
The Umbrella Type Canopy Increases Tolerance to Abiotic Stress-Leaf Microenvironment Temperature and Tropospheric Ozone in ‘Chambourcin’

 and
and
Abstract
:1. Introduction
2. Materials and Methods
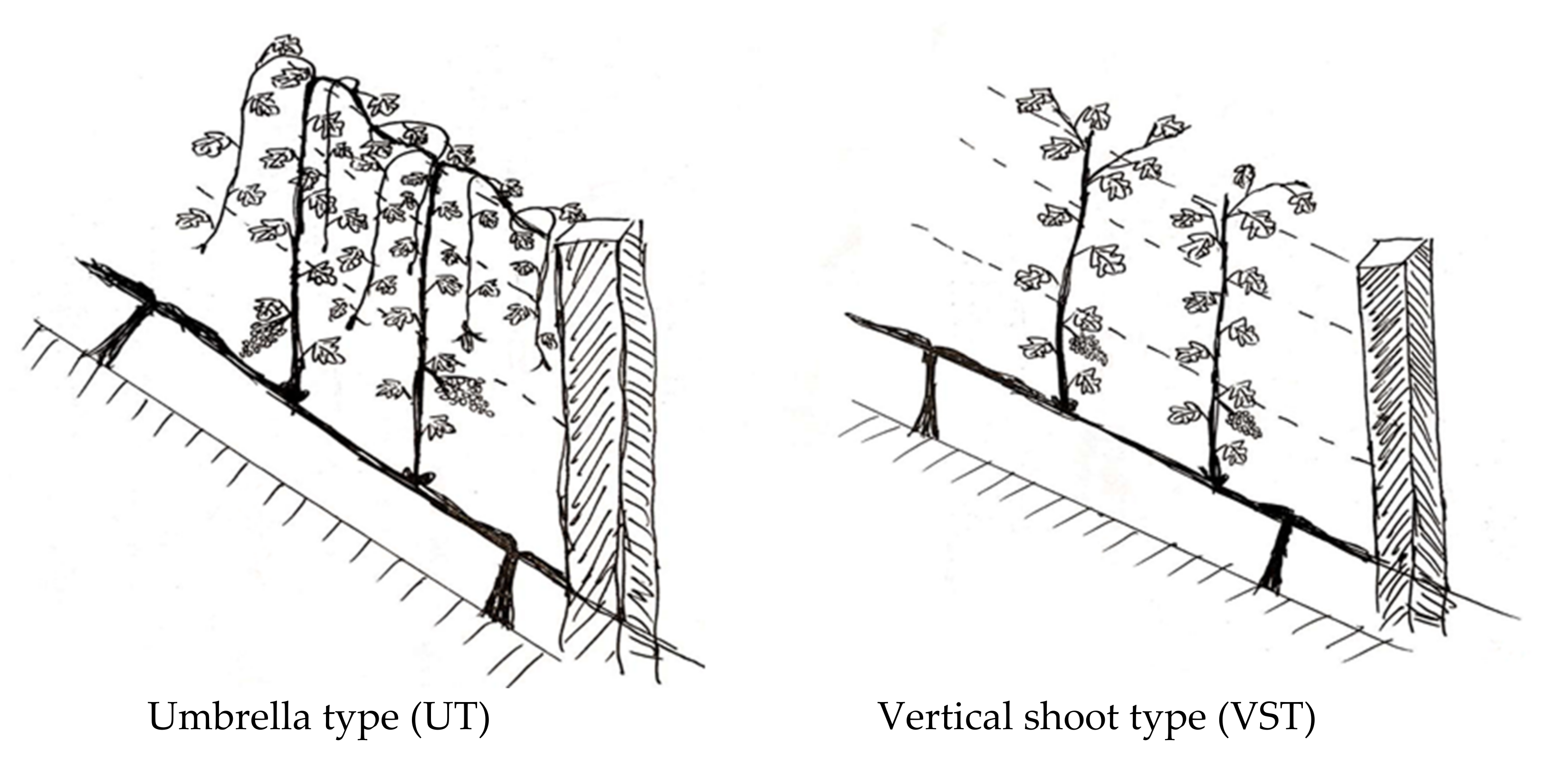
2.1. Materials and Canopy Management
2.2. Ozone Injury Classification
2.3. Monitoring of Temperature
2.4. Determination of the Leaf Area Index (LAI) and Diffuse Non-Interceptance (DIFN)
2.5. Determination of Chlorophyll Content in Grape Leaves
2.6. Determination of the Photosynthesis Rate
2.7. Determination of Light Transmittance and Space Photosynthetically Active Radiation
2.8. Berry Quality Indexes Determination
2.9. Determination of the Total Amount of Phenols, Flavonoids, Flavanols, and Anthocyanins
2.10. Statistical Analyses
3. Results
3.1. Effect of Canopy Types on the Temperature and Humidity around Grape Berries
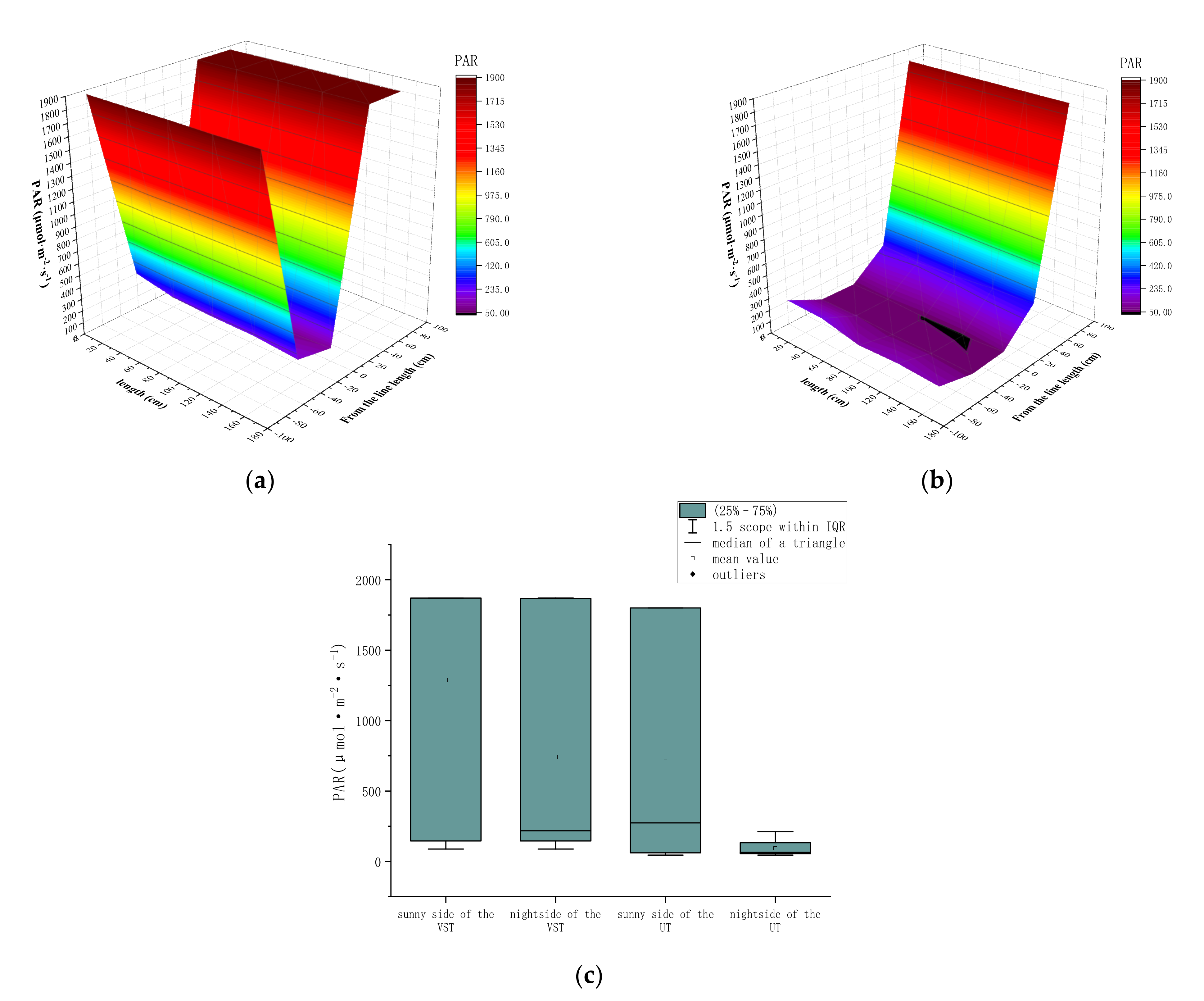
3.2. Effects of Canopy Types on the Spatial Distribution of Photosynthetic Active Radiation
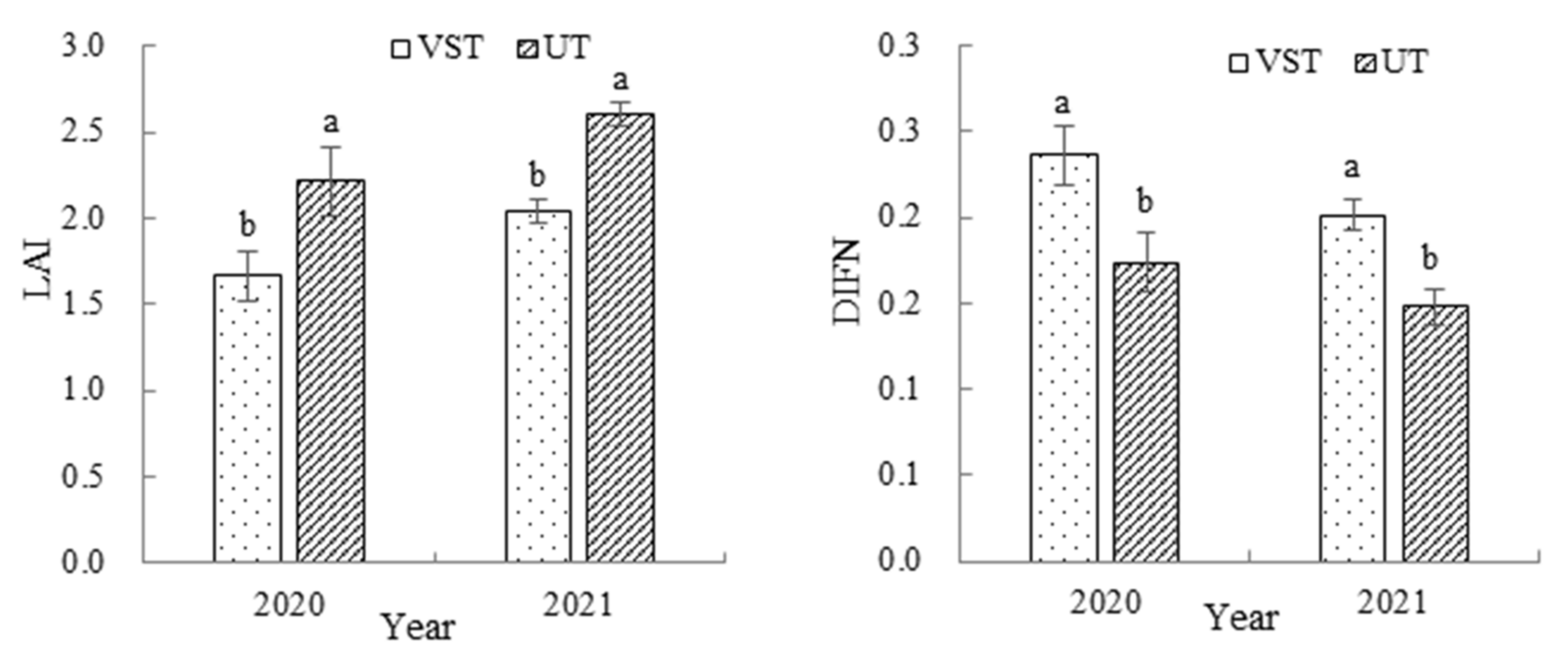
3.3. Effects of Canopy Types on the Leaf Area Index and Diffuse Non-Interceptance
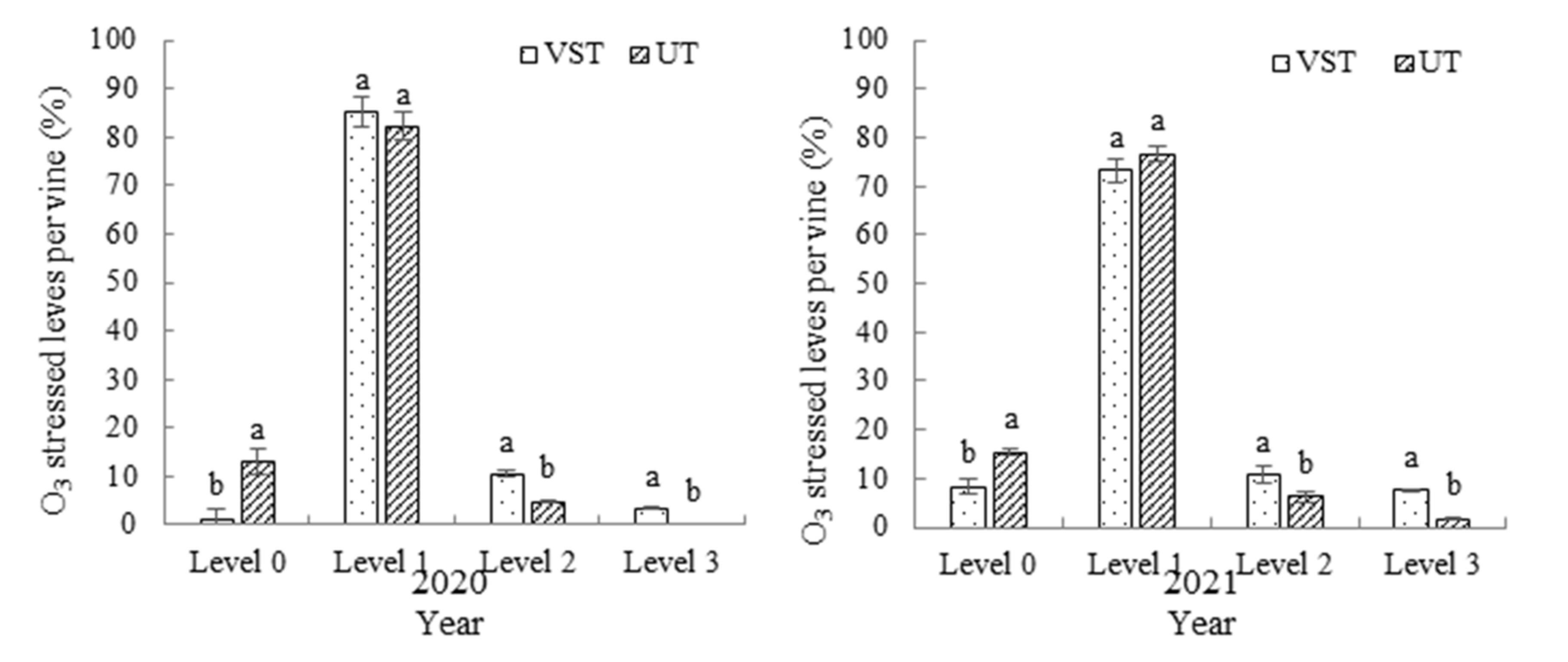
3.4. Effects of Different Canopy Types on Ozone Injury Symptoms in Grape Leaves
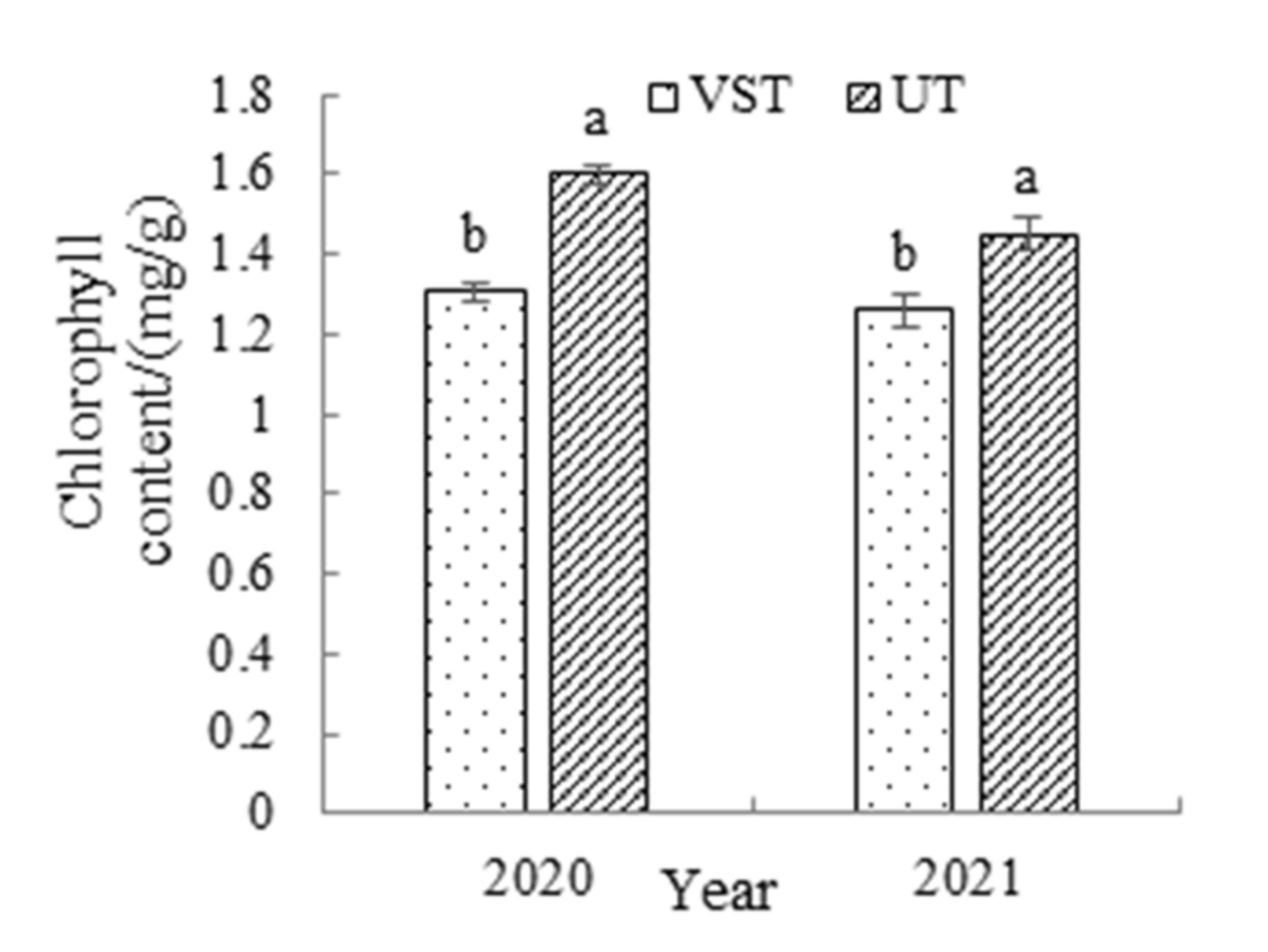
3.5. Effects of Different Canopy Types on the Chlorophyll Content
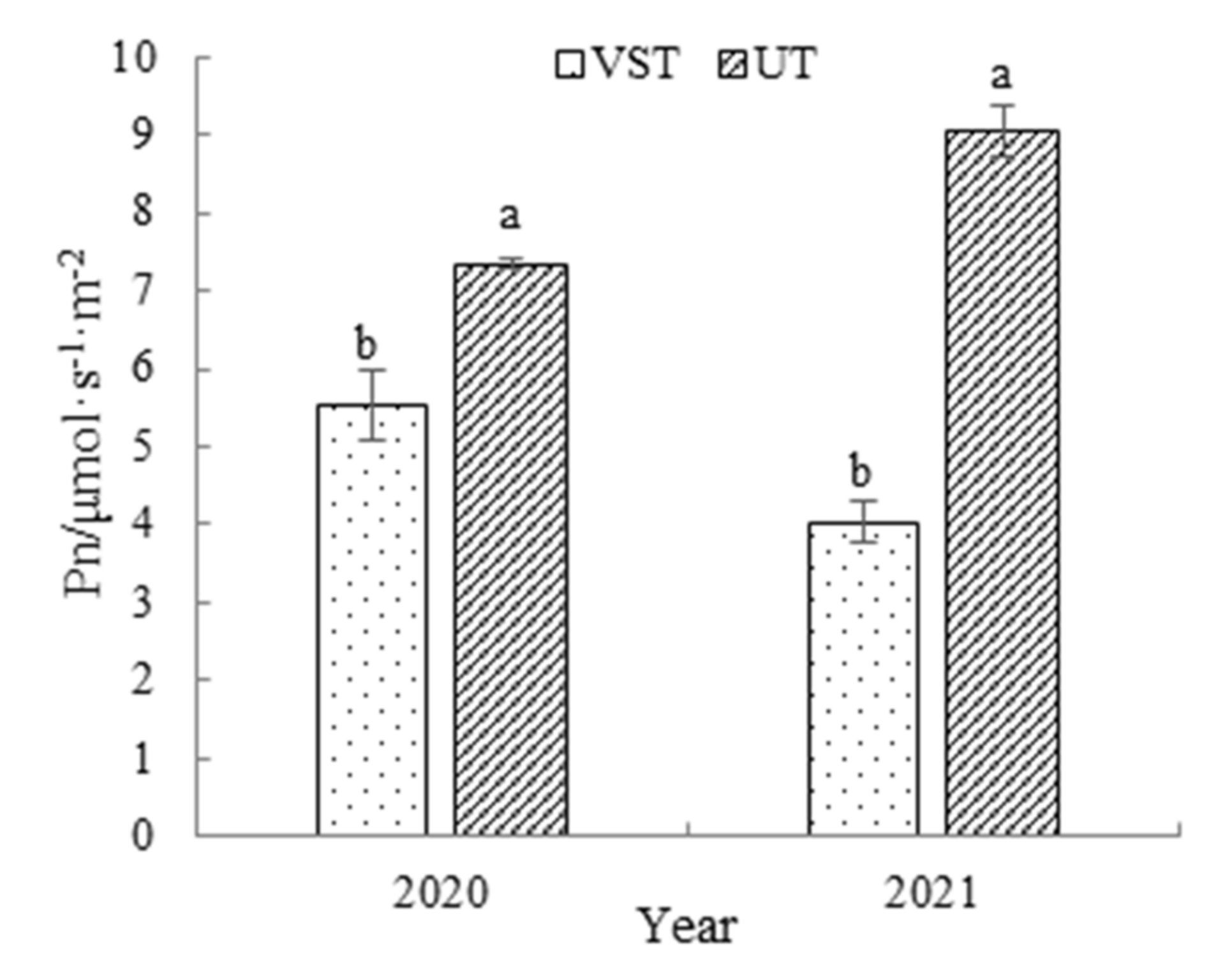
3.6. Effects of Canopy Types on the Net Leaf Photosynthetic Rate
3.7. Effects of Canopy Types on the Growth and Development of Grape Berries
3.8. Effects of Canopy Types on the Anthocyanins, Total Phenols, Flavonoids, and Flavanols of Grape Berries
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coia, L.R.; Ward, D.L. The hybrid grape Chambourcin has a role in quality red V. vinifera blends in a New World grape growing region. J. Wine Res. 2017, 28, 326–331. [Google Scholar] [CrossRef]
- Fan, D.; Wang, H.; Fu, C.; Zheng, Q.; Gao, Z.; Kang, H.; Du, Y. Comparison of leaf photosynthetic characteristics of pendulous canopy and horizontal canopy in Chambourcin grape. J. Fruit Sci. 2022, 39, 193–202. [Google Scholar]
- Antivilo, F.G.; Paz, R.C.; Keller, M.; Borgo, R.; Tognetti, J.; Juñent, F.R. Macro-and microclimate conditions may alter grapevine deacclimation: Variation in thermal amplitude in two contrasting wine regions from North and South America. Int. J. Biometeorol. 2017, 61, 2033–2045. [Google Scholar] [CrossRef] [PubMed]
- Conde, C.; Silva, P.; Fontes, N.; Dias, A.C.P.; Tavares, R.M.; Sousa, M.J.; Agasse, A.; Delrot, S.; Gerós, H. Biochemical changes throughout grape berry development and fruit and wine quality. Food 2007, 1, 1–22. [Google Scholar]
- Decoteau, D.R.; Marini, R.P.; Davis, D.D. Influence of ambient ozone on grape cultivars ‘Chambourcin’ and ‘Vidal’. J. Plant Sci. Res. 2019, 6, 1–5. [Google Scholar]
- Blanco-Ward, D.; Ribeiro, A.; Paoletti, E.; Miranda, A. Assessment of tropospheric ozone phytotoxic effects on the grapevine (Vitis vinifera L.): A review. Atmos. Environ. 2021, 244, 117924. [Google Scholar] [CrossRef]
- Geng, Q.-W.; Xing, H.; Sun, Y.-J.; Hao, G.-M.; Zhai, H.; Du, Y.-P. Analysis of the interaction effects of light and O3 on fluorescence properties of ‘Cabernet Sauvignon’grapes based on response surface methodology. Sci. Hortic. 2017, 225, 599–606. [Google Scholar] [CrossRef]
- Falcão, L.D.; Chaves, E.S.; Burin, V.M.; Falcão, A.P.; Gris, E.F.; Bonin, V.; Bordignon-Luiz, M.T. Maturity of Cabernet Sauvignon berries from grapevines grown with two different training systems in a new grape growing region in Brazil. Cienc. E Investig. Agrar. 2008, 35, 321–332. [Google Scholar] [CrossRef]
- White, R.; Vogel, A.; Scaduto, J.; Hickey, C. Evaluation of Canopy Division and Cane Pruning to Retrofit Spur-Pruned, Vertical Shoot-Positioned Petit Manseng. Catal. Discov. Into Pract. 2020, 4, 21–32. [Google Scholar] [CrossRef]
- Zoecklein, B.W.; Wolf, T.K.; Pélanne, L.; Miller, M.K.; Birkenmaier, S.S. Effect of vertical shoot-positioned, Smart-Dyson, and Geneva double-curtain training systems on Viognier grape and wine composition. Am. J. Enol. Vitic. 2008, 59, 11–21. [Google Scholar]
- Bavougian, C.M.; Read, P.E.; Walter-Shea, E. Training system effects on sunlight penetration, canopy structure, yield, and fruit characteristics of ‘Frontenac’grapevine (Vitis spp.). Int. J. Fruit Sci. 2012, 12, 402–409. [Google Scholar] [CrossRef]
- Reynolds, A.G.; Wardle, D.A.; Cliff, M.A.; King, M. Impact of training system and vine spacing on vine performance, berry composition, and wine sensory attributes of Seyval and Chancellor. Am. J. Enol. Vitic. 2004, 55, 84–95. [Google Scholar]
- Chevet, J.-M.; Lecocq, S.; Visser, M. Climate, grapevine phenology, wine production, and prices: Pauillac (1800–2009). Am. Econ. Rev. 2011, 101, 142–146. [Google Scholar] [CrossRef] [Green Version]
- Mosetti, D.; Herrera, J.; Sabbatini, P.; Green, A.; Alberti, G.; Peterlunger, E.; Lisjak, K.; Castellarin, S.D. Impact of leaf removal after berry set on fruit composition and bunch rot in’Sauvignon blanc’. VITIS-J. Grapevine Res. 2016, 55, 57–64. [Google Scholar]
- Rustioni, L.; Rocchi, L.; Guffanti, E.; Cola, G.; Failla, O. Characterization of grape (Vitis vinifera L.) berry sunburn symptoms by reflectance. J. Agric. Food Chem. 2014, 62, 3043–3046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Gao, Z.; Sun, Y.; Wang, Y.; Yao, Y.; Zhai, H.; Du, Y. Analyzing the grape leaf proteome and photosynthetic process provides insights into the injury mechanisms of ozone stress. Plant Growth Regul. 2020, 91, 143–155. [Google Scholar] [CrossRef]
- Candar, S.; Korkutal, I.; Bahar, E. Effect of canopy microclimate on Merlot (Vitis vinifera L.) grape composition. Appl. Ecol. Environ. Res. 2019, 17, 15431–15446. [Google Scholar] [CrossRef]
- Liu, X.; Song, Y.; Liu, Z.; Zhai, H. Effect of vertical and horizontal canopy on the secondary metabolites in’Moldova’grape. J. Fruit Sci. 2019, 36, 308–317. [Google Scholar]
- Lorenzini, G.; Nali, C.; Dota, M.R.; Martorana, F. Visual assessment of foliar injury induced by ozone on indicator tobacco plants: A data quality evaluation. Environ. Monit. Assess. 2000, 62, 175–191. [Google Scholar] [CrossRef]
- Gordon, R.; Brown, D.; Dixon, M. Non-destructive estimation of potato leaf area index using a fish-eye radiometer. Potato Res. 1994, 37, 393–402. [Google Scholar] [CrossRef]
- Welles, J.M.; Cohen, S. Canopy structure measurement by gap fraction analysis using commercial instrumentation. J. Exp. Bot. 1996, 47, 1335–1342. [Google Scholar] [CrossRef]
- Ling, Q.; Huang, W.; Jarvis, P. Use of a SPAD-502 meter to measure leaf chlorophyll concentration in Arabidopsis thaliana. Photosynth. Res. 2011, 107, 209–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.-L.; Wang, C.-R.; Li, X.-Y.; Yao, Y.-X.; Hao, Y.-J. Modifications of Kyoho grape berry quality under long-term NaCl treatment. Food Chem. 2013, 139, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Ignelzi, S.; Shirley, J. A comparison of methods for quantifying oligomeric proanthocyanidins from grape seed extracts. Am. J. Enol. Vitic. 2000, 51, 383–389. [Google Scholar]
- Orak, H.H. Total antioxidant activities, phenolics, anthocyanins, polyphenoloxidase activities of selected red grape cultivars and their correlations. Sci. Hortic. 2007, 3, 235–241. [Google Scholar] [CrossRef]
- Grappadelli, L.C. Early Season Patterns of Carbohydrate Partitioning in Exposed and Shaded Apple Branches. J. Amer. Soc. Hort. Sci 1994, 119, 596–603. [Google Scholar] [CrossRef] [Green Version]
- Booker, F.; Muntifering, R.; McGrath, M.; Burkey, K.; Decoteau, D.; Fiscus, E.; Manning, W.; Krupa, S.; Chappelka, A.; Grantz, D. The ozone component of global change: Potential effects on agricultural and horticultural plant yield, product quality and interactions with invasive species. J. Integr. Plant Biol. 2009, 51, 337–351. [Google Scholar] [CrossRef]
- Da Silva, D.; Han, L.; Faivre, R.; Costes, E. Influence of the variation of geometrical and topological traits on light interception efficiency of apple trees: Sensitivity analysis and metamodelling for ideotype definition. Ann. Bot. 2014, 114, 739–752. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Ma, X.; Ma, D.; Shi, J.; Hussain, S.; Han, M.; Costes, E.; Zhang, D. Modeling canopy photosynthesis and light interception partitioning among shoots in bi-axis and single-axis apple trees (Malus domestica Borkh.). Trees 2021, 35, 845–861. [Google Scholar] [CrossRef]
- Tang, L.; Yin, D.; Chen, C.; Yu, D.; Han, W. Optimal design of plant canopy based on light interception: A case study with loquat. Front. Plant Sci. 2019, 10, 364. [Google Scholar] [CrossRef]
- Long, S.P.; ZHU, X.G.; Naidu, S.L.; Ort, D.R. Can improvement in photosynthesis increase crop yields? Plant Cell Environ. 2006, 29, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Cominelli, E.; Gusmaroli, G.; Allegra, D.; Galbiati, M.; Wade, H.K.; Jenkins, G.I.; Tonelli, C. Expression analysis of anthocyanin regulatory genes in response to different light qualities in Arabidopsis thaliana. J. Plant Physiol. 2008, 165, 886–894. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortell, J.M.; Kennedy, J.A. Effect of shading on accumulation of flavonoid compounds in (Vitis vinifera L.) pinot noir fruit and extraction in a model system. J. Agric. Food Chem. 2006, 54, 8510–8520. [Google Scholar] [CrossRef] [PubMed]
- Wagner, G.; Loewus, F.A. L-ascorbic acid metabolism in vitaceae: Conversion to (+)-tartaric acid and hexoses. Plant Physiol. 1974, 54, 784–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergqvist, J.; Dokoozlian, N.; Ebisuda, N. Sunlight exposure and temperature effects on berry growth and composition of Cabernet Sauvignon and Grenache in the Central San Joaquin Valley of California. Am. J. Enol. Vitic. 2001, 52, 1–7. [Google Scholar]
- Downey, M.O.; Harvey, J.S.; Robinson, S.P. The effect of bunch shading on berry development and flavonoid accumulation in Shiraz grapes. Aust. J. Grape Wine Res. 2004, 10, 55–73. [Google Scholar] [CrossRef]
- Fujita, A.; Soma, N.; Goto-Yamamoto, N.; Shindo, H.; Kakuta, T.; Koizumi, T.; Hashizume, K. Anthocyanidin reductase gene expression and accumulation of flavan-3-ols in grape berry. Am. J. Enol. Vitic. 2005, 56, 336–342. [Google Scholar]
- Spayd, S.E.; Tarara, J.M.; Mee, D.L.; Ferguson, J. Separation of sunlight and temperature effects on the composition of Vitis vinifera cv. Merlot berries. Am. J. Enol. Vitic. 2002, 53, 171–182. [Google Scholar]
- Azuma, A.; Yakushiji, H.; Koshita, Y.; Kobayashi, S. Flavonoid biosynthesis-related genes in grape skin are differentially regulated by temperature and light conditions. Planta 2012, 236, 1067–1080. [Google Scholar] [CrossRef]
- Koyama, K.; Ikeda, H.; Poudel, P.R.; Goto-Yamamoto, N. Light quality affects flavonoid biosynthesis in young berries of Cabernet Sauvignon grape. Phytochemistry 2012, 78, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Matus, J.T.; Loyola, R.; Vega, A.; Peña-Neira, A.; Bordeu, E.; Arce-Johnson, P.; Alcalde, J.A. Post-veraison sunlight exposure induces MYB-mediated transcriptional regulation of anthocyanin and flavonol synthesis in berry skins of Vitis vinifera. J. Exp. Bot. 2009, 60, 853–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Pan, Q.; Jin, Z.; Mu, L.; Duan, C. Comparison on phenolic compounds in Vitis vinifera cv. Cabernet Sauvignon wines from five wine-growing regions in China. Food Chem. 2011, 125, 77–83. [Google Scholar] [CrossRef]
- Reynolds, A.G.; Heuvel, J.E.V. Influence of grapevine training systems on vine growth and fruit composition: A review. Am. J. Enol. Vitic. 2009, 60, 251–268. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period | Canopy Shape | Highest Temperature (°C) | Minimum Temperature (°C) | Day and Night Temperature Difference (°C) | ≥35 °C (%) | Humidity 60–80% (%) | Humidity >80% (%) |
|---|---|---|---|---|---|---|---|
| 2020 August | VST | 40.80 | 13.30 | 27.50 | 1.93 | 22.04 | 48.58 |
| UT | 39.90 | 13.20 | 26.70 | 1.80 | 22.73 | 51.37 | |
| 2021 August | VST | 37.40 | 15.30 | 22.10 | 3.94 | 15.51 | 67.52 |
| UT | 37.10 | 15.00 | 21.80 | 0.93 | 12.19 | 77.01 |
| Canopy Shape | Hundred-Grain Weight (g) | Soluble Solids (%) | Titratable Acid (g/L) | |
|---|---|---|---|---|
| 2020 | VST | 251.83 ± 5.35 b | 18.70 ± 1.33 b | 10.03 ± 0.64 a |
| UT | 264.29 ± 5.36 a | 20.97 ± 1.52 a | 10.05 ± 0.75 a | |
| 2021 | VST | 253.23 ± 2.49 b | 19.27 ± 0.23 a | 10.58 ± 0.68 b |
| UT | 258.04 ± 2.28 a | 19.77 ± 0.23 a | 11.53 ± 0.83 a |
| Canopy Shape | Anthocyanin (mg/g) | Total Phenol (mg/g) | Flavonoids (mg/g) | Flavanols (mg/g) | |
|---|---|---|---|---|---|
| 2020 | VST | 7.93 ± 0.04 b | 3.69 ± 0.12 b | 6.10 ± 0.49 b | 10.56 ± 0.72 b |
| UT | 10.51 ± 0.84 a | 5.93 ± 0.46 a | 10.44 ± 0.14 a | 20.26 ± 1.61 a | |
| 2021 | VST | 4.24 ± 0.22 b | 1.44 ± 0.03 b | 5.97 ± 0.28 a | 13.41 ± 0.54 a |
| UT | 6.45 ± 0.27 a | 1.55 ± 0.06 a | 6.25 ± 0.23 a | 14.85 ± 0.71 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Li, S.; Zhang, Y.; Huang, W.; Zhu, H.; Zhai, H.; Gao, Z.; Du, Y. The Umbrella Type Canopy Increases Tolerance to Abiotic Stress-Leaf Microenvironment Temperature and Tropospheric Ozone in ‘Chambourcin’. Atmosphere 2022, 13, 823. https://doi.org/10.3390/atmos13050823
Li X, Li S, Zhang Y, Huang W, Zhu H, Zhai H, Gao Z, Du Y. The Umbrella Type Canopy Increases Tolerance to Abiotic Stress-Leaf Microenvironment Temperature and Tropospheric Ozone in ‘Chambourcin’. Atmosphere. 2022; 13(5):823. https://doi.org/10.3390/atmos13050823
Chicago/Turabian StyleLi, Xinfeng, Shangrui Li, Yifan Zhang, Wenwei Huang, Huaping Zhu, Heng Zhai, Zhen Gao, and Yuanpeng Du. 2022. "The Umbrella Type Canopy Increases Tolerance to Abiotic Stress-Leaf Microenvironment Temperature and Tropospheric Ozone in ‘Chambourcin’" Atmosphere 13, no. 5: 823. https://doi.org/10.3390/atmos13050823
APA StyleLi, X., Li, S., Zhang, Y., Huang, W., Zhu, H., Zhai, H., Gao, Z., & Du, Y. (2022). The Umbrella Type Canopy Increases Tolerance to Abiotic Stress-Leaf Microenvironment Temperature and Tropospheric Ozone in ‘Chambourcin’. Atmosphere, 13(5), 823. https://doi.org/10.3390/atmos13050823