
Effects of Climate Change on Forest Regeneration in Central Spain


Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Vilalta, J.; Lloret, F.; Breshears, D.D. Drought-induced forest decline: Causes, scope and implications. Biol. Lett. 2012, 8, 689–691. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Lloret, F.; Montoya, R. Severe drought effects on Mediterranean woody flora in Spain. For. Sci. 2001, 47, 214–218. [Google Scholar] [CrossRef]
- Landmann, G.; Dreyer, E. Impacts of drought and heat on forests. Synthesis of available knowledge, with emphasis on the 2003 event in Europe. Ann. For. Sci. 2006, 63, 567–568. [Google Scholar] [CrossRef] [Green Version]
- Pardos, M.; Madrigal, G.; de Dios-García, J.; Gordo, J.; Calama, R. Sapling recruitment in mixed stands in the Northern Plateau of Spain: A patch model approach. Trees-Struct. Funct. 2021, 35, 2043–2058. [Google Scholar] [CrossRef]
- Marañón, T.; Zamora, R.; Villar, R.; Zavala, M.A.; Quero, J.L.; Pérez-Ramos, I.; Mendoza, I.; Castro, J. Regeneration of tree species and restoration under constrasted Mediterranean habitats: Field and glasshouse experiments. Int. J. Ecol. Environ. Sci. 2004, 30, 187–196. [Google Scholar]
- Matías, L.; Zamora, R.; Castro, J. Sporadic rainy events are more critical than increasing of drought intensity for woody species recruitment in a Mediterranean community. Oecologia 2012, 169, 833–844. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Camarero, J.J.; Dobbertin, M.; Fernández-Cancio, Á.; Vilà-Cabrera, A.; Manzanedo, R.D.; Zavala, M.A.; Navarro-Cerrillo, R.M. Contrasting vulnerability and resilience to drought-induced decline of densely planted vs. natural rear-edge Pinus nigra forests. For. Ecol. Manag. 2013, 310, 956–967. [Google Scholar] [CrossRef]
- Marqués, L.; Madrigal-González, J.; Zavala, M.A.; Camarero, J.J.; Hartig, F. Last-century forest productivity in a managed dry-edge Scots pine population: The two sides of climate warming. Ecol. Appl. 2018, 28, 95–105. [Google Scholar] [CrossRef]
- Hogg, E.H.; Schwarz, A.G. Regeneration of planted conifers across climatic moisture gradients on the Canadian prairies: Implications for distribution and climate change. J. Biogeogr. 1997, 24, 527–534. [Google Scholar] [CrossRef]
- Dulamsuren, C.; Wommelsdorf, T.; Zhao, F.; Xue, Y.; Zhumadilov, B.Z.; Leuschner, C.; Hauck, M. Increased summer temperatures reduce the growth and regeneration of Larix sibirica in southern boreal forests of Eastern Kazakhstan. Ecosystems 2013, 16, 1536–1549. [Google Scholar] [CrossRef]
- Nardini, A.; Lo Gullo, M.A.; Trifilò, P.; Salleo, S. The challenge of the Mediterranean climate to plant hydraulics: Responses and adaptations. Environ. Exp. Bot. 2014, 103, 68–79. [Google Scholar] [CrossRef]
- Calama, R.; Manso, R.; Lucas-Borja, M.; Espelta, J.; Piqué, M.; Bravo, F.; del Peso, C.; Pardos, M. Natural regeneration in Iberian pines: A review of dynamic processes and proposals for management. For. Syst. 2017, 26, eR02S. [Google Scholar] [CrossRef]
- Castro, J.; Zamora, R.; Hódar, J.A.; Gómez, J.M. Seedling establishment of a boreal tree species (Pinus sylvestris) at its southernmost distribution limit: Consequences of being in a marginal Mediterranean habitat. J. Ecol. 2004, 92, 266–277. [Google Scholar] [CrossRef]
- Benavides, R.; Escudero, A.; Coll, L.; Ferrandis, P.; Ogaya, R.; Gouriveau, F.; Peñuelas, J.; Valladares, F. Recruitment patterns of four tree species along elevation gradients in Mediterranean mountains: Not only climate matters. For. Ecol. Manag. 2016, 360, 287–296. [Google Scholar] [CrossRef] [Green Version]
- Karavani, A.; Boer, M.M.; Baudena, M.; Colinas, C.; Díaz-Sierra, R.; Pemán, J.; Luis, M.; Enríquez-de-Salamanca, Á.; Resco, V. Fire induced deforestation in drought-prone Mediterranean forests: Drivers and unknowns from leaves to communities. Ecol. Monogr. 2018, 88, 141–169. [Google Scholar] [CrossRef] [Green Version]
- Enríquez-de-Salamanca, Á. Dynamics of mediterranean pine forests reforested after fires. J. For. Res. 2022. [CrossRef]
- Gazol, A.; Camarero, J.J.; Sangüesa-Barreda, G.; Vicente-Serrano, S.M. Post-drought resilience after forest die-off: Shifts in regeneration, composition, growth and productivity. Front. Plant Sci. 2018, 871, 1546. [Google Scholar] [CrossRef]
- Mutke, S.; Gordo, J.; Gil, L. Variability of Mediterranean stone pine cone production: Yield loss as response to climate change. Agric. For. Meteorol. 2005, 132, 263–272. [Google Scholar] [CrossRef]
- Gentilesca, T.; Colangelo, M.; Nolè, A.; Ripullone, F.; Camarero, J.J. Drought-induced oak decline in the western Mediterranean region: An overview on current evidences, mechanisms and management options to improve forest resilience. iForest 2017, 10, 796–806. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Fayos, P.; Monleon, V.J.; Espigares, T.; Nicolau, J.M.; Bochet, E. Increasing aridity threatens the sexual regeneration of Quercus ilex (holm oak) in Mediterranean ecosystems. PLoS ONE 2020, 15, e0239755. [Google Scholar] [CrossRef] [PubMed]
- Plieninger, T.; Rolo, V.; Moreno, G. Large-scale patterns of Quercus ilex, Quercus suber, and Quercus pyrenaica regeneration in Central-Western Spain. Ecosystems 2010, 13, 644–660. [Google Scholar] [CrossRef]
- Lloret, F.; Siscart, D.; Dalmases, C. Canopy recovery after drought dieback in holm-oak Mediterranean forests of Catalonia (NE Spain). Glob. Chang. Biol. 2004, 10, 2092–2099. [Google Scholar] [CrossRef]
- Ibáñez, B.; Gómez-Aparicio, L.; Stoll, P.; Ávila, J.M.; Pérez-Ramos, I.M.; Marañón, T. A Neighborhood analysis of the consequences of Quercus suber decline for regeneration dynamics in Mediterranean forests. PLoS ONE 2015, 10, e0117827. [Google Scholar] [CrossRef] [Green Version]
- Saura-Mas, S.; Bonas, A.; Lloret, F. Plant community response to drought-induced canopy defoliation in a Mediterranean Quercus ilex forest. Eur. J. For. Res. 2015, 134, 261–272. [Google Scholar] [CrossRef]
- Calama, R.; Montero, G. Cone and seed production from stone pine (Pinus pinea L.) stands in Central Range (Spain). Eur. J. Forest. Res. 2017, 126, 23–35. [Google Scholar] [CrossRef]
- Lloret, F.; Peñuelas, J.; Prieto, P.; Llorens, L.; Estiarte, M. Plant community changes induced by experimental climate change: Seedling and adult species composition. Perspect. Plant Ecol. Evol. Syst. 2009, 11, 53–63. [Google Scholar] [CrossRef]
- Pérez-Ramos, I.M.; Marañón, T. Community-level seedling dynamics in Mediterranean forests: Uncoupling between the canopy and the seedling layers. J. Veg. Sci. 2012, 23, 526–540. [Google Scholar] [CrossRef] [Green Version]
- WMO. State of the Global Climate 2021; World Meteorological Organization: Gèneve, Switzerland, 2022; Available online: https://library.wmo.int/doc_num.php?explnum_id=11178 (accessed on 5 June 2022).
- Gea-Izquierdo, G.; Montes, F.; Gavilán, R.G.; Cañellas, I.; Rubio, A. Is this the end? Dynamics of a relict stand from pervasively deforested ancient Iberian pine forests. Eur. J. For. Res. 2015, 134, 525–536. [Google Scholar] [CrossRef]
- Pardos, M.; Montes, F.; Aranda, I.; Cañellas, I. Influence of environmental conditions on germinant survival and diversity of Scots pine (Pinus sylvestris L.) in central Spain. Eur. J. For. Res. 2007, 126, 37–47. [Google Scholar] [CrossRef]
- Manso, R.; Calama, R.; Madrigal, G.; Pardos, M. A silvicultureoriented spatio-temporal model for germination in Pinus pinea L. in the Spanish Northern Plateau based on a direct seeding experiment. Eur. J. For. Res. 2013, 132, 969–982. [Google Scholar] [CrossRef] [Green Version]
- Manso, R.; Pukkala, T.; Pardos, M.; Miina, J.; Calama, R. Modelling Pinus pinea forest management to attain natural regeneration under present and future climatic scenarios. Can. J. For. Res. 2014, 44, 250–262. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Fernández, D.; Montes, F.; Sánchez-González, M.; Gordo, F.J.; Cañellas, I. Regeneration dynamics of mixed stands of Pinus pinaster Ait. and Pinus pinea L. in Central Spain. Eur. J. For. Res. 2018, 137, 17–27. [Google Scholar] [CrossRef]
- WMO. Guide to Climatological Practices; World Meteorological Organization, Gèneve, Switzerland. 2018. Available online: https://library.wmo.int/doc_num.php?explnum_id=5541 (accessed on 5 June 2022).
- AEMET. AEMET OpenData. Agencia Estatal de Meteorología. 2022. Available online: https://opendata.aemet.es/centrodedescargas/inicio (accessed on 7 June 2022).
- Thornthwaite, C.W.; Mather, J.R. The water balance. Publ. Climatol. 1955, 8, 5–86. [Google Scholar]
- Thornthwaite, C.W.; Mather, J.R. Instructions for evaluating the water balance. Publ. Climatol. 1957, 10, 185–204. [Google Scholar]
- Gaussen, H.; Bagnouls, F. Dry season and xerothermic index. Bull. Soc. Hist. Nat. Toulouse 1953, 88, 193–240. [Google Scholar]
- Thornthwaite, C.W. An approach toward a rational classification of climate. Geogr. Rev. 1948, 38, 55–94. [Google Scholar] [CrossRef]
- Viñegla, B.; Lechuga, V.; Linares, J.C. Natural regeneration and drought effects in the Mediterranean basin. In Forest Management of Mediterranean Forests under the New Context of Climate Change: Building Alternatives for the Coming Future; Lucas-Borja, M.E., Ed.; Nova Science Publishers: New York, NY, USA, 2013; pp. 53–69. [Google Scholar]
- Will, R.E.; Wilson, S.M.; Zou, C.B.; Hennessey, T.C. Increased vapor pressure deficit due to higher temperature leads to greater transpiration and faster mortality during drought for tree seedlings common to the forest–grassland ecotone. New Phytol. 2013, 200, 366–374. [Google Scholar] [CrossRef]
- Broz, A.; Retallack, G.J.; Maxwell, T.M.; Silva, L.C.R. A record of vapour pressure deficit preserved in wood and soil across biomes. Sci. Rep. 2021, 11, 662. [Google Scholar] [CrossRef]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.W.; Sperry, J.S.; McDowell, N.G. Plant responses to rising vapor pressure deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Qin, Z.; Lin, S.; Chen, X.; Chen, B.; He, B.; Wei, J.; Yuan, W. Large influence of atmospheric vapor pressure deficit on ecosystem production efficiency. Nat. Commun. 2022, 13, 1653. [Google Scholar] [CrossRef] [PubMed]
- AdapteCCa. Visor de escenarios de cambio climático. Plataforma Sobre Adaptación al Cambio Climático en España. Ministerio para la Transición Ecológica y el Reto Demográfico. 2022. Available online: https://escenarios.adaptecca.es (accessed on 8 June 2022).
- Williams, A.P.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D.; et al. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Chang. 2013, 3, 292–297. [Google Scholar] [CrossRef]
- Martinez-Vilalta, J.; Piñol, J. Drought-induced mortality and hydraulic architecture in pine populations of the NE Iberian Peninsula. For. Ecol. Manag. 2002, 161, 247–256. [Google Scholar] [CrossRef]
- Fernández, A.; Navarro, R.M.; Sánchez, R.; Fernández, R.; Manrique, E. Viabilidad fitoclimática de las repoblaciones de pino silvestre (Pinus sylvestris L.) en la Sierra de los Filabres (Almería). Ecosistemas 2011, 20, 124–144. [Google Scholar]
- Sánchez de Dios, R.; Benito-Garzón, M.; Sainz-Ollero, H. Present and future extension of the Iberian submediterranean territories as determined from the distribution of marcescent oaks. Plant. Ecol. 2009, 204, 189–205. [Google Scholar] [CrossRef]
- Rubio-Cuadrado, Á.; Camarero, J.J.; Aspizua, R.; Sánchez-González, M.; Gil, L.; Montes, F. Abiotic factors modulate post-drought growth resilience of Scots pine plantations and rear-edge Scots pine and oak forests. Dendrochronologia 2018, 51, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Peñuelas, J.; Ogaya, R.; Boada, M.; Jump, A.S. Migration, invasion and decline: Changes in recruitment and forest structure in a warming-linked shift of European beech forest in Catalonia (NE Spain). Ecography 2007, 30, 829–837. [Google Scholar] [CrossRef]
- Mendoza, I.; Zamora, R.; Castro, J. A seeding experiment for testing tree-community recruitment under variable environments: Implications for forest regeneration and conservation in Mediterranean habitats. Biol. Conserv. 2009, 142, 1491–1499. [Google Scholar] [CrossRef]
- Chirino, E.; Vilagrosa, A.; Cortina, J.; Valdecantos, A.; Fuentes, D.; Trubat, R.; Luis, V.C.; Puértolas, J.; Bautista, S.; Baeza, M.J.; et al. Ecological restoration in degraded drylands: The need to improve the seedling quality and site conditions in the field. In Forest Management; Grossberg, S.P., Ed.; Nova Science Publishers: New York, NY, USA, 2009; pp. 85–158. [Google Scholar]
- Hlásny, T.; Mátyás, C.; Seidl, R.; Kulla, L.; Merganicová, K.; Trombik, J.; Dobor, L.; Barcza, Z.; Konôpka, B. Climate change increases the drought risk in central European forests: What are the options for adaptation? Lesn. Cas. For. J. 2014, 60, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Konnert, M.; Fady, B.; Gömöry, D.; A’Hara, S.; Wolter, F.; Ducci, F.; Koskela, J.; Bozzano, M.; Maaten, T.; Kowalczyk, J. Use and Transfer of Forest Reproductive Material in Europe in the Context of Climate Change; Euforgen, Bioversity International: Rome, Italy, 2015. [Google Scholar]
- Seidel, H.; Schunk, C.; Matiu, M.; Menzel, A. Diverging drought resistance of scots pine provenances revealed by infrared thermography. Front. Plant Sci. 2016, 7, 1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Code | Name | X | Y | Elevation | Period |
|---|---|---|---|---|---|
| 3195 | Madrid—Retiro | 442470 | 4473702 | 667 m | 1952–2021 |
| 3196 | Madrid—Cuatro Vientos | 433267 | 4469738 | 690 m | 1952–2021 |
| 3191E | Colmenar Viejo-FAMET | 435367 | 4505305 | 1004 m | 1978–2021 |
| 2465 | Segovia | 405190 | 4533294 | 1005 m | 1960–2021 |
| 2462 | Puerto de Navacerrada | 414745 | 4516276 | 1894 m | 1952–2021 |
| Parameter | Code | Period | Unit |
|---|---|---|---|
| Mean temperature | TA | Annual | °C |
| TS | July–August | °C | |
| TSE | June–September | °C | |
| Rainfall | RA | Annual | mm |
| RS | July–August | mm | |
| RSE | June–September | mm | |
| Potential evapotranspiration | PETA | Annual | mm |
| PETS | July–August | mm | |
| PETSE | June–September | mm | |
| Physiological drought | PDA | Annual | mm |
| PDS | July–August | mm | |
| PDSE | June–September | mm | |
| Summer drought | SD | Annual | days |
| Par | 3195 | 3196 | 3191E | 2465 | 2462 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| p | r | R2 | p | r | R2 | p | r | R2 | p | Corr | r | p | r | R2 | |
| SD | 0.4830 | 0.0852 | 0.7264 | 0.3520 | 0.1129 | 1.2750 | 0.6480 | 0.0708 | 0.5001 | 0.1552 | 0.1717 | 2.9482 | 0.4709 | 0.0933 | 0.8697 |
| TA | 0.0000 | 0.7921 | 62.7371 | 0.0000 | 0.8033 | 64.5347 | 0.0875 | 0.2606 | 6.7937 | 0.0000 | 0.7163 | 51.3114 | 0.0000 | 0.6194 | 38.3613 |
| TS | 0.0000 | 0.7527 | 56.6554 | 0.0000 | 0.7526 | 56.6352 | 0.0177 | 0.3561 | 12.6823 | 0.0032 | 0.3473 | 12.0624 | 0.0008 | 0.4140 | 17.1413 |
| TSE | 0.0000 | 0.7806 | 60.9265 | 0.0000 | 0.7705 | 59.3644 | 0.0110 | 0.3798 | 14.4259 | 0.0000 | 0.6699 | 44.8790 | 0.0002 | 0.4620 | 21.3438 |
| RA | 0.2638 | −0.1354 | 1.8325 | 0.0480 | −0.2372 | 5.6745 | 0.7948 | 0.0403 | 0.1628 | 0.1982 | −0.1557 | 2.4228 | 0.7554 | −0.0404 | 0.1630 |
| RS | 0.6845 | −0.0494 | 0.2443 | 0.5273 | −0.0768 | 0.5903 | 0.7365 | −0.0522 | 0.2723 | 0.9836 | 0.0025 | 0.0006 | 0.4111 | 0.1063 | 1.1292 |
| RSE | 0.1914 | −0.1580 | 2.4967 | 0.0696 | −0.2182 | 4.7613 | 0.3916 | −0.1324 | 1.7525 | 0.0099 | −0.3063 | 9.3802 | 0.2463 | −0.1495 | 2.2341 |
| PETA | 0.0000 | 0.7908 | 62.5410 | 0.0000 | 0.8029 | 64.4627 | 0.0000 | 0.5859 | 34.3324 | 0.0000 | 0.6123 | 37.4948 | 0.0000 | 0.5802 | 33.6659 |
| PETS | 0.0000 | 0.7417 | 55.0156 | 0.0000 | 0.7401 | 54.7804 | 0.0445 | 0.3045 | 9.2696 | 0.0000 | 0.4703 | 22.1158 | 0.0083 | 0.0332 | 11.0506 |
| PETSE | 0.0000 | 0.7735 | 59.8260 | 0.0000 | 0.7611 | 57.9236 | 0.0363 | 0.3165 | 10.0180 | 0.0000 | 0.6181 | 38.2090 | 0.0020 | 0.3850 | 14.8222 |
| PDA | 0.0005 | 0.4027 | 16.2135 | 0.0002 | 0.4258 | 18.1314 | 0.5348 | 0.0961 | 0.9242 | 0.0025 | 0.3556 | 12.6419 | 0.1916 | 0.1681 | 2.8252 |
| PDS | 0.0000 | 0.5510 | 30.3551 | 0.0000 | 0.5320 | 28.3032 | 0.1396 | 0.2263 | 5.1233 | 0.0046 | 0.3351 | 11.2291 | 0.0000 | 0.5647 | 31.8839 |
| PDSE | 0.0000 | 0.4755 | 22.6131 | 0.0000 | 0.5333 | 28.4355 | 0.0953 | 0.2547 | 6.4845 | 0.0021 | 0.3610 | 13.0348 | 0.1002 | 0.2107 | 4.4411 |
| Period | Est | 1st Regression | 2nd Regression | 3rd Regression | 4rd Regression | Remain | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| p | R2adj | Out | p | R2adj | Out | p | R2adj | Out | p | R2adj | Out | |||
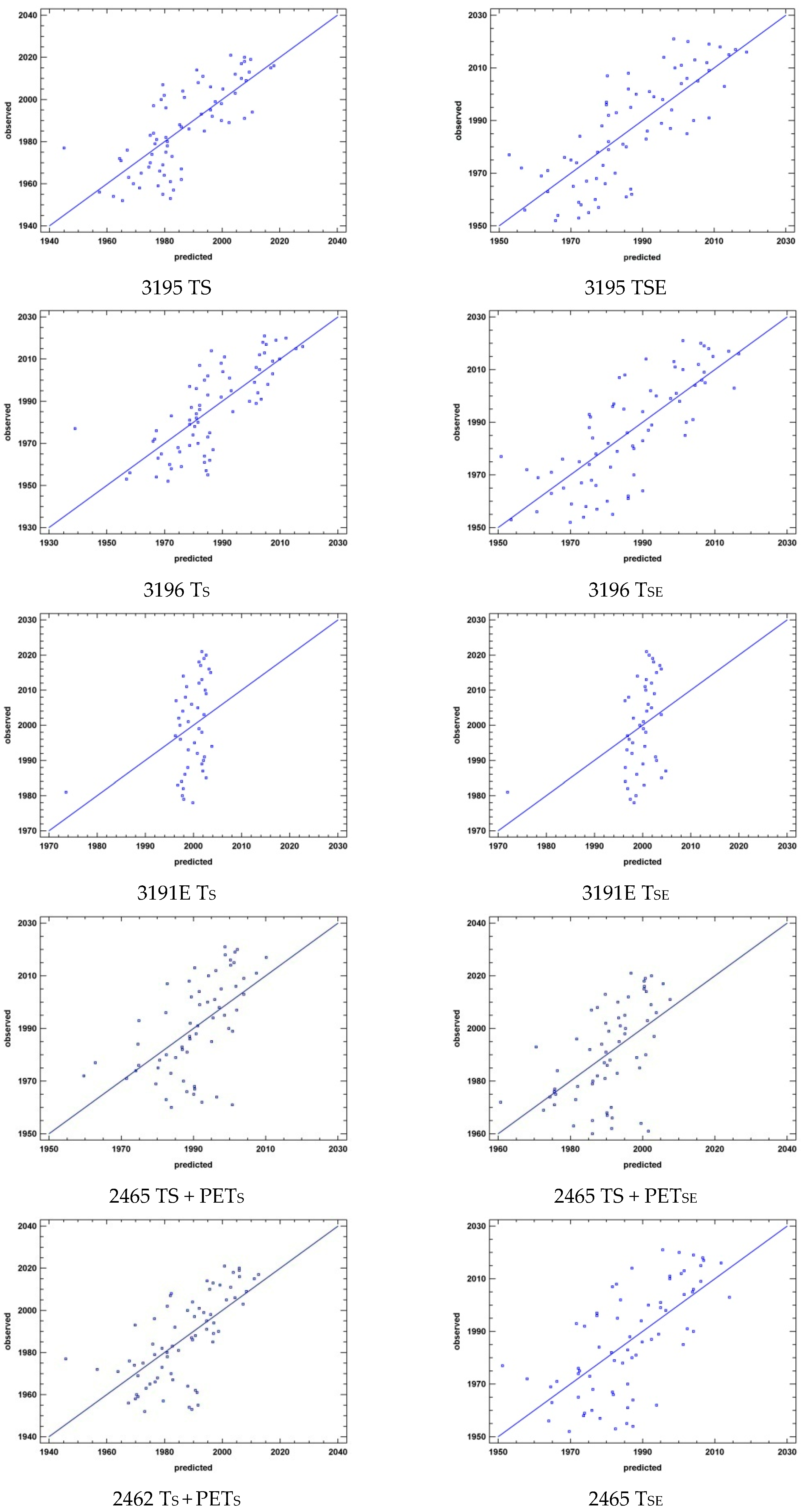
| Summer SD + ∑XS | 3195 | 0.0000 | 54.9726 | SD | 0.0000 | 55.6536 | PDS | 0.0000 | 56.2178 | RS | 0.0000 | 56.1576 | PETS | TS |
| 3196 | 0.0000 | 54.2578 | SD | 0.0000 | 54.9487 | PDS | 0.0000 | 55.6154 | RS | 0.0000 | 56.0413 | PETS | TS | |
| 3191E | 0.2872 | 3.2982 | RS | 0.1795 | 5.7636 | SD | 0.0967 | 8.0560 | PETS | 0.0444 | 9.9092 | PDS | TS | |
| 2465 | 0.0001 | 31.0426 | SD | 0.0000 | 32.0316 | RS | 0.0000 | 32.6101 | PDS | 0.0000 | 30.3015 | - | TS + PETS | |
| 2462 | 0.0000 | 43.8003 | SD | 0.0000 | 44.6218 | PDS | 0.0000 | 43.6710 | RS | 0.0000 | 43.2437 | - | TS + PETS | |
| Expanded summer SD + ∑XSE | 3195 | 0.0000 | 57.4294 | PDSE | 0.0000 | 58.0825 | PETSE | 0.0000 | 58.6507 | RSE | 0.0000 | 59.1684 | SD | TSE |
| 3196 | 0.0000 | 60.0179 | PETSE | 0.0000 | 60.5997 | RSE | 0.0000 | 61.1009 | PDSE | 0.0000 | 61.3526 | SD | TSE | |
| 3191E | 0.1071 | 10.0407 | PDSE | 0.0587 | 12.1870 | PETSE | 0.0388 | 12.6003 | SD | 0.0217 | 12.9929 | RSE | TSE | |
| 2465 | 0.0002 | 29.0314 | SD | 0.0001 | 30.2764 | RSE | 0.0001 | 27.3419 | PDSE | 0.0000 | 26.9267 | - | TSE + PETSE | |
| 2462 | 0.0000 | 42.6008 | RSE | 0.0000 | 43.4835 | PDSE | 0.0000 | 43.7984 | SD | 0.0000 | 44.1704 | PETSE | TSE | |
| Par | 1962–1981 | 1982–2001 | 2002–2021 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3195 | 3196 | 2465 | 2462 | 3195 | 3196 | 3191E | 2465 | 2462 | 3195 | 3196 | 3191E | 2465 | 2462 | |
| SD | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| TA | - | - | - | - | - | 0.0154 | - | - | - | 0.0256 | 0.0235 | 0.0200 | - | 0.0142 |
| TS | - | - | 0.0353 | - | - | - | - | - | - | 0.0042 | 0.0251 | 0.0165 | - | 0.0500 |
| TSE | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| RA | - | - | - | - | - | - | - | 0.029 | - | - | - | - | - | - |
| RS | - | - | - | - | - | - | - | - | - | - | 0.0307 | - | - | - |
| RSE | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| PETA | - | - | 0.0266 | - | - | 0.0098 | - | - | - | - | - | - | - | - |
| PETS | - | - | 0.0421 | - | - | - | - | - | - | 0.0048 | 0.0284 | 0.0271 | - | - |
| PETSE | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| PDA | - | - | - | - | - | - | - | 0.0196 | - | - | - | - | - | - |
| PDS | - | - | - | - | - | 0.0195 | - | - | - | - | - | - | - | - |
| PDSE | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Meteorological Station | Municipality | PET (mm/Month) | |||
|---|---|---|---|---|---|
| Scenario RCP 4.5 | Scenario RCP 8.5 | ||||
| 2021 | 2100 | 2021 | 2100 | ||
| 3195, 3196 | Madrid | 66.98 | 72.37 | 68.56 | 92.03 |
| 3191E | Colmenar Viejo | 57.47 | 62.60 | 58.70 | 70.17 |
| 2465 | Segovia | 63.41 | 69.99 | 64.67 | 78.79 |
| 2462 | San Ildefonso | 63.88 | 70.03 | 65.26 | 79.54 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Enríquez-de-Salamanca, Á. Effects of Climate Change on Forest Regeneration in Central Spain. Atmosphere 2022, 13, 1143. https://doi.org/10.3390/atmos13071143
Enríquez-de-Salamanca Á. Effects of Climate Change on Forest Regeneration in Central Spain. Atmosphere. 2022; 13(7):1143. https://doi.org/10.3390/atmos13071143
Chicago/Turabian StyleEnríquez-de-Salamanca, Álvaro. 2022. "Effects of Climate Change on Forest Regeneration in Central Spain" Atmosphere 13, no. 7: 1143. https://doi.org/10.3390/atmos13071143
APA StyleEnríquez-de-Salamanca, Á. (2022). Effects of Climate Change on Forest Regeneration in Central Spain. Atmosphere, 13(7), 1143. https://doi.org/10.3390/atmos13071143