydroxylase EctD
5.2.5. Specialized Aspartokinase Ask_Ect
5.2.6. Adjusting Central Carbon Metabolism to the Drain Exerted by Ectoine Biosynthesis
6. Genetics and Phylogenomics of Ectoine and 5-Hydroxyectoine Biosynthetic Genes
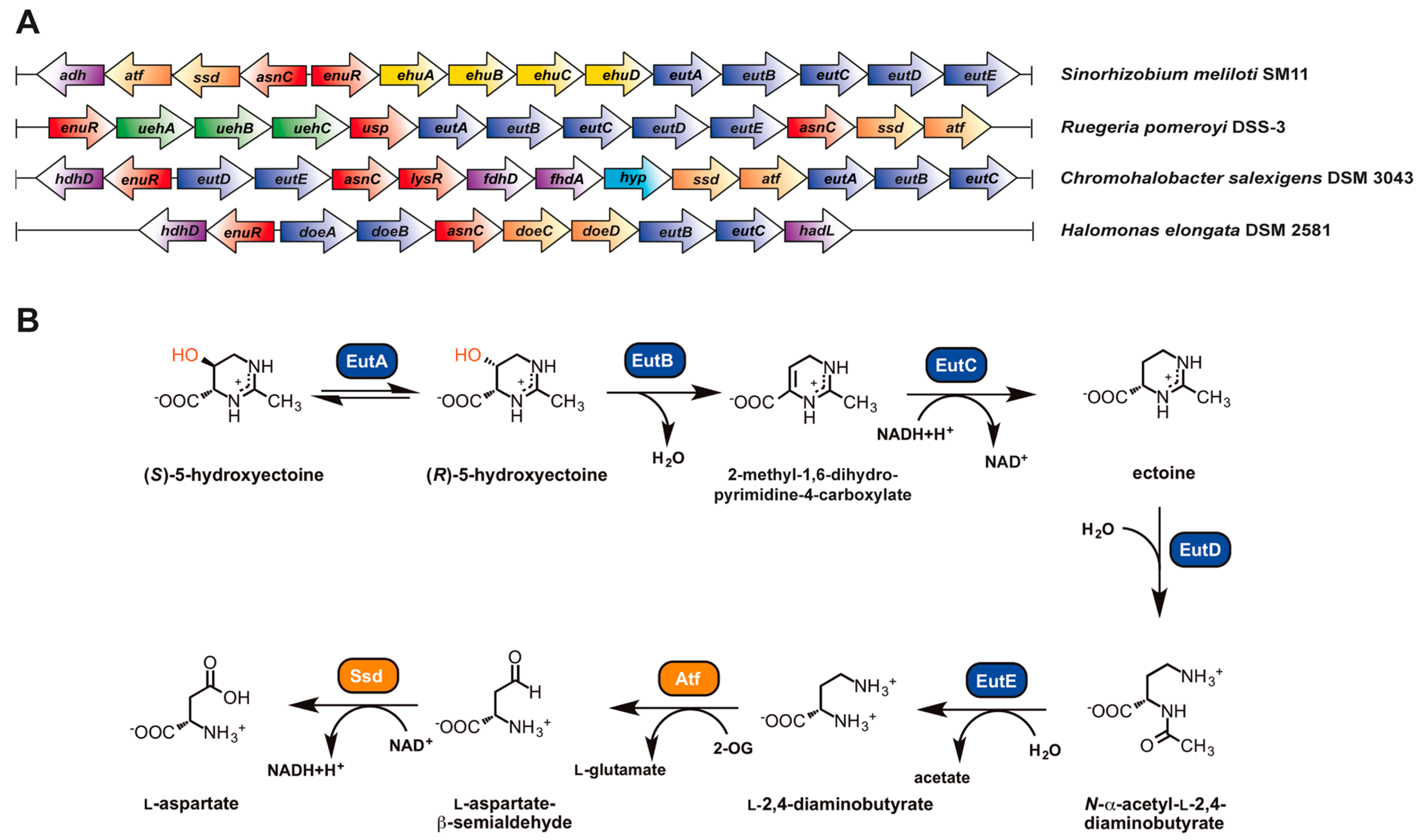
6.1. Genetic Organization of the Ectoine/5-Hydroxyectoine Biosynthetic Gene Clusters
6.2. Regulation of ect Gene Expression
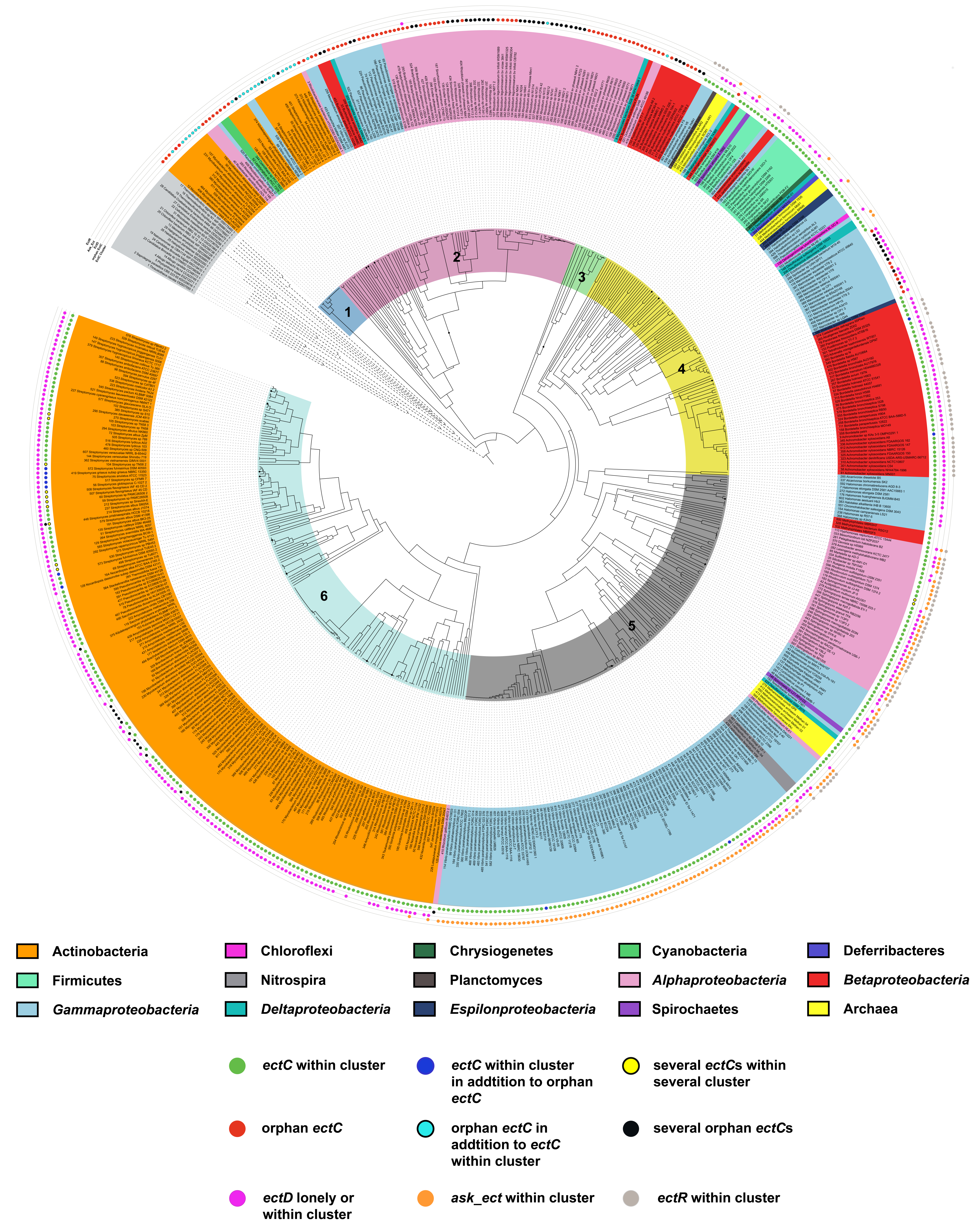
6.3. Phylogenomics of ect Genes
7. Scavenging Ectoines as Stress Protectants from Environmental Sources
8. Ectoines as Nutrients
8.1. Physiology
8.2. Genetics and Phylogenomics of Ectoine Catabolic Genes
8.3. Transporters for the Scavenging of Ectoines for Their Use as Nutrients
8.4. Biochemistry of Ectoine/5-Hydroxyectoine Catabolism
8.5. Genetic Regulation of Ectoine/5-Hydroxyectoine Catabolism
9. Ectoines in Eukarya: A Recent Discovery
10. Conclusions and Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kempf, B.; Bremer, E. Uptake and synthesis of compatible solutes as microbial stress responses to high osmolality environments. Arch. Microbiol. 1998, 170, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Bremer, E.; Krämer, R. Coping with osmotic challenges: Osmoregulation through accumulation and release of compatible solutes. In Bacterial Stress Responses; Storz, G., Hengge-Aronis, R., Eds.; ASM Press: Washington, DC, USA, 2000; pp. 79–97. [Google Scholar]
- Wood, J.M.; Bremer, E.; Csonka, L.N.; Krämer, R.; Poolman, B.; van der Heide, T.; Smith, L.T. Osmosensing and osmoregulatory compatible solute accumulation by bacteria. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 130, 437–460. [Google Scholar] [CrossRef]
- Csonka, L.N. Physiological and genetic responses of bacteria to osmotic stress. Microbiol. Rev. 1989, 53, 121–147. [Google Scholar] [PubMed]
- Roesser, M.; Müller, V. Osmoadaptation in bacteria and archaea: Common principles and differences. Environ. Microbiol. 2001, 3, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.M. Osmosensing by bacteria: Signals and membrane-based sensors. Microbiol. Mol. Biol. Rev. 1999, 63, 230–262. [Google Scholar] [CrossRef] [PubMed]
- Ball, P. Water is an active matrix of life for cell and molecular biology. Proc. Natl. Acad. Sci. USA 2017, 114, 13327–13335. [Google Scholar] [CrossRef] [PubMed]
- De Lima Alves, F.; Stevenson, A.; Baxter, E.; Gillion, J.L.; Hejazi, F.; Hayes, S.; Morrison, I.E.; Prior, B.A.; McGenity, T.J.; Rangel, D.E.; et al. Concomitant osmotic and chaotropicity-induced stresses in Aspergillus wentii: Compatible solutes determine the biotic window. Curr. Genet. 2015, 61, 457–477. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, J.; Boersma, A.J.; Poolman, B. Microorganisms maintain crowding homeostasis. Nat. Rev. Microbiol. 2017, 15, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.M. Bacterial osmoregulation: A paradigm for the study of cellular homeostasis. Annu. Rev. Microbiol. 2011, 65, 215–238. [Google Scholar] [CrossRef] [PubMed]
- Booth, I.R. Bacterial mechanosensitive channels: Progress towards an understanding of their roles in cell physiology. Curr. Opin. Microbiol. 2014, 18, 16–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whatmore, A.M.; Reed, R.H. Determination of turgor pressure in Bacillus subtilis: A possible role for K+ in turgor regulation. J. Gen. Microbiol. 1990, 136, 2521–2526. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Sun, M.; Shaevitz, J.W. Direct measurement of cell wall stress stiffening and turgor pressure in live bacterial cells. Phys. Rev. Lett. 2011, 107, 158101. [Google Scholar] [CrossRef] [PubMed]
- Cayley, D.S.; Guttman, H.J.; Record, M.T., Jr. Biophysical characterization of changes in amounts and activity of Escherichia coli cell and compartment water and turgor pressure in response to osmotic stress. Biophys. J. 2000, 78, 1748–1764. [Google Scholar] [CrossRef]
- Rojas, E.R.; Huang, K.C. Regulation of microbial growth by turgor pressure. Curr. Opin. Microbiol. 2017, 42, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Levina, N.; Totemeyer, S.; Stokes, N.R.; Louis, P.; Jones, M.A.; Booth, I.R. Protection of Escherichia coli cells against extreme turgor by activation of MscS and MscL mechanosensitive channels: Identification of genes required for mscs activity. EMBO J. 1999, 18, 1730–1737. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.; Boiangiu, C.; Moses, S.; Bremer, E. Responses of Bacillus subtilis to hypotonic challenges: Physiological contributions of mechanosensitive channels to cellular survival. Appl. Environ. Microbiol. 2008, 74, 2454–2460. [Google Scholar] [CrossRef] [PubMed]
- Cetiner, U.; Rowe, I.; Schams, A.; Mayhew, C.; Rubin, D.; Anishkin, A.; Sukharev, S. Tension-activated channels in the mechanism of osmotic fitness in Pseudomonas aeruginosa. J. Gen. Physiol. 2017, 149, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Reuter, M.; Hayward, N.J.; Black, S.S.; Miller, S.; Dryden, D.T.; Booth, I.R. Mechanosensitive channels and bacterial cell wall integrity: Does life end with a bang or a whimper? J. R. Soc. Interface 2014, 11, 20130850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, C.D.; Bavi, N.; Martinac, B. Bacterial mechanosensors. Annu. Rev. Physiol. 2018, 80, 71–93. [Google Scholar] [CrossRef] [PubMed]
- Calamita, G. The Escherichia coli aquaporin-Z water channel. Mol. Microbiol. 2000, 37, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Daniels, B.V.; Fu, D. Crystal structure of AqpZ tetramer reveals two distinct Arg-189 conformations associated with water permeation through the narrowest constriction of the water-conducting channel. J. Biol. Chem. 2006, 281, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Ventosa, A.; Nieto, J.J.; Oren, A. Biology of moderately halophilic aerobic bacteria. Microbiol. Mol. Biol. Rev. 1998, 62, 504–544. [Google Scholar] [PubMed]
- Oren, A. Life at high salt concentrations, intracellular KCl concentrations, and acidic proteomes. Front. Microbiol. 2013, 4, 315. [Google Scholar] [CrossRef] [PubMed]
- Galinski, E.A.; Trüper, H.G. Microbial behaviour in salt-stressed ecosystems. FEMS Microbiol. Rev. 1994, 15, 95–108. [Google Scholar] [CrossRef]
- Buda, R.; Liu, Y.; Yang, J.; Hegde, S.; Stevenson, K.; Bai, F.; Pilizota, T. Dynamics of Escherichia coli’s passive response to a sudden decrease in external osmolarity. Proc. Natl. Acad. Sci. USA 2016, 113, E5838–E5846. [Google Scholar] [CrossRef] [PubMed]
- Bialecka-Fornal, M.; Lee, H.J.; Phillips, R. The rate of osmotic downshock determines the survival probability of bacterial mechanosensitive channel mutants. J. Bacteriol. 2015, 197, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Egan, A.J.; Cleverley, R.M.; Peters, K.; Lewis, R.J.; Vollmer, W. Regulation of bacterial cell wall growth. FEBS J. 2017, 284, 851–867. [Google Scholar] [CrossRef] [PubMed]
- Typas, A.; Banzhaf, M.; Gross, C.A.; Vollmer, W. From the regulation of peptidoglycan synthesis to bacterial growth and morphology. Nat. Rev. Microbiol. 2012, 10, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, T. How do mechanosensitive channels sense membrane tension? Biochem. Soc. Trans. 2016, 44, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Booth, I.R.; Blount, P. The MscS and MscL families of mechanosensitive channels act as microbial emergency release valves. J. Bacteriol. 2012, 194, 4802–4809. [Google Scholar] [CrossRef] [PubMed]
- Pliotas, C.; Naismith, J.H. Spectator no more, the role of the membrane in regulating ion channel function. Curr. Opin. Struct. Biol. 2017, 45, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Coquelle, N.; Talon, R.; Juers, D.H.; Girard, E.; Kahn, R.; Madern, D. Gradual adaptive changes of a protein facing high salt concentrations. J. Mol. Biol. 2010, 404, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Talon, R.; Coquelle, N.; Madern, D.; Girard, E. An experimental point of view on hydration/solvation in halophilic proteins. Front. Microbiol. 2014, 5, 66. [Google Scholar] [CrossRef] [PubMed]
- Tadeo, X.; Lopez-Mendez, B.; Trigueros, T.; Lain, A.; Castano, D.; Millet, O. Structural basis for the aminoacid composition of proteins from halophilic archea. PLoS Biol. 2009, 7, e1000257. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Thermodynamic limits to microbial life at high salt concentrations. Environ. Microbiol. 2011, 13, 1908–1923. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Bioenergetic aspects of halophilism. Microbiol. Mol. Biol. Rev. 1999, 63, 334–348. [Google Scholar] [PubMed]
- Youssef, N.H.; Savage-Ashlock, K.N.; McCully, A.L.; Luedtke, B.; Shaw, E.I.; Hoff, W.D.; Elshahed, M.S. Trehalose/2-sulfotrehalose biosynthesis and glycine-betaine uptake are widely spread mechanisms for osmoadaptation in the Halobacteriales. ISME J. 2014, 8, 636–649. [Google Scholar] [CrossRef] [PubMed]
- Deole, R.; Challacombe, J.; Raiford, D.W.; Hoff, W.D. An extremely halophilic proteobacterium combines a highly acidic proteome with a low cytoplasmic potassium content. J. Biol. Chem. 2013, 288, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Kokoeva, M.V.; Storch, K.F.; Klein, C.; Oesterhelt, D. A novel mode of sensory transduction in archaea: Binding protein-mediated chemotaxis towards osmoprotectants and amino acids. EMBO J. 2002, 21, 2312–2322. [Google Scholar] [CrossRef] [PubMed]
- Becker, E.A.; Seitzer, P.M.; Tritt, A.; Larsen, D.; Krusor, M.; Yao, A.I.; Wu, D.; Madern, D.; Eisen, J.A.; Darling, A.E.; et al. Phylogenetically driven sequencing of extremely halophilic archaea reveals strategies for static and dynamic osmo-response. PLoS Genet. 2014, 10, e1004784. [Google Scholar] [CrossRef] [PubMed]
- Lippert, K.; Galinski, E.A.; Truper, H.G. Biosynthesis and function of trehalose in Ectothiorhodospira halochloris. Antonie Leeuwenhoek 1993, 63, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, S.; Dev, K.; Sourirajan, A. Distinct osmoadaptation strategies in the strict halophilic and halotolerant bacteria isolated from Lunsu salt water body of North West Himalayas. Curr. Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Yancey, P.H.; Clark, M.E.; Hand, S.C.; Bowlus, R.D.; Somero, G.N. Living with water stress: Evolution of osmolyte systems. Science 1982, 217, 1214–1222. [Google Scholar] [CrossRef] [PubMed]
- Burg, M.B.; Ferraris, J.D. Intracellular organic osmolytes: Function and regulation. J. Biol. Chem. 2008, 283, 7309–7313. [Google Scholar] [CrossRef] [PubMed]
- Gunde-Cimerman, N.; Plemenitas, A.; Oren, A. Strategies of adaptation of microorganisms of the three domains of life to high-salt concentrations. FEMS Microbiol. Rev. 2018, in press. [Google Scholar] [CrossRef] [PubMed]
- Le Rudulier, D.; Strom, A.R.; Dandekar, A.M.; Smith, L.T.; Valentine, R.C. Molecular biology of osmoregulation. Science 1984, 224, 1064–1068. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.D. Microbial water stress. Bacteriol. Rev. 1976, 40, 803–846. [Google Scholar] [PubMed]
- Gilles, R. “Compensatory” organic osmolytes in high osmolarity and dehydration stresses: History and perspectives. Comp. Biochem. Physiol. A Physiol. 1997, 117, 279–290. [Google Scholar] [CrossRef]
- Yancey, P.H. Organic osmolytes as compatible, metabolic and counteracting cytoprotectants in high osmolarity and other stresses. J. Exp. Biol. 2005, 208, 2819–2830. [Google Scholar] [CrossRef] [PubMed]
- Holtmann, G.; Bremer, E. Thermoprotection of Bacillus subtilis by exogenously provided glycine betaine and structurally related compatible solutes: Involvement of opu transporters. J. Bacteriol. 2004, 186, 1683–1693. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.; Bremer, E. Protection of Bacillus subtilis against cold stress via compatible-solute acquisition. J. Bacteriol. 2011, 193, 1552–1562. [Google Scholar] [CrossRef] [PubMed]
- Caldas, T.; Demont-Caulet, N.; Ghazi, A.; Richarme, G. Thermoprotection by glycine betaine and choline. Microbiology 1999, 145, 2543–2548. [Google Scholar] [CrossRef] [PubMed]
- Diamant, S.; Eliahu, N.; Rosenthal, D.; Goloubinoff, P. Chemical chaperones regulate molecular chaperones in vitro and in cells under combined salt and heat stresses. J. Biol. Chem. 2001, 276, 39586–39591. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, M.S.; Santos, H.; Galinski, E.A. An overview of the role and diversity of compatible solutes in Bacteria and Archaea. Adv. Biochem. Eng. Biotechnol. 1998, 61, 117–153. [Google Scholar] [PubMed]
- Martins, L.O.; Huber, R.; Huber, H.; Stetter, K.O.; da Costa, M.S.; Santos, H. Organic solutes in hyperthermophilic archaea. Appl. Environ. Microbiol. 1997, 63, 896–902. [Google Scholar] [PubMed]
- Santos, H.; da Costa, M.S. Compatible solutes of organisms that live in hot saline environments. Environ. Microbiol. 2002, 4, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Yancey, P.H. Compatible and counteracting solutes: Protecting cells from the Dead Sea to the deep sea. Sci. Prog. 2004, 87, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, T.; Timasheff, S.N. The stabilization of proteins by osmolytes. Biophys. J. 1985, 47, 411–414. [Google Scholar] [CrossRef]
- Auton, M.; Rösgen, J.; Sinev, M.; Holthauzen, L.M.; Bolen, D.W. Osmolyte effects on protein stability and solubility: A balancing act between backbone and side-chains. Biophys. Chem. 2011, 159, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Bolen, D.W.; Baskakov, I.V. The osmophobic effect: Natural selection of a thermodynamic force in protein folding. J. Mol. Biol. 2001, 310, 955–963. [Google Scholar] [CrossRef] [PubMed]
- Street, T.O.; Bolen, D.W.; Rose, G.D. A molecular mechanism for osmolyte-induced protein stability. Proc. Natl. Acad. Sci. USA 2006, 103, 13997–14002. [Google Scholar] [CrossRef] [PubMed]
- Bourot, S.; Sire, O.; Trautwetter, A.; Touze, T.; Wu, L.F.; Blanco, C.; Bernard, T. Glycine betaine-assisted protein folding in a lysA mutant of Escherichia coli. J. Biol. Chem. 2000, 275, 1050–1056. [Google Scholar] [CrossRef] [PubMed]
- Ignatova, Z.; Gierasch, L.M. Inhibition of protein aggregation in vitro and in vivo by a natural osmoprotectant. Proc. Natl. Acad. Sci. USA 2006, 103, 13357–13361. [Google Scholar] [CrossRef] [PubMed]
- Stadmiller, S.S.; Gorensek-Benitez, A.H.; Guseman, A.J.; Pielak, G.J. Osmotic shock induced protein destabilization in living cells and its reversal by glycine betaine. J. Mol. Biol. 2017, 429, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Harishchandra, R.K.; Wulff, S.; Lentzen, G.; Neuhaus, T.; Galla, H.J. The effect of compatible solute ectoines on the structural organization of lipid monolayer and bilayer membranes. Biophys. Chem. 2010, 150, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Smiatek, J.; Harishchandra, R.K.; Galla, H.J.; Heuer, A. Low concentrated hydroxyectoine solutions in presence of DPPC lipid bilayers: A computer simulation study. Biophys. Chem. 2013, 180–181, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Record, M.T., Jr.; Courtenay, E.S.; Cayley, S.; Guttman, H.J. Biophysical compensation mechanisms buffering E. coli protein-nucleic acid interactions against changing environments. Trends Biochem. Sci. 1998, 23, 190–194. [Google Scholar] [CrossRef]
- Record, M.T., Jr.; Courtenay, E.S.; Cayley, D.S.; Guttman, H.J. Responses of E. coli to osmotic stress: Large changes in amounts of cytoplasmic solutes and water. Trends Biochem. Sci. 1998, 23, 143–148. [Google Scholar] [CrossRef]
- Tatzelt, J.; Prusiner, S.B.; Welch, W.J. Chemical chaperones interfere with the formation of scrapie prion protein. EMBO J. 1996, 15, 6363–6373. [Google Scholar] [PubMed]
- Kolp, S.; Pietsch, M.; Galinski, E.A.; Gutschow, M. Compatible solutes as protectants for zymogens against proteolysis. Biochim. Biophys. Acta 2006, 1764, 1234–1242. [Google Scholar] [CrossRef] [PubMed]
- Lippert, K.; Galinski, E.A. Enzyme stabilization by ectoine-type compatible solutes: Protection against heating, freezing and drying. Appl. Micro Biotechnol. 1992, 37, 61–65. [Google Scholar] [CrossRef]
- Knapp, S.; Ladenstein, R.; Galinski, E.A. Extrinsic protein stabilization by the naturally occurring osmolytes beta-hydroxyectoine and betaine. Extremophiles 1999, 3, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Manzanera, M.; Garcia de Castro, A.; Tondervik, A.; Rayner-Brandes, M.; Strom, A.R.; Tunnacliffe, A. Hydroxyectoine is superior to trehalose for anhydrobiotic engineering of Pseudomonas putida KT2440. Appl. Environ. Microbiol. 2002, 68, 4328–4333. [Google Scholar] [CrossRef] [PubMed]
- Manzanera, M.; Vilchez, S.; Tunnacliffe, A. High survival and stability rates of Escherichia coli dried in hydroxyectoine. FEMS Microbiol. Lett. 2004, 233, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, M.K.; Kern, R.; Mistou, M.Y.; Dandekar, A.M.; Uratsu, S.L.; Richarme, G. The chemical chaperone proline relieves the thermosensitivity of a dnaK deletion mutant at 42 °C. J. Bacteriol. 2004, 186, 8149–8152. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmann, A.U.; Bursy, J.; Gimpel, S.; Hoffmann, T.; Bremer, E. Synthesis of the compatible solute ectoine in Virgibacillus pantothenticus is triggered by high salinity and low growth temperature. Appl. Environ. Microbiol. 2008, 74, 4560–4563. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmann, A.U.; Hoffmann, T.; Bursy, J.; Jebbar, M.; Bremer, E. Ectoine and hydroxyectoine as protectants against osmotic and cold stress: Uptake through the SigB-controlled betaine-choline- carnitine transporter-type carrier EctT from Virgibacillus pantothenticus. J. Bacteriol. 2011, 193, 4699–4708. [Google Scholar] [CrossRef] [PubMed]
- Bursy, J.; Kuhlmann, A.U.; Pittelkow, M.; Hartmann, H.; Jebbar, M.; Pierik, A.J.; Bremer, E. Synthesis and uptake of the compatible solutes ectoine and 5-hydroxyectoine by Streptomyces coelicolor A3(2) in response to salt and heat stresses. Appl. Environ. Microbiol. 2008, 74, 7286–7296. [Google Scholar] [CrossRef] [PubMed]
- Vargas, C.; Argandona, M.; Reina-Bueno, M.; Rodriguez-Moya, J.; Fernandez-Aunion, C.; Nieto, J.J. Unravelling the adaptation responses to osmotic and temperature stress in Chromohalobacter salexigens, a bacterium with broad salinity tolerance. Saline Syst. 2008, 4, 14. [Google Scholar] [CrossRef] [PubMed]
- Malin, G.; Lapidot, A. Induction of synthesis of tetrahydropyrimidine derivatives in Streptomyces strains and their effect on Escherichia coli in response to osmotic and heat stress. J. Bacteriol. 1996, 178, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Tschapek, B.; Pittelkow, M.; Sohn-Bosser, L.; Holtmann, G.; Smits, S.H.; Gohlke, H.; Bremer, E.; Schmitt, L. Arg149 is involved in switching the low affinity, open state of the binding protein AfProX into its high affinity, closed state. J. Mol. Biol. 2011, 411, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Lamosa, P.; Burke, A.; Peist, R.; Huber, R.; Liu, M.Y.; Silva, G.; Rodrigues-Pousada, C.; LeGall, J.; Maycock, C.; Santos, H. Thermostabilization of proteins by diglycerol phosphate, a new compatible solute from the hyperthermophile Archaeoglobus fulgidus. Appl. Environ. Microbiol. 2000, 66, 1974–1979. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.; Bremer, E. Management of osmotic stress by Bacillus subtilis: Genetics and physiology. In Stress and Environmental Regulation of Gene Expression and Adaptation in Bacteria; de Bruijn, F.J., Ed.; Wiley-Blackwell Publishers: Hoboken, NJ, USA, 2016; Volume 1, pp. 657–676. [Google Scholar]
- Hoffmann, T.; Bremer, E. Guardians in a stressful world: The Opu family of compatible solute transporters from Bacillus subtilis. Biol. Chem. 2017, 398, 193–214. [Google Scholar] [CrossRef] [PubMed]
- Lentzen, G.; Schwarz, T. Extremolytes: Natural compounds from extremophiles for versatile applications. Appl. Microbiol. Biotechnol. 2006, 72, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.F. Osmoadaptation and osmoregulation in archaea: Update 2004. Front. BioSci. 2004, 9, 1999–2019. [Google Scholar] [CrossRef] [PubMed]
- Galinski, E.A.; Pfeiffer, H.P.; Trüper, H.G. 1,4,5,6-tetrahydro-2-methyl-4-pyrimidinecarboxylic acid. A novel cyclic amino acid from halophilic phototrophic bacteria of the genus Ectothiorhodospira. Eur. J. Biochem. 1985, 149, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Inbar, L.; Lapidot, A. The structure and biosynthesis of new tetrahydropyrimidine derivatives in actinomycin D producer Streptomyces parvulus. Use of 13C- and 15N-labeled l-glutamate and 13C and 15N NMR spectroscopy. J. Biol. Chem. 1988, 263, 16014–16022. [Google Scholar] [PubMed]
- Inbar, L.; Frolow, F.; Lapidot, A. The conformation of new tetrahydropyrimidine derivatives in solution and in the crystal. Eur. J. Biochem. 1993, 214, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Pastor, J.M.; Salvador, M.; Argandona, M.; Bernal, V.; Reina-Bueno, M.; Csonka, L.N.; Iborra, J.L.; Vargas, C.; Nieto, J.J.; Canovas, M. Ectoines in cell stress protection: Uses and biotechnological production. Biotechnol. Adv. 2010, 28, 782–801. [Google Scholar] [CrossRef] [PubMed]
- Widderich, N.; Czech, L.; Elling, F.J.; Könneke, M.; Stöveken, N.; Pittelkow, M.; Riclea, R.; Dickschat, J.S.; Heider, J.; Bremer, E. Strangers in the archaeal world: Osmostress-responsive biosynthesis of ectoine and hydroxyectoine by the marine thaumarchaeon Nitrosopumilus maritimus. Environ. Microbiol. 2016, 18, 1227–1248. [Google Scholar] [CrossRef] [PubMed]
- Widderich, N.; Höppner, A.; Pittelkow, M.; Heider, J.; Smits, S.H.; Bremer, E. Biochemical properties of ectoine hydroxylases from extremophiles and their wider taxonomic distribution among microorganisms. PLoS ONE 2014, 9, e93809. [Google Scholar] [CrossRef] [PubMed]
- Kunte, H.J.; Lentzen, G.; Galinski, E. Industrial production of the cell protectant ectoine: Protection, mechanisms, processes, and products. Curr. Biotechnol. 2014, 3, 10–25. [Google Scholar] [CrossRef]
- Louis, P.; Galinski, E.A. Characterization of genes for the biosynthesis of the compatible solute ectoine from Marinococcus halophilus and osmoregulated expression in Escherichia coli. Microbiology 1997, 143, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, J.; Schauwecker, F.; Grammel, N.; Keller, U.; Bernhard, M. Functional expression of the ectoine hydroxylase gene (thpD) from Streptomyces chrysomallus in Halomonas elongata. Appl. Environ. Microbiol. 2004, 70, 3130–3132. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Estepa, R.; Argandona, M.; Reina-Bueno, M.; Capote, N.; Iglesias-Guerra, F.; Nieto, J.J.; Vargas, C. The ectD gene, which is involved in the synthesis of the compatible solute hydroxyectoine, is essential for thermoprotection of the halophilic bacterium Chromohalobacter salexigens. J. Bacteriol. 2006, 188, 3774–3784. [Google Scholar] [CrossRef] [PubMed]
- Bursy, J.; Pierik, A.J.; Pica, N.; Bremer, E. Osmotically induced synthesis of the compatible solute hydroxyectoine is mediated by an evolutionarily conserved ectoine hydroxylase. J. Biol. Chem. 2007, 282, 31147–31155. [Google Scholar] [CrossRef] [PubMed]
- Reshetnikov, A.S.; Khmelenina, V.N.; Mustakhimov, I.I.; Kalyuzhnaya, M.; Lidstrom, M.; Trotsenko, Y.A. Diversity and phylogeny of the ectoine biosynthesis genes in aerobic, moderately halophilic methylotrophic bacteria. Extremophiles 2011, 15, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Harding, T.; Roger, A.J.; Simpson, A.G.B. Adaptations to high salt in a halophilic protist: Differential expression and gene acquisitions through duplications and gene transfers. Front. Microbiol. 2017, 8, 944. [Google Scholar] [CrossRef] [PubMed]
- Harding, T.; Brown, M.W.; Simpson, A.G.; Roger, A.J. Osmoadaptative strategy and its molecular signature in obligately halophilic heterotrophic protists. Genome Biol. Evol. 2016, 8, 2241–2258. [Google Scholar] [CrossRef] [PubMed]
- Weinisch, L.; Kuhner, S.; Roth, R.; Grimm, M.; Roth, T.; Netz, D.J.A.; Pierik, A.J.; Filker, S. Identification of osmoadaptive strategies in the halophile, heterotrophic ciliate Schmidingerothrix salinarum. PLoS Biol. 2018, 16, e2003892. [Google Scholar] [CrossRef] [PubMed]
- Harding, T.; Simpson, A.G.B. Recent advances in halophilic protozoa research. J. Eukaryot. Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Czech, L.; Bremer, E. With a pinch of extra salt—Did predatory protists steal genes from their food? PLoS Biol. 2018, 16, e2005163. [Google Scholar] [CrossRef] [PubMed]
- Zaccai, G.; Bagyan, I.; Combet, J.; Cuello, G.J.; Deme, B.; Fichou, Y.; Gallat, F.X.; Galvan Josa, V.M.; von Gronau, S.; Haertlein, M.; et al. Neutrons describe ectoine effects on water H-bonding and hydration around a soluble protein and a cell membrane. Sci. Rep. 2016, 6, 31434. [Google Scholar] [CrossRef] [PubMed]
- Dötsch, A.; Severin, J.; Alt, W.; Galinski, E.A.; Kreft, J.U. A mathematical model for growth and osmoregulation in halophilic bacteria. Microbiology 2008, 154, 2956–2969. [Google Scholar] [CrossRef] [PubMed]
- Held, C.; Neuhaus, T.; Sadowski, G. Compatible solutes: Thermodynamic properties and biological impact of ectoines and prolines. Biophys. Chem. 2010, 152, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Eiberweiser, A.; Nazet, A.; Kruchinin, S.E.; Fedotova, M.V.; Buchner, R. Hydration and ion binding of the osmolyte ectoine. J. Phys. Chem. B 2015, 119, 15203–15211. [Google Scholar] [CrossRef] [PubMed]
- Hahn, M.B.; Solomun, T.; Wellhausen, R.; Hermann, S.; Seitz, H.; Meyer, S.; Kunte, H.J.; Zeman, J.; Uhlig, F.; Smiatek, J.; et al. Influence of the compatible solute ectoine on the local water structure: Implications for the binding of the protein G5P to DNA. J. Phys. Chem. B 2015, 119, 15212–15220. [Google Scholar] [CrossRef] [PubMed]
- Smiatek, J. Osmolyte effects: Impact on the aqueous solution around charged and neutral spheres. J. Phys. Chem. B 2014, 118, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Smiatek, J.; Harishchandra, R.K.; Rubner, O.; Galla, H.J.; Heuer, A. Properties of compatible solutes in aqueous solution. Biophys. Chem. 2012, 160, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Malin, G.; Iakobashvili, R.; Lapidot, A. Effect of tetrahydropyrimidine derivatives on protein-nucleic acids interaction. Type II restriction endonucleases as a model system. J. Biol. Chem. 1999, 274, 6920–6929. [Google Scholar] [CrossRef] [PubMed]
- Lapidot, A.; Ben-Asher, E.; Eisenstein, M. Tetrahydropyrimidine derivatives inhibit binding of a Tat-like, arginine-containing peptide, to HIV TAR RNA in vitro. FEBS Lett. 1995, 367, 33–38. [Google Scholar] [CrossRef]
- Kurz, M. Compatible solute influence on nucleic acids: Many questions but few answers. Saline Syst. 2008, 4, 6. [Google Scholar] [CrossRef] [PubMed]
- Barth, S.; Huhn, M.; Matthey, B.; Klimka, A.; Galinski, E.A.; Engert, A. Compatible-solute-supported periplasmic expression of functional recombinant proteins under stress conditions. Appl. Environ. Microbiol. 2000, 66, 1572–1579. [Google Scholar] [CrossRef] [PubMed]
- Czech, L.; Poehl, S.; Hub, P.; Stoeveken, N.; Bremer, E. Tinkering with osmotically controlled transcription allows enhanced production and excretion of ectoine and hydroxyectoine from a microbial cell factory. Appl. Environ. Microbiol. 2108, 84, e01772-17. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmann, A.U.; Bremer, E. Osmotically regulated synthesis of the compatible solute ectoine in Bacillus pasteurii and related Bacillus spp. Appl. Environ. Microbiol. 2002, 68, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Calderon, M.I.; Vargas, C.; Rojo, F.; Iglesias-Guerra, F.; Csonka, L.N.; Ventosa, A.; Nieto, J.J. Complex regulation of the synthesis of the compatible solute ectoine in the halophilic bacterium Chromohalobacter salexigens DSM 3043T. Microbiology 2004, 150, 3051–3063. [Google Scholar] [CrossRef] [PubMed]
- Göller, K.; Ofer, A.; Galinski, E.A. Construction and characterization of an NaCl sensitive mutant of Halomonas elongata impaired in ectoine biosynthesis. FEMS Microbiol. Lett. 1998, 161, 293–300. [Google Scholar] [CrossRef]
- Canovas, D.; Vargas, C.; Iglesias-Guerra, F.; Csonka, L.N.; Rhodes, D.; Ventosa, A.; Nieto, J.J. Isolation and characterization of salt-sensitive mutants of the moderate halophile Halomonas elongata and cloning of the ectoine synthesis genes. J. Biol. Chem. 1997, 272, 25794–25801. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Wang, Q.; Xu, W.; Liu, X.; Gao, X.; Zhang, Y. Stationary phase-dependent accumulation of ectoine is an efficient adaptation strategy in Vibrio anguillarum against cold stress. Microbiol. Res. 2017, 205, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Stöveken, N.; Pittelkow, M.; Sinner, T.; Jensen, R.A.; Heider, J.; Bremer, E. A specialized aspartokinase enhances the biosynthesis of the osmoprotectants ectoine and hydroxyectoine in Pseudomonas stutzeri A1501. J. Bacteriol. 2011, 193, 4456–4468. [Google Scholar] [CrossRef] [PubMed]
- Seip, B.; Galinski, E.A.; Kurz, M. Natural and engineered hydroxyectoine production based on the Pseudomonas stutzeri ectABCD-ask gene cluster. Appl. Environ. Microbiol. 2011, 77, 1368–1374. [Google Scholar] [CrossRef] [PubMed]
- Saum, S.H.; Müller, V. Salinity-dependent switching of osmolyte strategies in a moderately halophilic bacterium: Glutamate induces proline biosynthesis in Halobacillus halophilus. J. Bacteriol. 2007, 189, 6968–6975. [Google Scholar] [CrossRef] [PubMed]
- Saum, S.H.; Müller, V. Regulation of osmoadaptation in the moderate halophile Halobacillus halophilus: Chloride, glutamate and switching osmolyte strategies. Saline Syst. 2008, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Guinn, E.J.; Buechel, E.; Wong, R.; Sengupta, R.; Shkel, I.A.; Record, M.T. Basis of protein stabilization by K glutamate: Unfavorable interactions with carbon, oxygen groups. Biophys. J. 2016, 111, 1854–1865. [Google Scholar] [CrossRef] [PubMed]
- Diehl, R.C.; Guinn, E.J.; Capp, M.W.; Tsodikov, O.V.; Record, M.T., Jr. Quantifying additive interactions of the osmolyte proline with individual functional groups of proteins: Comparisons with urea and glycine betaine, interpretation of m-values. Biochemistry 2013, 52, 5997–6010. [Google Scholar] [CrossRef] [PubMed]
- Cayley, S.; Lewis, B.A.; Record, M.T., Jr. Origins of the osmoprotective properties of betaine and proline in Escherichia coli K-12. J. Bacteriol. 1992, 174, 1586–1595. [Google Scholar] [CrossRef] [PubMed]
- Tao, P.; Li, H.; Yu, Y.; Gu, J.; Liu, Y. Ectoine and 5-hydroxyectoine accumulation in the halophile Virgibacillus halodenitrificans PDB-F2 in response to salt stress. Appl. Microbiol. Biotechnol. 2016, 100, 6779–6789. [Google Scholar] [CrossRef] [PubMed]
- Klauck, E.; Typas, A.; Hengge, R. The sigmaS subunit of RNA polymerase as a signal integrator and network master regulator in the general stress response in Escherichia coli. Sci. Prog. 2007, 90, 103–127. [Google Scholar] [PubMed]
- Hengge-Aronis, R. Back to log phase: Sigma S as a global regulator in the osmotic control of gene expression in escherichia coli. Mol. Microbiol. 1996, 21, 887–893. [Google Scholar] [CrossRef] [PubMed]
- Tanne, C.; Golovina, E.A.; Hoekstra, F.A.; Meffert, A.; Galinski, E.A. Glass-forming property of hydroxyectoine is the cause of its superior function as a dessication protectant. Front. Microbiol. 2014, 5, 150. [Google Scholar] [CrossRef] [PubMed]
- Borges, N.; Ramos, A.; Raven, N.D.; Sharp, R.J.; Santos, H. Comparative study of the thermostabilizing properties of mannosylglycerate and other compatible solutes on model enzymes. Extremophiles 2002, 6, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Van-Thuoc, D.; Hashim, S.O.; Hatti-Kaul, R.; Mamo, G. Ectoine-mediated protection of enzyme from the effect of pH and temperature stress: A study using Bacillus halodurans xylanase as a model. Appl. Microbiol. Biotechnol. 2013, 97, 6271–6278. [Google Scholar] [CrossRef] [PubMed]
- Argandona, M.; Nieto, J.J.; Iglesias-Guerra, F.; Calderon, M.I.; Garcia-Estepa, R.; Vargas, C. Interplay between iron homeostasis and the osmotic stress response in the halophilic bacterium Chromohalobacter salexigens. Appl. Environ. Microbiol. 2010, 76, 3575–3589. [Google Scholar] [CrossRef] [PubMed]
- Scoma, A.; Boon, N. Osmotic stress confers enhanced cell integrity to hydrostatic pressure but impairs growth in Alcanivorax borkumensis SK2. Front. Microbiol. 2016, 7, 729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kish, A.; Griffin, P.L.; Rogers, K.L.; Fogel, M.L.; Hemley, R.J.; Steele, A. High-pressure tolerance in Halobacterium salinarum NCR-1 and other non-piezophilic prokaryotes. Extremophiles 2012, 16, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Schröter, M.A.; Meyer, S.; Hahn, M.B.; Solomun, T.; Sturm, H.; Kunte, H.J. Ectoine protects DNA from damage by ionizing radiation. Sci. Rep. 2017, 7, 15272. [Google Scholar] [CrossRef] [PubMed]
- Hahn, M.B.; Meyer, S.; Schröter, M.A.; Kunte, H.J.; Solomun, T.; Sturm, H. DNA protection by ectoine from ionizing radiation: Molecular mechanisms. Phys. Chem. Chem. Phys. 2017, 19, 25717–25722. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.; Schröter, M.A.; Hahn, M.B.; Solomun, T.; Sturm, H.; Kunte, H.J. Ectoine can enhance structural changes in DNA in vitro. Sci. Rep. 2017, 7, 7170. [Google Scholar] [CrossRef] [PubMed]
- Buenger, J.; Driller, H. Ectoine: An effective natural substance to prevent UVA-induced premature photoaging. Skin Pharmacol. Physiol. 2004, 17, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Bünger, J.; Degwert, J.; Driller, H. The protective function of compatible solute ectoine on skin cells and its biomolecules with respect to uv-radiation, immunosuppression and membrane damage. IFSCC Mag. 2001, 4, 1–6. [Google Scholar]
- Schnoor, M.; Voss, P.; Cullen, P.; Boking, T.; Galla, H.J.; Galinski, E.A.; Lorkowski, S. Characterization of the synthetic compatible solute homoectoine as a potent PCR enhancer. Biochem. Biophys. Res. Commun. 2004, 322, 867–872. [Google Scholar] [CrossRef] [PubMed]
- Wedeking, A.; Hagen-Euteneuer, N.; Gurgui, M.; Broere, R.; Lentzen, G.; Tolba, R.H.; Galinski, E.; van Echten-Deckert, G. A lipid anchor improves the protective effect of ectoine in inflammation. Curr. Med. Chem. 2014, 21, 2565–2572. [Google Scholar] [CrossRef] [PubMed]
- Graf, R.; Anzali, S.; Buenger, J.; Pfluecker, F.; Driller, H. The multifunctional role of ectoine as a natural cell protectant. Clin. Dermatol. 2008, 26, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Jorge, C.D.; Borges, N.; Bagyan, I.; Bilstein, A.; Santos, H. Potential applications of stress solutes from extremophiles in protein folding diseases and healthcare. Extremophiles 2016, 20, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Bownik, A.; Stepniewska, Z. Ectoine as a promising protective agent in humans and animals. Arch. Ind. Hig. Toksikol. 2016, 67, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Strong, P.J.; Kalyuzhnaya, M.; Silverman, J.; Clarke, W.P. A methanotroph-based biorefinery: Potential scenarios for generating multiple products from a single fermentation. Bioresour. Technol. 2016, 215, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Cantera, S.; Munoz, R.; Lebrero, R.; Lopez, J.C.; Rodriguez, Y.; Garcia-Encina, P.A. Technologies for the bioconversion of methane into more valuable products. Curr. Opin. Biotechnol. 2018, 50, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Vyrides, I.; Stuckey, D.C. Compatible solute addition to biological systems treating waste/wastewater to counteract osmotic and other environmental stresses: A review. Crit. Rev. Biotechnol. 2017, 37, 865–879. [Google Scholar] [CrossRef] [PubMed]
- Czech, L.; Stöveken, N.; Bremer, E. EctD-mediated biotransformation of the chemical chaperone ectoine into hydroxyectoine and its mechanosensitive channel-independent excretion. Microb. Cell Fact. 2016, 15, 126. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.; Schafer, R.; Kohlstedt, M.; Harder, B.J.; Borchert, N.S.; Stöveken, N.; Bremer, E.; Wittmann, C. Systems metabolic engineering of Corynebacterium glutamicum for production of the chemical chaperone ectoine. Microb. Cell Fact. 2013, 12, 110. [Google Scholar] [CrossRef] [PubMed]
- Ning, Y.; Wu, X.; Zhang, C.; Xu, Q.; Chen, N.; Xie, X. Pathway construction and metabolic engineering for fermentative production of ectoine in Escherichia coli. Metabol. Eng. 2016, 36, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Moya, J.; Argandona, M.; Iglesias-Guerra, F.; Nieto, J.J.; Vargas, C. Temperature- and salinity-decoupled overproduction of hydroxyectoine by Chromohalobacter salexigens. Appl. Environ. Microbiol. 2013, 79, 1018–1023. [Google Scholar] [CrossRef] [PubMed]
- Perez-Garcia, F.; Ziert, C.; Risse, J.M.; Wendisch, V.F. Improved fermentative production of the compatible solute ectoine by Corynebacterium glutamicum from glucose and alternative carbon sources. J. Biotechnol. 2017, 258, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhang, S.; Jiang, P.X.; Yao, J.; He, Y.Z.; Chen, L.C.; Gui, X.W.; Dong, Z.Y.; Tang, S.Y. Design of an ectoine-responsive arac mutant and its application in metabolic engineering of ectoine biosynthesis. Metabol. Eng. 2015, 30, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.C.; Hsu, C.C.; Lan, J.C.; Chang, Y.K.; Wang, L.F.; Wei, Y.H. Production and characterization of ectoine using a moderately halophilic strain Halomonas salina BCRC17875. J. BioSci. Bioeng. 2018. [Google Scholar] [CrossRef] [PubMed]
- Schwibbert, K.; Marin-Sanguino, A.; Bagyan, I.; Heidrich, G.; Lentzen, G.; Seitz, H.; Rampp, M.; Schuster, S.C.; Klenk, H.P.; Pfeiffer, F.; et al. A blueprint of ectoine metabolism from the genome of the industrial producer Halomonas elongata DSM 2581T. Environ. Microbiol. 2011, 13, 1973–1994. [Google Scholar] [CrossRef] [PubMed]
- Sauer, T.; Galinski, E.A. Bacterial milking: A novel bioprocess for production of compatible solutes. Biotechnol. Bioeng. 1998, 57, 306–313. [Google Scholar] [CrossRef]
- Rosa, L.T.; Bianconi, M.E.; Thomas, G.H.; Kelly, D.J. Tripartite ATP-independent periplasmic (TRAP) tansporters and tripartite tricarboxylate transporters (TTT): From uptake to pathogenicity. Front. Cell. Infect. Microbiol. 2018, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- Grammann, K.; Volke, A.; Kunte, H.J. New type of osmoregulated solute transporter identified in halophilic members of the bacteria domain: TRAP transporter TeaABC mediates uptake of ectoine and hydroxyectoine in Halomonas elongata DSM 2581(T). J. Bacteriol. 2002, 184, 3078–3085. [Google Scholar] [CrossRef] [PubMed]
- Cyplik, P.; Piotrowska-Cyplik, A.; Marecik, R.; Czarny, J.; Drozdzynska, A.; Chrzanowski, L. Biological denitrification of brine: The effect of compatible solutes on enzyme activities and fatty acid degradation. Biodegradation 2012, 23, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Peng, Y.; Wang, S.; Liu, T.; Xiao, H. Enhancement of anammox activity by addition of compatible solutes at high salinity conditions. Bioresour. Technol. 2014, 167, 560–563. [Google Scholar] [CrossRef] [PubMed]
- Peters, P.; Galinski, E.A.; Trüper, H.G. The biosyntheis of ectoine. FEMS Microbiol. Lett. 1990, 71, 157–162. [Google Scholar] [CrossRef]
- Ono, H.; Sawada, K.; Khunajakr, N.; Tao, T.; Yamamoto, M.; Hiramoto, M.; Shinmyo, A.; Takano, M.; Murooka, Y. Characterization of biosynthetic enzymes for ectoine as a compatible solute in a moderately halophilic eubacterium, Halomonas elongata. J. Bacteriol. 1999, 181, 91–99. [Google Scholar] [PubMed]
- Reshetnikov, A.S.; Khmelenina, V.N.; Mustakhimov, I.I.; Trotsenko, Y.A. Genes and enzymes of ectoine biosynthesis in halotolerant methanotrophs. Methods Enzymol. 2011, 495, 15–30. [Google Scholar] [PubMed]
- Lo, C.C.; Bonner, C.A.; Xie, G.; D’Souza, M.; Jensen, R.A. Cohesion group approach for evolutionary analysis of aspartokinase, an enzyme that feeds a branched network of many biochemical pathways. Microbiol. Mol. Biol. Rev. 2009, 73, 594–651. [Google Scholar] [CrossRef] [PubMed]
- Reshetnikov, A.S.; Khmelenina, V.N.; Trotsenko, Y.A. Characterization of the ectoine biosynthesis genes of haloalkalotolerant obligate methanotroph “Methylomicrobium alcaliphilum 20z”. Arch. Microbiol. 2006, 184, 286–297. [Google Scholar] [CrossRef] [PubMed]
- Nyyssölä, A.; Kerovuo, J.; Kaukinen, P.; von Weymarn, N.; Reinikainen, T. Extreme halophiles synthesize betaine from glycine by methylation. J. Biol. Chem. 2000, 275, 22196–22201. [Google Scholar] [CrossRef] [PubMed]
- Boch, J.; Kempf, B.; Schmid, R.; Bremer, E. Synthesis of the osmoprotectant glycine betaine in Bacillus subtilis: Characterization of the gbsAB genes. J. Bacteriol. 1996, 178, 5121–5129. [Google Scholar] [CrossRef] [PubMed]
- Lamark, T.; Kaasen, I.; Eshoo, M.W.; Falkenberg, P.; McDougall, J.; Strom, A.R. DNA sequence and analysis of the bet genes encoding the osmoregulatory choline-glycine betaine pathway of Escherichia coli. Mol. Microbiol. 1991, 5, 1049–1064. [Google Scholar] [CrossRef] [PubMed]
- Salvi, F.; Wang, Y.F.; Weber, I.T.; Gadda, G. Structure of choline oxidase in complex with the reaction product glycine betaine. Acta Crystallogr. D Biol. Crystallogr. 2014, 70, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, E.F.; Cerqueira, N.M.; Fernandes, P.A.; Ramos, M.J. Mechanism of formation of the internal aldimine in pyridoxal 5′-phosphate-dependent enzymes. J. Am. Chem. Soc. 2011, 133, 15496–15505. [Google Scholar] [CrossRef] [PubMed]
- Vetting, M.W.; de Carvalho, L.P.S.; Yu, M.; Hegde, S.S.; Magnet, S.; Roderick, S.L.; Blanchard, J.S. Structure and functions of the GNAT superfamily of acetyltransferases. Arch. Biochem. Biophys. 2005, 433, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Mustakhimov, I.I.; Rozova, O.N.; Reshetnikov, A.S.; Khmelenina, V.N.; Murrell, J.C.; Trotsenko, Y.A. Characterization of the recombinant diaminobutyric acid acetyltransferase from Methylophaga thalassica and Methylophaga alcalica. FEMS Microbiol. Lett. 2008, 283, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Widderich, N.; Kobus, S.; Höppner, A.; Ricela, R.; Seubert, A.; Dickschat, J.S.; Heider, J.; Smits, S.H.J.; Bremer, E. Biochemistry and crystal structure of the ectoine synthase: A metal-containing member of the cupin superfamily. PLoS ONE 2016, 11, e0151285. [Google Scholar] [CrossRef] [PubMed]
- Höppner, A.; Widderich, N.; Lenders, M.; Bremer, E.; Smits, S.H.J. Crystal structure of the ectoine hydroxylase, a snapshot of the active site. J. Biol. Chem. 2014, 289, 29570–29583. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Estepa, R.; Canovas, D.; Iglesias-Guerra, F.; Ventosa, A.; Csonka, L.N.; Nieto, J.J.; Vargas, C. Osmoprotection of Salmonella enterica serovar Typhimurium by N-γ-acetyldiaminobutyrate, the precursor of the compatible solute ectoine. Syst. Appl. Microbiol. 2006, 29, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Canovas, D.; Borges, N.; Vargas, C.; Ventosa, A.; Nieto, J.J.; Santos, H. Role of N-γ-acetyldiaminobutyrate as an enzyme stabilizer and an intermediate in the biosynthesis of hydroxyectoine. Appl. Environ. Microbiol. 1999, 65, 3774–3779. [Google Scholar] [PubMed]
- Moritz, K.D.; Amendt, B.; Witt, E.M.H.J.; Galinski, E.A. The hydroxyectoine gene cluster of the non-halophilic acidophile Acidiphilium cryptum. Extremophiles 2015, 19, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Witt, E.M.; Davies, N.W.; Galinski, E.A. Unexpected property of ectoine synthase and its application for synthesis of the engineered compatible solute ADPC. Appl. Microbiol. Biotechnol. 2011, 91, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Dunwell, J.M.; Purvis, A.; Khuri, S. Cupins: The most functionally diverse protein superfamily? Phytochemistry 2004, 65, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Dunwell, J.M.; Culham, A.; Carter, C.E.; Sosa-Aguirre, C.R.; Goodenough, P.W. Evolution of functional diversity in the cupin superfamily. Trends Biochem. Sci. 2001, 26, 740–746. [Google Scholar] [CrossRef]
- Reuter, K.; Pittelkow, M.; Bursy, J.; Heine, A.; Craan, T.; Bremer, E. Synthesis of 5-hydroxyectoine from ectoine: Crystal structure of the non-heme iron(II) and 2-oxoglutarate-dependent dioxygenase EctD. PLoS ONE 2010, 5, e10647. [Google Scholar] [CrossRef] [PubMed]
- Clifton, I.J.; McDonough, M.A.; Ehrismann, D.; Kershaw, N.J.; Granatino, N.; Schofield, C.J. Structural studies on 2-oxoglutarate oxygenases and related double-stranded beta-helix fold proteins. J. Inorg. Biochem. 2006, 100, 644–669. [Google Scholar] [CrossRef] [PubMed]
- Widderich, N.; Pittelkow, M.; Hoppner, A.; Mulnaes, D.; Buckel, W.; Gohlke, H.; Smits, S.H.; Bremer, E. Molecular dynamics simulations and structure-guided mutagenesis provide insight into the architecture of the catalytic core of the ectoine hydroxylase. J. Mol. Biol. 2014, 426, 586–600. [Google Scholar] [CrossRef] [PubMed]
- Jebbar, M.; Talibart, R.; Gloux, K.; Bernard, T.; Blanco, C. Osmoprotection of Escherichia coli by ectoine: Uptake and accumulation characteristics. J. Bacteriol. 1992, 174, 5027–5035. [Google Scholar] [CrossRef] [PubMed]
- Lucht, J.M.; Bremer, E. Adaptation of Escherichia coli to high osmolarity environments: Osmoregulation of the high-affinity glycine betaine transport system ProU. FEMS Microbiol. Rev. 1994, 14, 3–20. [Google Scholar] [CrossRef] [PubMed]
- MacMillan, S.V.; Alexander, D.A.; Culham, D.E.; Kunte, H.J.; Marshall, E.V.; Rochon, D.; Wood, J.M. The ion coupling and organic substrate specificities of osmoregulatory transporter ProP in Escherichia coli. Biochim. Biophys. Acta 1999, 1420, 30–44. [Google Scholar] [CrossRef]
- Rodriguez-Moya, J.; Argandona, M.; Reina-Bueno, M.; Nieto, J.J.; Iglesias-Guerra, F.; Jebbar, M.; Vargas, C. Involvement of EupR, a response regulator of the NarL/FixJ family, in the control of the uptake of the compatible solutes ectoines by the halophilic bacterium Chromohalobacter salexigens. BMC Microbiol. 2010, 10, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bestvater, T.; Louis, P.; Galinski, E.A. Heterologous ectoine production in Escherichia coli: By-passing the metabolic bottle-neck. Saline Syst. 2008, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuraku, S.; Zmasek, C.M.; Nishimura, O.; Katoh, K. aLeaves facilitates on-demand exploration of metazoan gene family trees on MAFFT sequence alignment server with enhanced interactivity. Nucleic Acids Res. 2013, 41, W22–W28. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef] [PubMed]
- Salar-Garcia, M.J.; Bernal, V.; Pastor, J.M.; Salvador, M.; Argandona, M.; Nieto, J.J.; Vargas, C.; Canovas, M. Understanding the interplay of carbon and nitrogen supply for ectoines production and metabolic overflow in high density cultures of Chromohalobacter salexigens. Microb. Cell Fact. 2017, 16, 23. [Google Scholar] [CrossRef] [PubMed]
- Pastor, J.M.; Bernal, V.; Salvador, M.; Argandona, M.; Vargas, C.; Csonka, L.; Sevilla, A.; Iborra, J.L.; Nieto, J.J.; Canovas, M. Role of central metabolism in the osmoadaptation of the halophilic bacterium Chromohalobacter salexigens. J. Biol. Chem. 2013, 288, 17769–17781. [Google Scholar] [CrossRef] [PubMed]
- Piubeli, F.; Salvador, M.; Argandona, M.; Nieto, J.J.; Bernal, V.; Pastor, J.M.; Canovas, M.; Vargas, C. Insights into metabolic osmoadaptation of the ectoines-producer bacterium Chromohalobacter salexigens through a high-quality genome scale metabolic model. Microb. Cell Fact. 2018, 17, 2. [Google Scholar] [CrossRef] [PubMed]
- Kindzierski, V.; Raschke, S.; Knabe, N.; Siedler, F.; Scheffer, B.; Pfluger-Grau, K.; Pfeiffer, F.; Oesterhelt, D.; Marin-Sanguino, A.; Kunte, H.J. Osmoregulation in the halophilic bacterium Halomonas elongata: A case study for integrative systems biology. PLoS ONE 2017, 12, e0168818. [Google Scholar] [CrossRef] [PubMed]
- Mustakhimov, I.I.; Reshetnikov, A.S.; Glukhov, A.S.; Khmelenina, V.N.; Kalyuzhnaya, M.G.; Trotsenko, Y.A. Identification and characterization of EctR1, a new transcriptional regulator of the ectoine biosynthesis genes in the halotolerant methanotroph Methylomicrobium alcaliphilum 20Z. J. Bacteriol. 2010, 192, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Leon, M.J.; Hoffmann, T.; Sanchez-Porro, C.; Heider, J.; Ventosa, A.; Bremer, E. Compatible solute synthesis and import by the moderate halophile Spiribacter salinus: Physiology and genomics. Front. Microbiol. 2018, 9, 108. [Google Scholar] [CrossRef] [PubMed]
- León, M.J.; Fernandez, A.B.; Ghai, R.; Sanchez-Porro, C.; Rodriguez-Valera, F.; Ventosa, A. From metagenomics to pure culture: Isolation and characterization of the moderately halophilic bacterium Spiribacter salinus gen. nov., sp. nov. Appl. Environ. Microbiol. 2014, 80, 3850–3857. [Google Scholar]
- Haldenwang, W.G. The sigma factors of Bacillus subtilis. Microbiol. Rev. 1995, 59, 1–30. [Google Scholar] [PubMed]
- Hecker, M.; Pane-Farre, J.; Völker, U. SigB-dependent general stress response in Bacillus subtilis and related gram-positive bacteria. Annu. Rev. Microbiol. 2007, 61, 215–236. [Google Scholar] [CrossRef] [PubMed]
- Nannapaneni, P.; Hertwig, F.; Depke, M.; Hecker, M.; Mäder, U.; Völker, U.; Steil, L.; van Hijum, S.A. Defining the structure of the general stress regulon of Bacillus subtilis using targeted microarray analysis and random forest classification. Microbiology 2012, 158, 696–707. [Google Scholar] [CrossRef] [PubMed]
- Feklistov, A.; Sharon, B.D.; Darst, S.A.; Gross, C.A. Bacterial sigma factors: A historical, structural, and genomic perspective. Annu. Rev. Microbiol. 2014, 68, 357–376. [Google Scholar] [CrossRef] [PubMed]
- Typas, A.; Becker, G.; Hengge, R. The molecular basis of selective promoter activation by the sigmaS subunit of RNA polymerase. Mol. Microbiol. 2007, 63, 1296–1306. [Google Scholar] [CrossRef] [PubMed]
- Bestvater, T.; Galinski, E.A. Investigation into a stress-inducible promoter region from Marinococcus halophilus using green fluorescent protein. Extremophiles 2002, 6, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Gralla, J.D.; Huo, Y.X. Remodeling and activation of Escherichia coli RNA polymerase by osmolytes. Biochemistry 2008, 47, 13189–13196. [Google Scholar] [CrossRef] [PubMed]
- Gralla, J.D.; Vargas, D.R. Potassium glutamate as a transcriptional inhibitor during bacterial osmoregulation. EMBO J. 2006, 25, 1515–1521. [Google Scholar] [CrossRef] [PubMed]
- Salvador, M.; Argandona, M.; Pastor, J.M.; Bernal, V.; Canovas, M.; Csonka, L.N.; Nieto, J.J.; Vargas, C. Contribution of RpoS to metabolic efficiency and ectoines synthesis during the osmo- and heat-stress response in the halophilic bacterium Chromohalobacter salexigens. Environ. Microbiol. Rep. 2015, 7, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Higgins, C.F.; Dorman, C.J.; Stirling, D.A.; Waddell, L.; Booth, I.R.; May, G.; Bremer, E. A physiological role for DNA supercoiling in the osmotic regulation of gene expression in S. typhimurium and E. coli. Cell 1988, 52, 569–584. [Google Scholar] [CrossRef]
- Booth, I.R.; Higgins, C.F. Enteric bacteria and osmotic stress: Intracellular potassium glutamate as a secondary signal of osmotic stress? FEMS Microbiol. Rev. 1990, 6, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.; Wensing, A.; Brosius, M.; Steil, L.; Völker, U.; Bremer, E. Osmotic control of opuA expression in Bacillus subtilis and its modulation in response to intracellular glycine betaine and proline pools. J. Bacteriol. 2013, 195, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.; Bleisteiner, M.; Sappa, P.K.; Steil, L.; Mader, U.; Volker, U.; Bremer, E. Synthesis of the compatible solute proline by Bacillus subtilis: Point mutations rendering the osmotically controlled proHJ promoter hyperactive. Environ. Microbiol. 2017, 19, 3700–3720. [Google Scholar] [CrossRef] [PubMed]
- Deochand, D.K.; Grove, A. MarR family transcription factors: Dynamic variations on a common scaffold. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 595–613. [Google Scholar] [CrossRef] [PubMed]
- Mustakhimov, I.I.; Reshetnikov, A.S.; Fedorov, D.N.; Khmelenina, V.N.; Trotsenko, Y.A. Role of RctR as transcriptional regulator of ectoine biosynthesis genes in Methylophaga thalassica. Biochem. Biokhimiia 2012, 77, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Romeo, Y.; Bouvier, J.; Gutierrez, C. Osmotic regulation of transcription in Lactococcus lactis: Ionic strength-dependent binding of the BusR repressor to the busA promoter. FEBS Lett. 2007, 581, 3387–3390. [Google Scholar] [CrossRef] [PubMed]
- Romeo, Y.; Obis, D.; Bouvier, J.; Guillot, A.; Fourcans, A.; Bouvier, I.; Gutierrez, C.; Mistou, M.Y. Osmoregulation in Lactococcus lactis: BusR, a transcriptional repressor of the glycine betaine uptake system BusA. Mol. Microbiol. 2003, 47, 1135–1147. [Google Scholar] [CrossRef] [PubMed]
- Shikuma, N.J.; Davis, K.R.; Fong, J.N.C.; Yildiz, F.H. The transcriptional regulator, CosR, controls compatible solute biosynthesis and transport, motility and biofilm formation in Vibrio cholerae. Environ. Microbiol. 2013, 15, 1387–1399. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.; Deng, W.; Li, S.; He, J.; Ren, S.; Huang, W.; Lu, Y.; Zhao, G.; Cai, Z.; Wang, J. GlnR-mediated regulation of ectABCD transcription expands the role of the GlnR regulon to osmotic stress management. J. Bacteriol. 2015, 197, 3041–3307. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, A.; Soltani, B.M.; Nekouei, M.K.; Jouzani, G.S.; Mirzaei, H.H.; Sadeghizadeh, M. Diversity of the ectoines biosynthesis genes in the salt tolerant Streptomyces and evidence for inductive effect of ectoines on their accumulation. Microbiol. Res. 2014, 169, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Kurz, M.; Burch, A.Y.; Seip, B.; Lindow, S.E.; Gross, H. Genome-driven investigation of compatible solute biosynthesis pathways of Pseudomonas syringae pv. syringae and their contribution to water stress tolerance. Appl. Environ. Microbiol. 2010, 76, 5452–5462. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.A.; Markowitz, V.M.; Chu, K.; Palaniappan, K.; Szeto, E.; Pillay, M.; Ratner, A.; Huang, J.; Andersen, E.; Huntemann, M.; et al. IMG/M: Integrated genome and metagenome comparative data analysis system. Nucleic Acids Res. 2017, 45, D507–D516. [Google Scholar] [CrossRef] [PubMed]
- Soucy, S.M.; Huang, J.; Gogarten, J.P. Horizontal gene transfer: Building the web of life. Nat. Rev. Genet. 2015, 16, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Treangen, T.J.; Rocha, E.P.C. Horizontal transfer, not duplication, drives the expansion of protein families in prokaryotes. PLoS Genet. 2011, 7, e1001284. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.; Whitaker, R.J.; Krause, D.J.; Heilers, J.H.; van Wolferen, M.; van der Does, C.; Albers, S.V. Mechanisms of gene flow in archaea. Nat. Rev. Microbiol. 2017, 15, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.C.; Madupu, R.; Durkin, A.S.; Ekborg, N.A.; Pedamallu, C.S.; Hostetler, J.B.; Radune, D.; Toms, B.S.; Henrissat, B.; Coutinho, P.M.; et al. The complete genome of Teredinibacter turnerae T7901: An intracellular endosymbiont of marine wood-boring bivalves (shipworms). PLoS ONE 2009, 4, e6085. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.; Soares, A.R.; Lamosa, P.; Santos, M.A.; da Costa, M.S. Comparison of the compatible solute pool of two slightly halophilic planctomycetes species, Gimesia maris and Rubinisphaera brasiliensis. Extremophiles 2016, 20, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Amiri Moghaddam, J.; Boehringer, N.; Burdziak, A.; Kunte, H.J.; Galinski, E.A.; Schäberle, T.F. Different strategies of osmoadaptation in the closely related marine myxobacteria Enhygromyxa salina SWB007 and Plesiocystis pacifica SIR-1. Microbiology 2016, 162, 641–661. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Heal, K.R.; Ramdasi, R.; Kobelt, J.N.; Martens-Habbena, W.; Bertagnolli, A.D.; Amin, S.A.; Walker, C.B.; Urakawa, H.; Konneke, M.; et al. Nitrosopumilus maritimus gen. nov., sp. nov., Nitrosopumilus cobalaminigenes sp. nov., Nitrosopumilus oxyclinae sp. nov., and Nitrosopumilus ureiphilus sp. Nov., four marine ammonia-oxidizing archaea of the phylum thaumarchaeota. Int. J. Syst. Evol. Microbiol. 2017, 67, 5067–5079. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.; Scheuner, C.; Meier-Kolthoff, J.P.; Brinkhoff, T.; Wagner-Döbler, I.; Ulbrich, M.; Klenk, H.P.; Schomburg, D.; Petersen, J.; Göker, M. Phylogenomics of Rhodobacteraceae reveals evolutionary adaptation to marine and non-marine habitats. ISME J. 2017, 11, 1483–1499. [Google Scholar] [CrossRef] [PubMed]
- Wagner-Döbler, I.; Biebl, H. Environmental biology of the marine Roseobacter lineage. Annu. Rev. Microbiol. 2006, 60, 255–280. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Moran, M.A. Evolutionary ecology of the marine Roseobacter clade. Microbiol. Mol. Biol. Rev. 2014, 78, 573–587. [Google Scholar] [CrossRef] [PubMed]
- Welsh, D.T. Ecological significance of compatible solute accumulation by micro-organisms: From single cells to global climate. FEMS Microbiol. Rev. 2000, 24, 263–290. [Google Scholar] [CrossRef] [PubMed]
- Warren, C. Do microbial osmolytes or extracellular depolymerization products accumulate as soil dries? Soil Biol. Biochem. 2016, 98, 54–63. [Google Scholar] [CrossRef]
- Warren, C.R. Quaternary ammonium compounds can be abundant in some soils and are taken up as intact molecules by plants. New Phytol. 2013, 198, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Warren, C.R. Response of osmolytes in soil to drying and rewetting. Soil Biol. Biochem. 2014, 70, 22–32. [Google Scholar] [CrossRef]
- Bouskill, N.J.; Wood, T.E.; Baran, R.; Hao, Z.; Ye, Z.; Bowen, B.P.; Lim, H.C.; Nico, P.S.; Holman, H.Y.; Gilbert, B.; et al. Belowground response to drought in a tropical forest soil. II. Change in microbial function impacts carbon composition. Front. Microbiol. 2016, 7, 323. [Google Scholar] [CrossRef] [PubMed]
- Bouskill, N.J.; Wood, T.E.; Baran, R.; Ye, Z.; Bowen, B.P.; Lim, H.; Zhou, J.; Nostrand, J.D.; Nico, P.; Northen, T.R.; et al. Belowground response to drought in a tropical forest soil. I. Changes in microbial functional potential and metabolism. Front. Microbiol. 2016, 7, 525. [Google Scholar] [CrossRef] [PubMed]
- Mosier, A.C.; Justice, N.B.; Bowen, B.P.; Baran, R.; Thomas, B.C.; Northen, T.R.; Banfield, J.F. Metabolites associated with adaptation of microorganisms to an acidophilic, metal-rich environment identified by stable-isotope-enabled metabolomics. mBio 2013, 4, e00484-12. [Google Scholar] [CrossRef] [PubMed]
- Poolman, B.; Spitzer, J.J.; Wood, J.M. Bacterial osmosensing: Roles of membrane structure and electrostatics in lipid-protein and protein-protein interactions. Biochim. Biophys. Acta 2004, 1666, 88–104. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, C.; Bremer, E.; Krämer, R. The BCCT family of carriers: From physiology to crystal structure. Mol. Microbiol. 2010, 78, 13–34. [Google Scholar] [CrossRef] [PubMed]
- Krämer, R. Bacterial stimulus perception and signal transduction: Response to osmotic stress. Chem. Rec. 2010, 10, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Culham, D.E.; Shkel, I.A.; Record, M.T., Jr.; Wood, J.M. Contributions of coulombic and Hofmeister effects to the osmotic activation of Escherichia coli transporter ProP. Biochemistry 2016, 55, 1301–1313. [Google Scholar] [CrossRef] [PubMed]
- Gul, N.; Schuurman-Wolters, G.; Karasawa, A.; Poolman, B. Functional characterization of amphipathic alpha-helix in the osmoregulatory ABC transporter OpuA. Biochemistry 2012, 51, 5142–5152. [Google Scholar] [CrossRef] [PubMed]
- Perez, C.; Faust, B.; Mehdipour, A.R.; Francesconi, K.A.; Forrest, L.R.; Ziegler, C. Substrate-bound outward-open state of the betaine transporter BetP provides insights into Na+ coupling. Nat. Commun. 2014, 5, 4231. [Google Scholar] [CrossRef] [PubMed]
- Perez, C.; Koshy, C.; Yildiz, O.; Ziegler, C. Alternating-access mechanism in conformationally asymmetric trimers of the betaine transporter BetP. Nature 2012, 490, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Hahne, H.; Mäder, U.; Otto, A.; Bonn, F.; Steil, L.; Bremer, E.; Hecker, M.; Becher, D. A comprehensive proteomics and transcriptomics analysis of Bacillus subtilis salt stress adaptation. J. Bacteriol. 2010, 192, 870–882. [Google Scholar] [CrossRef] [PubMed]
- Kappes, R.M.; Kempf, B.; Kneip, S.; Boch, J.; Gade, J.; Meier-Wagner, J.; Bremer, E. Two evolutionarily closely related ABC transporters mediate the uptake of choline for synthesis of the osmoprotectant glycine betaine in Bacillus subtilis. Mol. Microbiol. 1999, 32, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Morbach, S.; Krämer, R. Body shaping under water stress: Osmosensing and osmoregulation of solute transport in bacteria. ChemBioChem 2002, 3, 384–397. [Google Scholar] [CrossRef]
- Rice, A.J.; Park, A.; Pinkett, H.W. Diversity in ABC transporters: Type I, II and III importers. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Ter Beek, J.; Guskov, A.; Slotboom, D.J. Structural diversity of ABC transporters. J. Gen. Physiol. 2014, 143, 419–435. [Google Scholar] [CrossRef] [PubMed]
- Jebbar, M.; von Blohn, C.; Bremer, E. Ectoine functions as an osmoprotectant in Bacillus subtilis and is accumulated via the ABC-transport system OpuC. FEMS Microbiol. Lett. 1997, 154, 325–330. [Google Scholar] [CrossRef]
- Choquet, G.; Jehan, N.; Pissavin, C.; Blanco, C.; Jebbar, M. OusB, a broad-specificity ABC-type transporter from Erwinia chrysanthemi, mediates uptake of glycine betaine and choline with a high affinity. Appl. Environ. Microbiol. 2005, 71, 3389–3398. [Google Scholar] [CrossRef] [PubMed]
- Yan, N. Structural biology of the major facilitator superfamily transporters. Annu. Rev. Biophys. 2015, 44, 257–283. [Google Scholar] [CrossRef] [PubMed]
- Gloux, K.; Touze, T.; Pagot, Y.; Jouan, B.; Blanco, C. Mutations of ousA alter the virulence of Erwinia chrysanthemi. Mol. Plant. Microbe Interact. 2005, 18, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Kappes, R.M.; Kempf, B.; Bremer, E. Three transport systems for the osmoprotectant glycine betaine operate in Bacillus subtilis: Characterization of OpuD. J. Bacteriol. 1996, 178, 5071–5079. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, V.; Kunte, H.J. Marinococcus halophilus DSM 20408T encodes two transporters for compatible solutes belonging to the betaine-carnitine-choline transporter family: Identification and characterization of ectoine transporter EctM and glycine betaine transporter BetM. Extremophiles 2004, 8, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Steger, R.; Weinand, M.; Krämer, R.; Morbach, S. LcoP, an osmoregulated betaine/ectoine uptake system from Corynebacterium glutamicum. FEBS Lett. 2004, 573, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Peter, H.; Weil, B.; Burkovski, A.; Kramer, R.; Morbach, S. Corynebacterium glutamicum is equipped with four secondary carriers for compatible solutes: Identification, sequencing, and characterization of the proline/ectoine uptake system, ProP, and the ectoine/proline/glycine betaine carrier, EctP. J. Bacteriol. 1998, 180, 6005–6012. [Google Scholar] [PubMed]
- Perez, C.; Koshy, C.; Ressl, S.; Nicklisch, S.; Krämer, R.; Ziegler, C. Substrate specificity and ion coupling in the Na+/betaine symporter BetP. EMBO J. 2011, 30, 1221–1229. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmann, S.I.; Terwisscha van Scheltinga, A.C.; Bienert, R.; Kunte, H.J.; Ziegler, C. 1.55 a structure of the ectoine binding protein TeaA of the osmoregulated TRAP-transporter TeaABC from Halomonas elongata. Biochemistry 2008, 47, 9475–9485. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Shi, W.W.; He, Y.X.; Yang, Y.H.; Zhou, C.Z.; Chen, Y. Structures of the substrate-binding protein provide insights into the multiple compatible solute binding specificities of the Bacillus subtilis ABC transporter OpuC. Biochem. J. 2011, 436, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Schulz, A.; Stöveken, N.; Binzen, I.M.; Hoffmann, T.; Heider, J.; Bremer, E. Feeding on compatible solutes: A substrate-induced pathway for uptake and catabolism of ectoines and its genetic control by EnuR. Environ. Microbiol. 2017, 19, 926–946. [Google Scholar] [CrossRef] [PubMed]
- Galinski, E.A.; Herzog, R.M. The role of trehalose as a substitute for nitrogen-containing compatible solutes (Ectothiorhodospira halochloris). Arch. Microbiol. 1990, 153, 607–613. [Google Scholar] [CrossRef]
- Jebbar, M.; Sohn-Bosser, L.; Bremer, E.; Bernard, T.; Blanco, C. Ectoine-induced proteins in Sinorhizobium meliloti include an ectoine ABC-type transporter involved in osmoprotection and ectoine catabolism. J. Bacteriol. 2005, 187, 1293–1304. [Google Scholar] [CrossRef] [PubMed]
- Vargas, C.; Jebbar, M.; Carrasco, R.; Blanco, C.; Calderon, M.I.; Iglesias-Guerra, F.; Nieto, J.J. Ectoines as compatible solutes and carbon and energy sources for the halophilic bacterium Chromohalobacter salexigens. J. Appl. Microbiol. 2006, 100, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Hanekop, N.; Höing, M.; Sohn-Bösser, L.; Jebbar, M.; Schmitt, L.; Bremer, E. Crystal structure of the ligand-binding protein EhuB from Sinorhizobium meliloti reveals substrate recognition of the compatible solutes ectoine and hydroxyectoine. J. Mol. Biol. 2007, 374, 1237–1250. [Google Scholar] [CrossRef] [PubMed]
- Lecher, J.; Pittelkow, M.; Zobel, S.; Bursy, J.; Bonig, T.; Smits, S.H.; Schmitt, L.; Bremer, E. The crystal structure of UehA in complex with ectoine—A comparison with other TRAP-T binding proteins. J. Mol. Biol. 2009, 389, 58–73. [Google Scholar] [CrossRef] [PubMed]
- Rigali, S.; Derouaux, A.; Giannotta, F.; Dusart, J. Subdivision of the helix-turn-helix GntR family of bacterial regulators in the FadR, HutC, MocR, and YtrA subfamilies. J. Biol. Chem. 2002, 277, 12507–12515. [Google Scholar] [CrossRef] [PubMed]
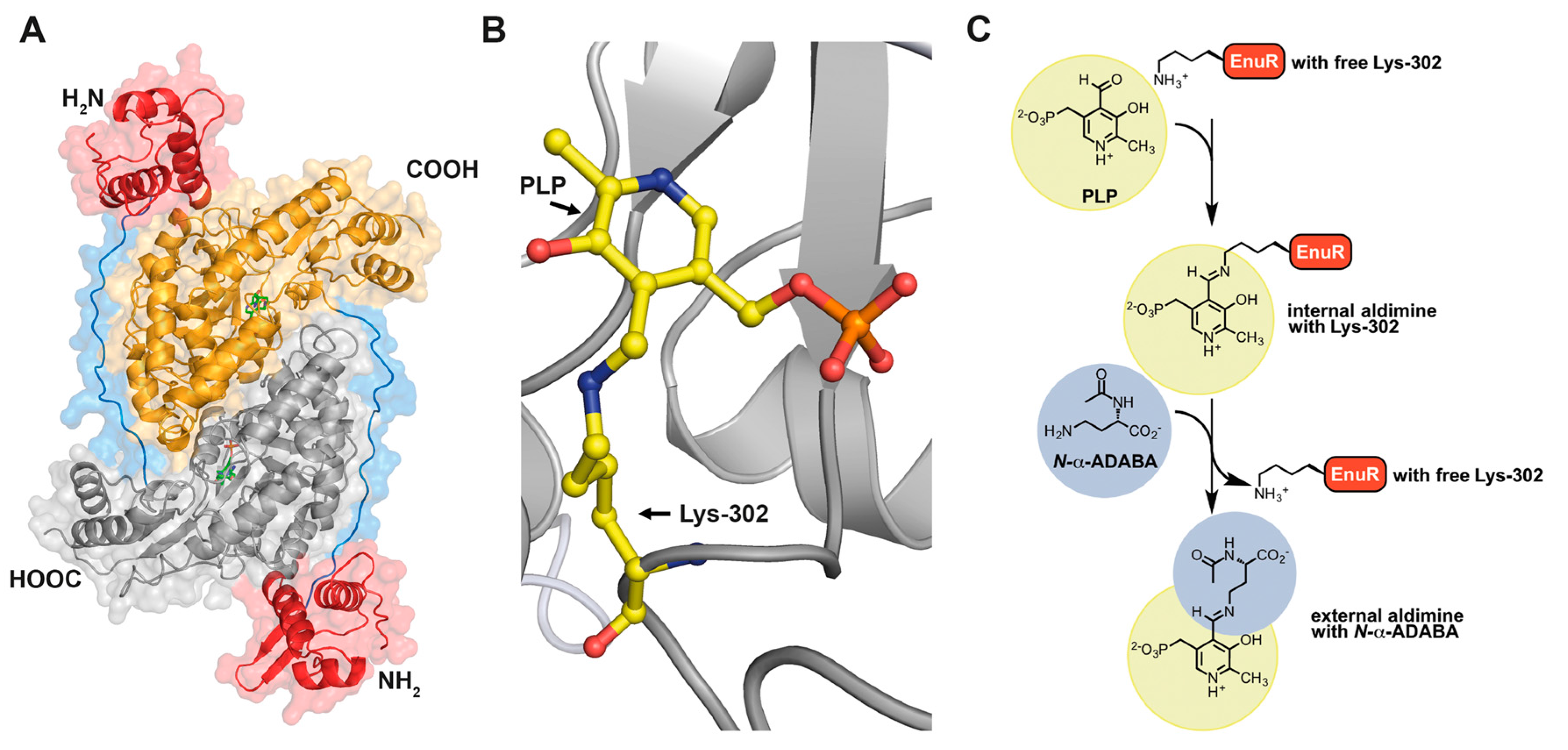
- Schulz, A.; Hermann, L.; Freibert, S.-A.; Bönig, T.; Hoffmann, T.; Riclea, R.; Dickschat, J.S.; Heider, J.; Bremer, E. Transcriptional regulation of ectoine catabolism in response to multiple metabolic and environmnetal cues. Environ. Microbiol. 2017, 19, 4599–4619. [Google Scholar] [CrossRef] [PubMed]
- Kumarevel, T.; Nakano, N.; Ponnuraj, K.; Gopinath, S.C.; Sakamoto, K.; Shinkai, A.; Kumar, P.K.; Yokoyama, S. Crystal structure of glutamine receptor protein from Sulfolobus tokodaii strain 7 in complex with its effector l-glutamine: Implications of effector binding in molecular association and DNA binding. Nucleic Acids Res. 2008, 36, 4808–4820. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, T.; Ramachandran, R. Mechanistic insights from the crystal structures of a feast/famine regulatory protein from Mycobacterium tuberculosis h37rv. Nucleic Acids Res. 2007, 35, 7324–7335. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, K.; Ishijima, S.A.; Clowney, L.; Koike, H.; Aramaki, H.; Tanaka, C.; Makino, K.; Suzuki, M. Feast/famine regulatory proteins (FFRPs): Escherichia coli LRP, AsnC and related archaeal transcription factors. FEMS Microbiol. Rev. 2006, 30, 89–108. [Google Scholar] [CrossRef] [PubMed]
- Copeland, A.; O’Connor, K.; Lucas, S.; Lapidus, A.; Berry, K.W.; Detter, J.C.; Del Rio, T.G.; Hammon, N.; Dalin, E.; Tice, H.; et al. Complete genome sequence of the halophilic and highly halotolerant Chromohalobacter salexigens type strain (1H11T). Stand. Genom. Sci. 2011, 5, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Bramucci, E.; Milano, T.; Pascarella, S. Genomic distribution and heterogeneity of MocR-like transcriptional factors containing a domain belonging to the superfamily of the pyridoxal-5′-phosphate dependent enzymes of fold type I. Biochem. Biophys. Res. Commun. 2011, 415, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Suvorova, I.; Rodionov, D. Comparitive genomics of pyridoxal 5′-phosphate-dependent transcription factor regulons in Bacteria. Microb. Genom. 2016, 2, e000047. [Google Scholar] [PubMed]
- Yu, Q.; Cai, H.; Zhang, Y.; He, Y.; Chen, L.; Merritt, J.; Zhang, S.; Dong, Z. Negative regulation of ectoine uptake and catabolism in Sinorhizobium meliloti: Characterization of the EhuR gene. J. Bacteriol. 2017, 199, e00119-16. [Google Scholar] [CrossRef] [PubMed]
- Belitsky, B.R. Bacillus subtilis GabR, a protein with DNA-binding and aminotransferase domains, is a PLP-dependent transcriptional regulator. J. Mol. Biol. 2004, 340, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Edayathumangalam, R.; Wu, R.; Garcia, R.; Wang, Y.; Wang, W.; Kreinbring, C.A.; Bach, A.; Liao, J.; Stone, T.A.; Terwilliger, T.C.; et al. Crystal structure of Bacillus subtilis GabR, an autorepressor and transcriptional activator of gabT. Proc. Natl. Acad. Sci. USA 2013, 110, 17820–17825. [Google Scholar] [CrossRef] [PubMed]
- Okuda, K.; Ito, T.; Goto, M.; Takenaka, T.; Hemmi, H.; Yoshimura, T. Domain characterization of Bacillus subtilis GabR, a pyridoxal 5′-phosphate-dependent transcriptional regulator. J. Biochem. 2015, 158, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Okuda, K.; Kato, S.; Ito, T.; Shiraki, S.; Kawase, Y.; Goto, M.; Kawashima, S.; Hemmi, H.; Fukada, H.; Yoshimura, T. Role of the aminotransferase domain in Bacillus subtilis GabR, a pyridoxal 5′-phosphate-dependent transcriptional regulator. Mol. Microbiol. 2015, 95, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Park, S.A.; Park, Y.S.; Lee, K.S. Crystal structure of the C-terminal domain of Bacillus subtilis GabR reveals a closed conformation by gamma-aminobutyric acid binding, inducing transcriptional activation. Biochem. Biophys. Res. Commun. 2017, 487, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Sanishvili, R.; Belitsky, B.R.; Juncosa, J.I.; Le, H.V.; Lehrer, H.J.; Farley, M.; Silverman, R.B.; Petsko, G.A.; Ringe, D.; et al. Plp and GABA trigger GabR-mediated transcription regulation in Bacillus subtilis via external aldimine formation. Proc. Natl. Acad. Sci. USA 2017, 114, 3891–3896. [Google Scholar] [CrossRef] [PubMed]
- Steffen-Munsberg, F.; Vickers, C.; Kohls, H.; Land, H.; Mallin, H.; Nobili, A.; Skalden, L.; van den Bergh, T.; Joosten, H.J.; Berglund, P.; et al. Bioinformatic analysis of a PLP-dependent enzyme superfamily suitable for biocatalytic applications. Biotechnol. Adv. 2015, 33, 566–604. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.S. Chemistry and diversity of pyridoxal-5′-phosphate dependent enzymes. Biochim. Biophys. Acta 2015, 1854, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Cassarino, T.G.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef] [PubMed]
- Delano, W.L. The PyMol Molecular Graphics System; Delano Scientific: San Carlos, CA, USA, 2002. [Google Scholar]
- Dey, A.; Shree, S.; Pandey, S.K.; Tripathi, R.P.; Ramachandran, R. Crystal structure of Mycobacterium tuberculosis H37Rv AldR (Rv2779c), a regulator of the ald gene: DNA-binding, and identification of small-molecule inhibitors. J. Biol. Chem. 2016, 291, 11967–11980. [Google Scholar] [CrossRef] [PubMed]
- Kamensek, S.; Browning, D.F.; Podlesek, Z.; Busby, S.J.; Zgur-Bertok, D.; Butala, M. Silencing of DNase colicin e8 gene expression by a complex nucleoprotein assembly ensures timely colicin induction. PLoS Genet. 2015, 11, e1005354. [Google Scholar] [CrossRef] [PubMed]
- Zschiedrich, C.P.; Keidel, V.; Szurmant, H. Molecular mechanisms of two-component signal transduction. J. Mol. Biol. 2016, 428, 3752–3775. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, I.; Cornaciu, I.; Carrica, M.D.; Uchikawa, E.; Hoffmann, G.; Sieira, R.; Marquez, J.A.; Goldbaum, F.A. Three-dimensional structure of full-length NtrX, an unusual member of the NtrC family of response regulators. J. Mol. Biol. 2017, 429, 1192–1212. [Google Scholar] [CrossRef] [PubMed]
- Bonato, P.; Alves, L.R.; Osaki, J.H.; Rigo, L.U.; Pedrosa, F.O.; Souza, E.M.; Zhang, N.; Schumacher, J.; Buck, M.; Wassem, R.; et al. The NtrY-NtrX two-component system is involved in controlling nitrate assimilation in Herbaspirillum seropedicae strain SmR1. FEBS J. 2016, 283, 3919–3930. [Google Scholar] [CrossRef] [PubMed]
- Calatrava-Morales, N.; Nogales, J.; Ameztoy, K.; van Steenbergen, B.; Soto, M.J. The NtrY/NtrX system of Sinorhizobium meliloti GR4 regulates motility, EPS I production and nitrogen metabolism but is dispensable for symbiotic nitrogen fixation. Mol. Plant. Microbe Interact. 2017, 30, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Robledo, M.; Peregrina, A.; Millan, V.; Garcia-Tomsig, N.I.; Torres-Quesada, O.; Mateos, P.F.; Becker, A.; Jimenez-Zurdo, J.I. A conserved alpha-proteobacterial small RNA contributes to osmoadaptation and symbiotic efficiency of rhizobia on legume roots. Environ. Microbiol. 2017, 19, 2661–2680. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Cho, B.C.; Simpson, A.G.B. Halocafeteria seosinensis gen. et. sp. nov. (bicosoecida), a halophilic bacterivorous nanoflagellate isolated from a solar saltern. Extremophiles 2006, 10, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Foissner, W.; Filker, S.; Stoeck, T. Schmidingerothrix salinarum nov spec. is the molecular sister of the large oxytrichid clade (ciliophora, hypotricha). J. Eukary Microbiol. 2014, 61, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Ventosa, A.; de la Haba, R.R.; Sanchez-Porro, C.; Papke, R.T. Microbial diversity of hypersaline environments: A metagenomic approach. Curr. Opin. Microbiol. 2015, 25, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Moreira, D.; Lopez-Garcia, P. Protist evolution: Stealing genes to gut it out. Curr. Biol. 2017, 27, R223–R225. [Google Scholar] [CrossRef] [PubMed]
- Rogozin, I.B.; Carmel, L.; Csuros, M.; Koonin, E.V. Origin and evolution of spliceosomal introns. Biol. Direct 2012, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Putnam, N.H.; Butts, T.; Ferrier, D.E.K.; Furlong, R.F.; Hellsten, U.; Kawashima, T.; Robinson-Rechavi, M.; Shoguchi, E.; Terry, A.; Yu, J.K.; et al. The amphioxus genome and the evolution of the chordate karyotype. Nature 2008, 453, 1064–1071. [Google Scholar] [CrossRef] [PubMed]
- Simakov, O.; Kawashima, T.; Marletaz, F.; Jenkins, J.; Koyanagi, R.; Mitros, T.; Hisata, K.; Bredeson, J.; Shoguchi, E.; Gyoja, F.; et al. Hemichordate genomes and deuterostome origins. Nature 2015, 527, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Husnik, F.; McCutcheon, J.P. Functional horizontal gene transfer from bacteria to eukaryotes. Nat. Rev. Microbiol. 2018, 16, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Landa, M.; Burns, A.S.; Roth, S.J.; Moran, M.A. Bacterial transcriptome remodeling during sequential co-culture with a marine dinoflagellate and diatom. ISMEJ 2017, 11, 2677–2690. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.; von Blohn, C.; Stanek, A.; Moses, S.; Barzantny, S.; Bremer, E. Synthesis, release, and recapture of the compatible solute proline by osmotically stressed Bacillus subtilis cells. Appl. Environ. Microbiol. 2012, 78, 5753–5762. [Google Scholar] [CrossRef] [PubMed]
- Börngen, K.; Battle, A.R.; Möker, N.; Morbach, S.; Marin, K.; Martinac, B.; Krämer, R. The properties and contribution of the Corynebacterium glutamicum MscS variant to fine-tuning of osmotic adaptation. Biochim. Biophys. Acta 2010, 1798, 2141–2149. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | ectC (in Total) | ectC (within ect Cluster) | ectC (Solitary) | ectD (within ect Cluster) | ectD (Separated from ect Cluster) | ask_ect (within ect Cluster) | ectR |
|---|---|---|---|---|---|---|---|
| Abundance | 582 | 437 | 145 | 259 | 68 | 133 | 97 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Czech, L.; Hermann, L.; Stöveken, N.; Richter, A.A.; Höppner, A.; Smits, S.H.J.; Heider, J.; Bremer, E. Role of the Extremolytes Ectoine and Hydroxyectoine as Stress Protectants and Nutrients: Genetics, Phylogenomics, Biochemistry, and Structural Analysis. Genes 2018, 9, 177. https://doi.org/10.3390/genes9040177
Czech L, Hermann L, Stöveken N, Richter AA, Höppner A, Smits SHJ, Heider J, Bremer E. Role of the Extremolytes Ectoine and Hydroxyectoine as Stress Protectants and Nutrients: Genetics, Phylogenomics, Biochemistry, and Structural Analysis. Genes. 2018; 9(4):177. https://doi.org/10.3390/genes9040177
Chicago/Turabian StyleCzech, Laura, Lucas Hermann, Nadine Stöveken, Alexandra A. Richter, Astrid Höppner, Sander H. J. Smits, Johann Heider, and Erhard Bremer. 2018. "Role of the Extremolytes Ectoine and Hydroxyectoine as Stress Protectants and Nutrients: Genetics, Phylogenomics, Biochemistry, and Structural Analysis" Genes 9, no. 4: 177. https://doi.org/10.3390/genes9040177
APA StyleCzech, L., Hermann, L., Stöveken, N., Richter, A. A., Höppner, A., Smits, S. H. J., Heider, J., & Bremer, E. (2018). Role of the Extremolytes Ectoine and Hydroxyectoine as Stress Protectants and Nutrients: Genetics, Phylogenomics, Biochemistry, and Structural Analysis. Genes, 9(4), 177. https://doi.org/10.3390/genes9040177