Environmental Variability and Macrophyte Assemblages in Coastal Lagoon Types of Western Greece (Mediterranean Sea)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling Design of Water Quality and Aquatic Macrophytes
2.3. Statistical Analysis
3. Results
3.1. Environmental Change and Water Quality Characteristics
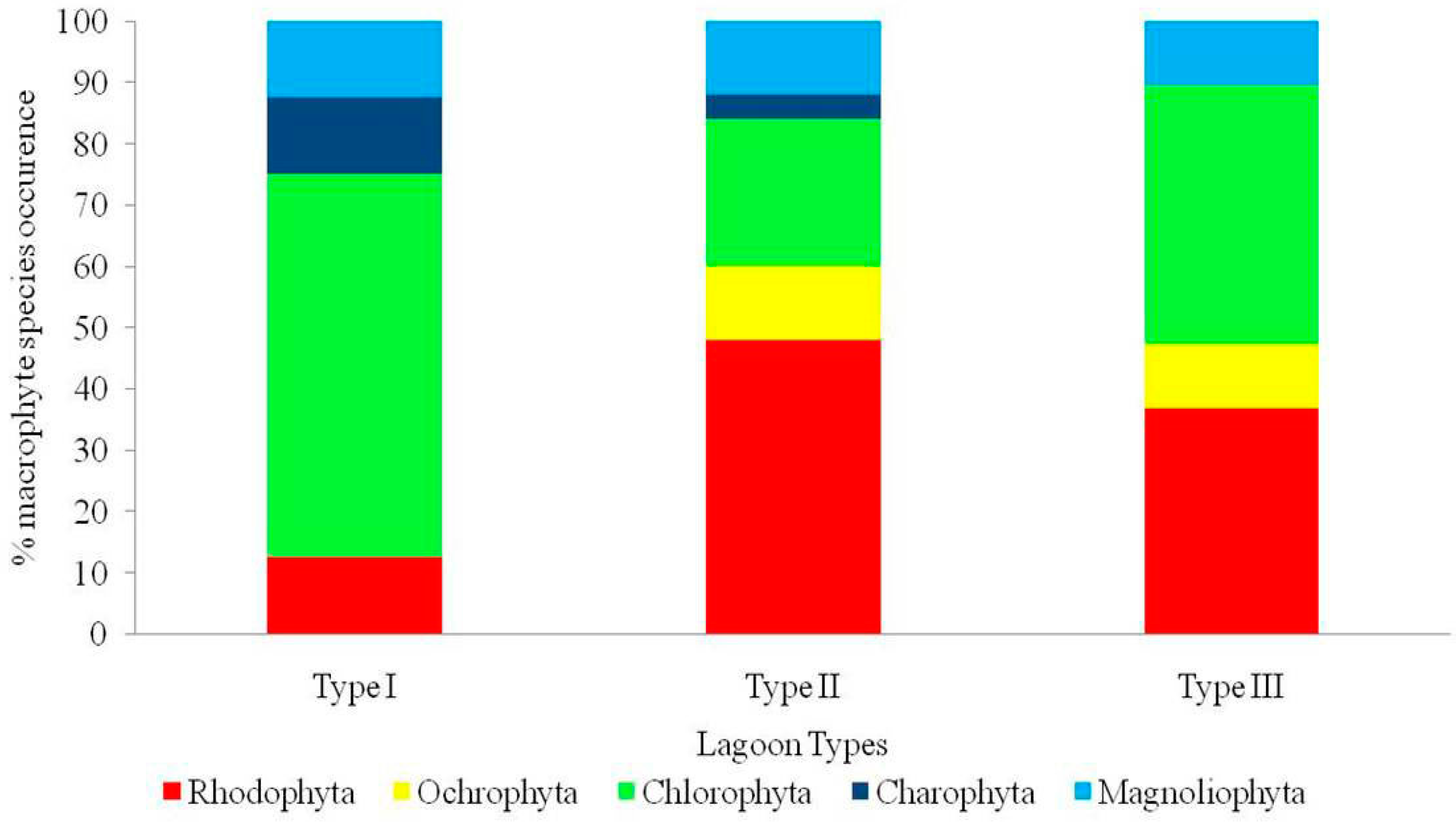
3.2. Macrophyte ‘Assemblages’ in the three Lagoon Types of Western Greece
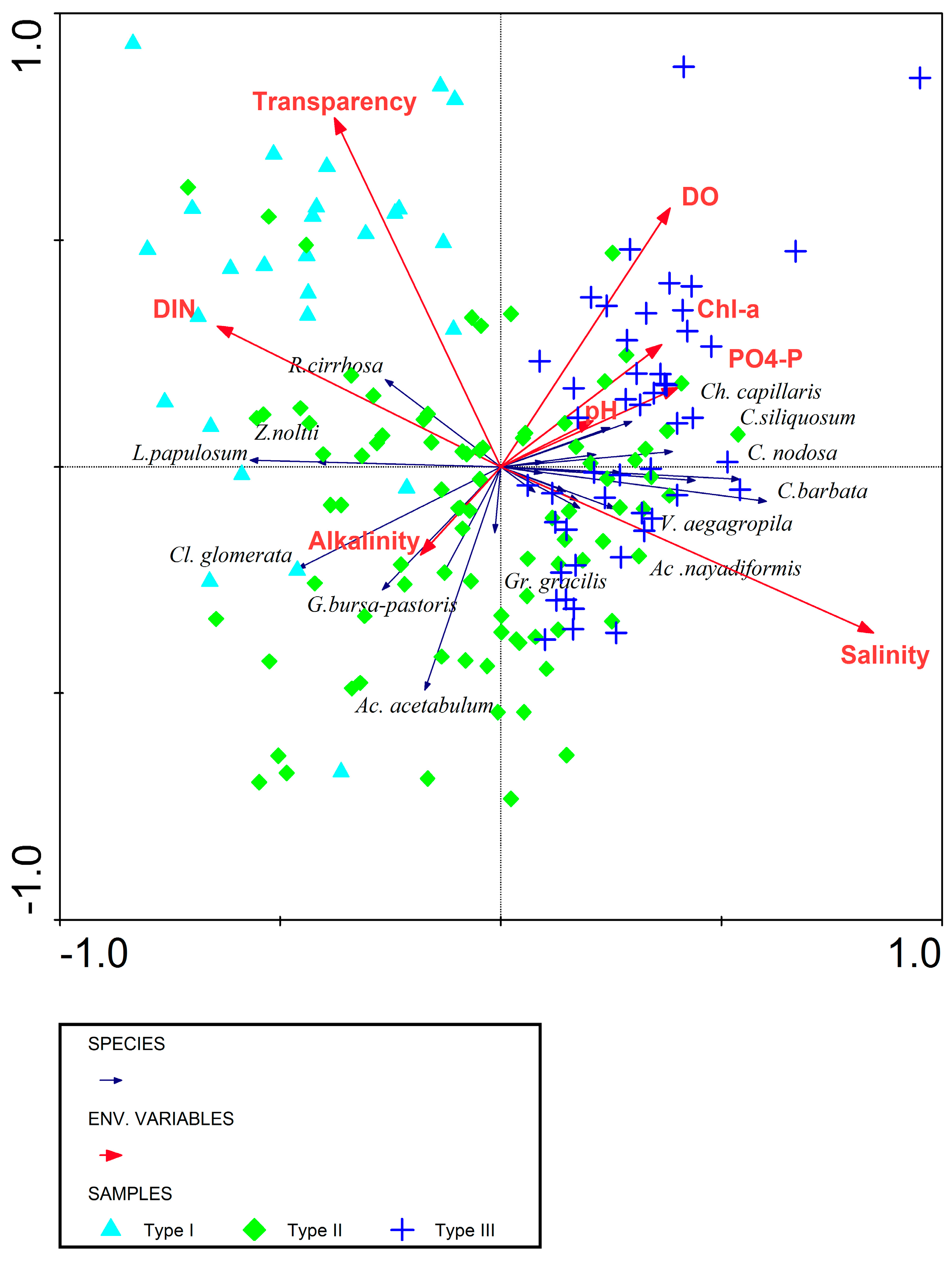
3.2.1. Relationship between Macrophytes and Environment
3.2.2. Comparisons of Macrophyte Assemblages among Lagoon Types
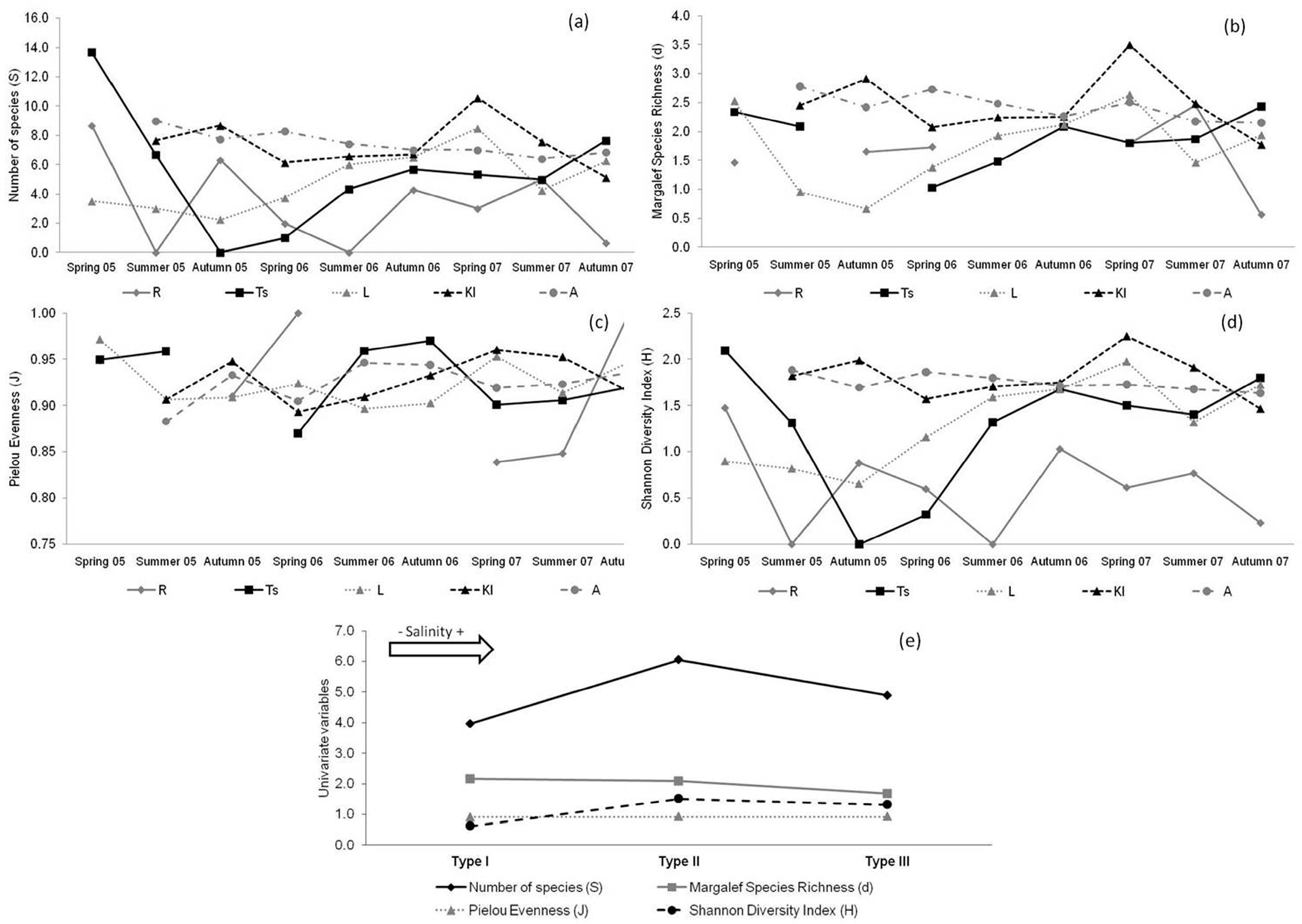
3.3. Univariate Variables of Diversity Indices and Environmental Variables
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kjerfve, B. Coastal Lagoons Processes; Elsevier Oceanography Series 60; Elsevier Science Publishers: Amsterdam, The Netherlands, 1994. [Google Scholar]
- Christia, C.; Papastergiadou, E. Spatial and temporal variations of aquatic macrophytes and water quality in six coastal lagoons of Western Greece. Belg. J. Bot. 2007, 140, 39–50. [Google Scholar]
- Rodriguez-Climent, S.; Caiola, N.; Ibanez, C. Salinity as the main factor structuring small-bodied fish assemblages in hydrologically altered Mediterranean coastal lagoons. Sci. Mar. 2013, 77, 37–45. [Google Scholar]
- Pulina, S.; Brutemark, A.; Suikkanen, S.; Padedda, B.M.; Grubisic, L.M.; Satta, C.T.; Caddeo, T.; Farina, P.; Sechi, N.; Lugliè, A. Effects of warming on a Mediterranean phytoplankton community. Web Ecol. 2016, 16, 89–92. [Google Scholar] [CrossRef]
- Bird, E.C.F. Physical setting and geomorphology of coastal lagoons. In Coastal Lagoon Processes; Kjerfve, B., Ed.; Elsevier: Amsterdam, The Netherlands, 1994; Chapter 2; pp. 9–40. [Google Scholar]
- Directive, E.C.W.F. Water Framework Directive 2000/60/EC of the European Parliament and of the Council establishing a framework for the Community action in the field of water policy. Off. J. Eur. Communities 2000, 43, 1–72. [Google Scholar]
- Guelorget, O.; Perthuisot, P. Le Domaine Paralique: Expressions Geologiques, Biologiques et Economiques du Confinement; Travaux du Laboratoire de Geologie de l’Ecole Normale Superiere: Paris, France, 1983; Volume 16, p. 136. [Google Scholar]
- Basset, A.; Sabbeta, L.; Fonnesu, A.; Mouillot, D.; Do Chi, T.; Viaroli, P.; Giardani, G.; Reizopoulou, S.; Abbiati, M.; Carrada, G.C. Typology in Mediterranean transitional waters: New challenges and perspectives. Aquat. Conserv. Mar. Freshw. Ecosyst. 2006, 16, 441–455. [Google Scholar] [CrossRef]
- Christia, C.; Giordani, G.; Papastergiadou, E. Assessment of ecological quality of coastal lagoons with a combination of phytobenthic and water quality indices. Mar. Pol. Bul. 2014, 86, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Umgiesser, G.; Ferrarin, C.; Cucco, A.; De Pascalis, F.; Bellafiore, D.; Ghezzo, M.; Bajo, M. Comparative hydrodynamics of 10 Mediterranean lagoons by means of numerical modeling. J. Geophys. Res. 2014, 119, 2212–22226. [Google Scholar] [CrossRef]
- Perez-Ruzafa, A.; Garcıa-Charton, J.A.; Barcala, E.; Marcos, C. Changes in benthic fish assemblages as a consequence of coastal works in a coastal lagoon: The Mar Menor (Spain, Western Mediterranean). Mar. Poll. Bull. 2006, 53, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, B. Final resolution of the symposium on the classification of brackish waters. Archo Oceanogr. Limnol. 1959, 11, 243–248. [Google Scholar]
- Tagliapietra, D.; Volpi Ghirardini, A. Notes on coastal lagoon typology in the light of the EU Water Framework Directive: Italy as a case study. Aquat. Conserv. Mar. Freshw. Ecosyst. 2006, 16, 457–467. [Google Scholar] [CrossRef]
- De Wit, R. Challenges for applying vulnerability assessments in coastal lagoons. Transit. Wat. Bull. 2011, 5, 32–41. [Google Scholar]
- Agostini, S.; Marchand, B.; Pergent, G. Temporal and spatial changes of seagrass meadows in a Mediterranean coastal lagoon. Oceanol. Acta 2003, 25, 297–302. [Google Scholar] [CrossRef]
- Adams, W.J.; Kimerle, R.A.; Barnett, R.A., Jr. Sediment quality and aquatic life assessment. Environ. Sci. Technol. 1992, 26, 1865–1875. [Google Scholar] [CrossRef]
- Lotze, H.K.; Lenihan, H.S.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.G.; Kay, M.C.; Kidwell, S.M.; Kirby, M.X.; Peterson, C.H.; Jackson, J.B.C. Depletion, degradation and recovery potential of estuaries and coastal seas. Science 2006, 312, 1806–1809. [Google Scholar] [CrossRef] [PubMed]
- Chappuis, E.; Gacia, E.; Ballesteros, E. Changes in aquatic macrophyte flora over the last century in Catalan water bodies (NE Spain). Aquat. Bot. 2011, 95, 268–277. [Google Scholar] [CrossRef]
- Chapman, P.M. Management of coastal lagoons under climate change. Estuar. Coast. Shelf Sci. 2012, 110, 32–35. [Google Scholar] [CrossRef]
- Duarte, C.M.; Borja, A.; Carstensen, J.; Elliott, M.; Krause-Jensen, D.; Marbà, N. Paradigms in the Recovery of Estuarine and Coastal Ecosystems. Estuar. Coasts 2015, 38, 1202–1212. [Google Scholar] [CrossRef]
- Littler, M.M.; Littler, D.S. A relative dominance moel for biotic reefs. In Proceedings of the Joint Meeting of the Atlantic Reef Committee Society of Reef Studies, Miami, FL, USA, 26–28 October 1984. [Google Scholar]
- Fodge, J.D.; Thomas, G.L.; Pauley, G.B. Effects of canopy formation by floating and submergent aquatic macrophytes on the water quality of two shallow Pacific Northwest lakes. Aquat. Bot. 1990, 38, 231–248. [Google Scholar] [CrossRef]
- Moore, K.A. Influence of seagrasses on water quality in shallow regions of the lower Chesapeake Bay. J. Coast. Res. 2004, 45, 162–178. [Google Scholar] [CrossRef]
- Malea, P.; Kevrekidis, Th.; Mogias, A. Annual versus perennial growth cycle in Ruppia maritima L: Temporal variation in population characteristics in Mediterranean lagoons (Monolimni and Drana Lagoons, Northern Aegean Sea). Bot. Mar. 2004, 47, 357–366. [Google Scholar] [CrossRef]
- Verhoeven, J.T.A. The ecology of Ruppia-dominated communities in Western Europe. III. Aspects of production, consumption and decomposition. Aquat. Bot. 1980, 8, 209–253. [Google Scholar] [CrossRef]
- Comín, F.A.; Menéndez, M.; Herrera, J.A. Spatial and temporal scales for monitoring coastal aquatic ecosystems. Aquat. Conserv. Mar. Freshw. Ecosyst. 2004, 14 (Suppl. S1), 5–17. [Google Scholar] [CrossRef]
- Viaroli, P.; Bartoli, M.; Azzoni, R.; Giordani, G.; Mucchino, C.; Naldi, M.; Nizzoli, D.; Tajé, L. Nutrient and iron limitation to Ulva blooms in a eutrophic coastal lagoon (Sacca di Goro, Italy). Hydrobiologia 2005, 550, 57–71. [Google Scholar] [CrossRef]
- Haritonidis, S.; Malea, P. Bioaccumulation of metals by the green alga Ulva rigida from Thermaikos Gulf, Greece. Environ. Pollut. 1999, 104, 365–372. [Google Scholar] [CrossRef]
- Orfanidis, S.; Panayotidis, P.; Stamatis, N. An insight to the Ecological Evaluation Index (EEI). Ecol. Ind. 2003, 3, 27–33. [Google Scholar] [CrossRef]
- Simboura, N.; Zenetos, A. Benthic indicators to use in ecological quality clas-sification of Mediterranean soft bottom marine ecosystems, including a new biotic index. Med. Mar. Sci. 2002, 3, 77–111. [Google Scholar] [CrossRef]
- Nicοlaidou, A.; Reizopoulou, S.; Koutsoubas, D.; Orfanidis, S.; Kevrekidis, T. Biological components of Greek lagoonal ecosystems: An overview. Mediterr. Mar. Sci. 2005, 6, 31–50. [Google Scholar] [CrossRef]
- Katselis, G.; Koukou, K.; Dimitriou, E.; Koutsikopoulos, C. Short- term seaward fish migration in the Messolonghi-Etoliko lagoons (Western Greek coast) in relation to climatic variables and the lunar cycle. Estuar. Coast. Shelf Sci. 2007, 73, 571–582. [Google Scholar] [CrossRef]
- Vasileiadou, K.; Pavloudi, C.; Kalantzi, I.; Apostolaki, E.T.; Chatzigeorgiou, G.; Chatzinikolaou, E.; Pafilis, E.; Papageorgiou, N.; Fanini, L.; Konstas, S.; et al. Environmental variability and heavy metal concentrations from five lagoons in the Ionian Sea (Amvrakikos Gulf, W. Greece). Biodiv. Data J. 2016, 4, e8233. [Google Scholar] [CrossRef] [PubMed]
- Orfanidis, S.; Panayotidis, P.; Stamatis, N. Ecological evaluation of transitional and coastal waters: A marine benthic macrophytes-based model. Mediterr. Mar. Sci. 2001, 2, 45–65. [Google Scholar] [CrossRef]
- Christia, C.; Tziortzis, I.; Fyttis, G.; Kashta, L.; Papastergiadou, E. A survey of the benthic aquatic flora of transitional water systems of Greece and Cyprus (Mediterranean Sea). Bot. Mar. 2011, 54, 169–178. [Google Scholar] [CrossRef]
- Sfriso, A.; Facca, C.; Ghetti, P.F. Validation of the Macrophyte Quality Index (MaQI) set up to assess the ecological status of Italian marine Transitional environments. Hydrobiologia 2009, 617, 117–141. [Google Scholar] [CrossRef]
- Le Fur, I.; De Wit, R.; Plus, M.; Oheix, J.; Simier, M.; Ouisse, V. Submerged benthic macrophytes in Mediterranean lagoons: Distribution patterns in relation to water chemistry and depth. Hydrobiologia 2017. [Google Scholar] [CrossRef]
- Sondergaard, M.; Johansson, L.S.; Lauridsen, T.L.; Jorgensen, T.B.; Liboriussen, L.; Jeppensen, E. Submerged macrophytes as indicators of the ecological quality of lakes. Freshw. Biol. 2010, 55, 893–908. [Google Scholar] [CrossRef]
- Viaroli, P.; Bartoli, M.; Giordani, G.; Naldi, M.; Orfanidis, S.; Zaldivar, J.M. Community shifts, alternative stable states, biogeochemical controls and feedbacks in eutrophic coastal lagoons: A brief overview. Aquat. Conserv. Mar. Freshw. Ecosyst. 2008, 18, 105–117. [Google Scholar] [CrossRef]
- Christia, C.; Papastergiadou, E.; Papatheodorou, G.; Geraga, M.; Papadakis, E. Seasonal and spatial variations of water quality, substrate and aquatic macrophytes based on side scan sonar, in an eastern Mediterranean lagoon (Kaiafas, Ionian Sea). Environ. Earth Sci. 2014, 71, 3543–3558. [Google Scholar] [CrossRef]
- AHPA. Standard Methods for the Examination of Water and Waste Water, 18th ed.; American Public Health Association: New York, NY, USA, 1989. [Google Scholar]
- Selig, U.; Schubert, M.; Eggert, A.; Steinhardt, T.; Sagert, S.; Schubert, H. The influence of sediments on soft bottom vegetation in inner coastal waters of Mecklenburg-Vorpommern (Germany). Estuar. Coast. Shelf Sci. 2007, 71, 241–249. [Google Scholar] [CrossRef]
- SPSS, Inc. SPSS v. 15.0 for Windows; SPSS, Inc.: Chicago, IL, USA, 2006. [Google Scholar]
- Ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Clarke, K.R.; Somerfield, P.J.; Chapman, M.G. On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray–Curtis coefficient for denuded assemblages. J. Exp. Mar. Biol. Ecol. 2006, 330, 55–80. [Google Scholar] [CrossRef]
- Ferrarin, C.; Umgiesser, G. Hydrodynamic modelling of a coastal lagoon: The Cabras lagoon in Sardinia, Italy. Ecol. Mod. 2005, 188, 340–357. [Google Scholar] [CrossRef]
- Rosqvist, K.; Mattila, J.; Sandström, A.; Snickars, M.; Westerbom, M. Regime shifts in vegetation composition of Baltic Sea coastal lagoons. Aquat. Bot. 2010, 93, 39–46. [Google Scholar] [CrossRef]
- Barker, P.; Leng, M.J.; Gasse, F.; Huang, Y. Century-tomillennial scale climatic variability in Lake Malawi reveales by isotope records. Earth Planet. Sc. Lett. 2007, 261, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Blomqvist, S.; Gunnars, A.; Elmgren, R. Why the limiting nutrient differs between temperate coastal seas and freshwater lakes: A matter of salt. Limnol. Oceanogr. 2004, 49, 2236–2241. [Google Scholar] [CrossRef]
- Carvalho, S.; Pereira, P.; Pereira, F.; de Pablo, H.; Vale, C.; Gaspar, M.B. Factors structuring temporal and spatial dynamics of macrobenthic communities in a eutrophic coastal lagoon (Obidos lagoon, Porugal). Mar. Environ. Res. 2011, 71, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Acquavita, A.; Aleffi, I.F.; Benci, C.; Bettoso, N.; Crevatin, E.; Milani, L.; Tamberlich, F.; Toniatti, L.; Barbieri, P.; Licen, S.; et al. Annual characterization of the nutrients and trophic state in a Mediterranean coastal lagoon: The Marano and Grado Lagoon (northern Adriatic Sea). Reg. Stud. Mar. Sci. 2015, 2, 132–144. [Google Scholar] [CrossRef]
- Van den Berg, M.S.; Coops, H.; Meijer, M.; Simons, J. Clear water associated with a dense Chara vegetation in the shallow and turbid lake Veluwenmeer. In The Structuring Role of Submerged Macrophytes in Lakes; Jeppesen, E., Søndergaard, M., Søndergaard, M., Christoffersen, K., Eds.; Springer: New York, NY, USA, 1998. [Google Scholar]
- Blindow, I.; Hargeby, A.; Andersson, G. Seasonal changes of mechanisms maintaining clear water in a shallow lake with abundant Chara vegetation. Aquat. Bot. 2002, 72, 315–334. [Google Scholar] [CrossRef]
- Del Pozo, R.; Fernandez-Alaez, C.; Fernadez-Alaez, M. The relative importance of natural and anthropogenic effects on community composition of aquatic macrophytes in Mediterranean ponds. Mar. Freshw. Res. 2011, 62, 101–109. [Google Scholar]
- Guidelines for Impact Assessment on Seagrass Meadows. Available online: http://www.rac-spa.org/sites/default/files/doc_vegetation/ld_etude_impact_en.pdf (accessed on 3 February 2018).
- Bamber, R.N.; Batten, S.D.; Sheader, M.; Bridgwater, N.D. On the ecology of brackish water lagoons in Great Britain. Aquat. Conserv. 1992, 2, 65–94. [Google Scholar] [CrossRef]
- Cognetti, G. Colonization of stressed coastal environment. Mar. Pol. Bul. 1992, 24, 12–14. [Google Scholar] [CrossRef]
- Green, E.P.; Short, F.T. World Atlas of Seagrasses; UNEP, WCMC: Berkeley, CA, USA, 2003. [Google Scholar]
- Reizopoulou, S.; Nicolaidou, A. Benthic diversity of coastal brackish-water lagoons in western Greece. Aquat. Conserv. 2004, 14, 93–102. [Google Scholar] [CrossRef]
- Rodriguez-Gallego, L.; Meerhoff, E.; Clemente, J.M.; Conde, D. Can ephemeral proliferations of submerged macrophytes influence zoobenthos and water quality in coastal lagoons? Hydrobiologia 2010, 646, 253–269. [Google Scholar] [CrossRef]
- Perez-Ruzafa, A.; Marcos, C.; Perez-Ruzafa, I.M.; Perez-Marcos, M. Coastal lagoons: “transitional ecosystems” between transitional and coastal waters. J. Coast Conserv. 2011, 15, 369–392. [Google Scholar] [CrossRef]
- Millet, B.; Guelorget, O. Spatial and seasonal variability in the relationships between benthic communities and physical environment in a lagoon ecosystem. Mar. Ecol. Prog. Ser. 1994, 198, 161–174. [Google Scholar] [CrossRef]
- Salas, F.; Neto, J.M.; Borja, A.; Marques, J.C. Evaluation of the applicability of a marine biotic index to characterize the status of estuarine ecosystems: The case of Mondego estuary (Portugal). Ecol. Ind. 2004, 4, 215–225. [Google Scholar] [CrossRef]
- Azzoni, R.; Giordani, G.; Bartoli, M.; Welsh, D.T.; Viaroli, P. Iron, sulphur and phosphorus cycling in the rhizosphere sediments of a eutrophic Ruppia cirrhosa meadow (Valle Smarlacca, Italy). J. Sea Res. 2001, 45, 15–26. [Google Scholar] [CrossRef]
- Zaldivar, J.M.; Cardoso, A.C.; Viaroli, P.; Newton, A.; De Wit, R.; Ibanez, C.; Reizopoulou, S.; Somma, F.; Razinkovas, A.; Basset, A.; et al. Eutrophication in transitional waters: An overview. Transit. Waters Monogr. 2008, 1, 1–78. [Google Scholar]
- Menéndez, M. Net production of Ruppia cirrhosa in Ebro Delta. Aquat. Bot. 2002, 73, 107–113. [Google Scholar] [CrossRef]
- Mannino, A.M.; Sara, G. The effect of Ruppia cirrhosa features on macroalgae and suspended matter in a Mediterranean shallow system. Mar. Ecol. 2006, 27, 350–360. [Google Scholar] [CrossRef]
- Warwick, R.M.; Clarke, K.R. Relationship between body-size, species abundance and diversity in marine benthic assemblages: Facts or artefacts? J. Exper. Mar. Biol. Ecol. 1996, 202, 63–71. [Google Scholar] [CrossRef]
- Charpentier, A.; Grillas, P.; Lescuyer, F.; Coulet, E.; Auby, I. Spatio-temporal dynamics of a Zostera noltii dominated community over a period of fluctuating salinity in a shallow lagoon, Southern France. Estuar. Coast. Shelf Sci. 2005, 64, 307–315. [Google Scholar] [CrossRef]
- Signorini, A.; Massini, G.; Migliore, G.; Tosoni, M.; Varrone, C.; Izzo, G. Sediment biogeochemical differences in two pristine Mediterranean coastal lagoons (in Italy) characterized by different phanerogam dominance—A comparative approach. Aquat. Conserv. Mar. Freshw. Ecosyst. 2008, 18, 27–44. [Google Scholar] [CrossRef]
- Pergent-Martini, C.; Pasqualini, V.; Ferrat, L.; Pergent, G.; Fernandez, C. Seasonal dynamics of Zostera noltii Hornem. In two Mediterranean lagoons. Hydrobiologia 2005, 543, 233–243. [Google Scholar] [CrossRef]
- Den Hartog, C. The Sea-Grasses of The World; North-Holland Publishing Company: Amsterdam, The Netherlands, 1970. [Google Scholar]
- Vermaat, J.E.; Verhagen, F.C.A.; Lindenburg, D. Contrasting responses in two populations of Zostera noltii Hornem. To experimental photoperiod manipulation at two salinities. Aquat. Bot. 2000, 67, 179–189. [Google Scholar] [CrossRef]
- Torquemada, Y.F.; Lizaso, J.L.S. Responses of two Mediterranean seagrasses to experimental changes in salinity. Hydrobiologia 2011, 669, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Prado, P.; Caiola, N.; Ibáñez, C. Spatio-temporal patterns of submerged macrophytes in three hydrologically altered mediterranean coastal lagoons. Estuar. Coasts 2013, 36, 414–429. [Google Scholar] [CrossRef]
- Marba, N.; Holmer, M.; Gacia, E.; Barron, C. Seagrass beds and coastal biogeochemistry. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R.J., Duarte, C.M., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 135–157. [Google Scholar]
- Fernández-Torquemada, Y.; Sánchez-Lizaso, J.L. Responses of two Mediterranean seagrasses to experimental changes in salinity. Hydrobiologia 2011, 669, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Menéndez, M.; Peñuelas, J. Sesonal photosynthetic and respiratory responses of Ruppia cirrhosa (Petagna) Grande to changes in light and temperature. Arch. Hydrobiol. 1993, 129, 221–230. [Google Scholar]
- Montesanto, B.; Panayotidis, P. The Cystoseira spp. Communities from the Aegean Sea (NE Mediterranean). Med. Mar. Sci. 2001, 2, 57–67. [Google Scholar] [CrossRef]
- Falace, A.; Curiel, D.; Sfriso, A. Study of the macrophyte assemblages and application of phytobenthic indices to assess the Ecological Status of the Marano-Grado lagoon (Italy). Mar. Ecol. 2009, 30, 480–494. [Google Scholar] [CrossRef]
- Peres, J.M.; Picard, J. Nouveau manuel de bionomie benthique de la mer Méditerranée. Rec. Trav. St. Mar. Endoume 1964, 31, 5–137. [Google Scholar]
- Giaccone, G.; Bruni, A. Le Cistoseire e la Vegetazione Sommersa del Mediterraneo; Atti dell’ Instituto Veneto de Scienze: Lett. ed Arti, Venezia, Italy, 1973; pp. 59–103. [Google Scholar]
- Waycott, M.; Duarte, C.M.; Carruthers, T.J.B.; Orth, R.J.; Dennison, W.C.; Olyarnik, S.; Calladine, A.; Fourqurean, J.W.; Heck, K.L., Jr.; Hughes, A.R.; et al. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12377–12381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aral, M.M.; Chang, B. Spatial Variation of Sea Level Rise at Atlantic and Mediterranean Coastline of Europe. Water 2017, 9, 522. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Parameters | Type I (n = 27) | Type II (n = 111) | Type III (n = 50) | Interaction | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Season | Year | Season | Year | Season | Year | Season*Year | ||||||||
| F | Sig | F | Sig | F | Sig | F | Sig | F | Sig | F | Sig | F | Sig | |
| Depth (m) | 411.7 | 0.000 | 2.2 | ns | 0.1 | ns | 1.2 | ns | 0.2 | ns | 1.2 | ns | 0.5 | ns |
| Transparency (m) | 301.7 | 0.000 | 0.1 | ns | 0.2 | ns | 0.0 | ns | 0.1 | ns | 0.0 | ns | 0.2 | ns |
| Temperature (°C) | 32,021.2 | 0.000 | 38.1 | 0.000 | 652.8 | 0.000 | 33.6 | 0.000 | 476.8 | 0.000 | 39.2 | 0.000 | 27.1 | 0.000 |
| pH | 249,734.3 | 0.000 | 32.9 | 0.000 | 22.3 | 0.000 | 25.2 | 0.000 | 9.6 | 0.002 | 27.1 | 0.000 | 33.9 | 0.000 |
| DO (mg/L) | 2308.2 | 0.000 | 19.6 | 0.000 | 43.1 | 0.000 | 7.1 | 0.006 | 26.1 | 0.000 | 10.9 | 0.001 | 37.8 | 0.000 |
| Salinity (‰) | 818,318.8 | 0.000 | 12,391.2 | 0.000 | 21,547.4 | 0.000 | 6788.5 | 0.000 | 19,105.6 | 0.000 | 7108.8 | 0.000 | 1414.0 | 0.000 |
| PO4-P (μg/L) | 434.8 | 0.000 | 9.9 | 0.001 | 14.4 | 0.000 | 10.0 | 0.001 | 8.1 | 0.003 | 8.6 | 0.003 | 7.4 | 0.001 |
| TP (μg/L) | 78.5 | 0.000 | 0.6 | ns | 2.3 | ns | 0.7 | ns | 1.4 | ns | 0.5 | ns | 5.1 | 0.007 |
| NO2-N (μg/L) | 39.4 | 0.000 | 11.9 | 0.001 | 22.4 | 0.000 | 6.2 | 0.010 | 29.3 | 0.000 | 6.9 | 0.006 | 7.2 | 0.001 |
| NO3-N (μg/L) | 57.3 | 0.000 | 10.8 | 0.001 | 8.0 | 0.004 | 6.6 | 0.007 | 2.2 | ns | 6.6 | 0.007 | 8.0 | 0.001 |
| NH4-N (μg/L) | 192.7 | 0.000 | 17.9 | 0.000 | 11.5 | 0.001 | 5.2 | 0.017 | 21.0 | 0.000 | 5.6 | 0.013 | 10.4 | 0.000 |
| Alkalinity (mg/L) | 132.4 | 0.000 | 16.9 | 0.000 | 14.8 | 0.000 | 11.0 | 0.001 | 8.5 | 0.003 | 10.6 | 0.001 | 6.4 | 0.002 |
| Chl-a (μg/L) | 1547.0 | 0.000 | 20.3 | 0.000 | 8.2 | 0.003 | 17.8 | 0.000 | 2.0 | ns | 19.5 | 0.000 | 18.2 | 0.000 |
| Lagoon Type | Macrophyte Assemblages |
|---|---|
| Τype Ι | i. Zostera noltii-Lamprothamnium papulosum-Gracilaria bursa pastoris-Cladophora glomerata ii. Ruppia cirrhosa-Acetabularia acetabulum-Gracilariopsis longissima-Ulva rigida |
| Τype ΙΙ | ii. Ruppia cirrhosa-Acetabularia acetabulum-Gracilariopsis longissima-Ulva rigida iii. Cymodocea nodosa-Chondria capillaris-Ceramium siliquosum-Ulva species iv. Cystoseira barbata, Alsidium corrallinum, Acanthophora nayadiformis, Gracilaria gracilis and Valonia aegagropila |
| Τype ΙΙΙ | iii. Cymodocea nodosa-Chondria capillaris-Ceramium siliquosum-Ulva species |
| Average Dissimilarity (%) between Lagoon Types | 2005 | 2006 | 2007 | ||||||
| Av. Dis. (I–II)= 77.1 Av. Dis. (I–III) = 95.9 Av. Dis. (II–III) = 83.8 | Av. Dis. (I–II) = 77.7 Av. Dis. (I–III) = 95.2 Av. Dis. (II–III) = 81.6 | Av. Dis. (I-II) = 77.8 Av. Dis. (I-III) = 97.6 Av. Dis. (II-III) = 83.0 | |||||||
| Type I | Type II | Type III | Type I | Type II | Type III | Type I | Type II | Type III | |
| Species | Av. Abund. | Av. Abund. | Av. Abund. | Av. Abund. | Av. Abund. | Av. Abund. | Av. Abund. | Av. Abund. | Av. Abund. |
| Acanthophora nayadiformis | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 6.1 | 0 |
| Acetabularia acetabulum | 0 | 14.3 | 0 | 0 | 4.2 | 0.2 | 0 | 7.1 | 0.5 |
| Ceramium diaphanum | 0 | 0 | 0 | 0 | 1.7 | 3.4 | 0 | 0.9 | 2.3 |
| Chaetomorpha linum | 6.6 | 5.6 | 1.5 | 1.8 | 2.9 | 1.6 | 0.7 | 3.4 | 0.9 |
| Chondria capillaris | 0 | 0 | 6 | 0 | 2.1 | 5.6 | 0 | 2.8 | 6.1 |
| Cladophora glomerata | 10 | 6.5 | 0 | 0.4 | 2.1 | 0 | 2.7 | 1.7 | 0 |
| Cladophora prolifera | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2.9 | 4.1 |
| Cladophora sp. | 0 | 0 | 0 | 0 | 2.8 | 3.2 | 0 | 0 | 0 |
| Cymodocea nodosa | 0 | 15.9 | 44.4 | 0 | 17.2 | 27.0 | 0 | 18.6 | 34.9 |
| Cystoseira barbata | 0 | 0 | 0 | 0 | 2.3 | 0 | 0 | 0 | 0 |
| Gracilaria bursa-pastoris | 0 | 6.4 | 0 | 0 | 0 | 0 | 0 | 4.4 | 3.3 |
| Gracilariopsis longissima | 0 | 0 | 4.5 | 0.8 | 4.1 | 16.1 | 0 | 0 | 9.3 |
| Lamprothamnium papulosum | 25.9 | 20.2 | 0 | 18.9 | 6.8 | 0 | 4.2 | 8.4 | 6.6 |
| Ruppia cirrhosa | 0 | 6.4 | 1.4 | 36.3 | 18.9 | 1.2 | 40.8 | 25.4 | 19.4 |
| Ulva lactuca | 0 | 0 | 12.1 | 6 | 0.4 | 9.7 | 8.4 | 0.4 | 4.6 |
| Ulva laetevirens | 0 | 0 | 0 | 0 | 0.5 | 2.2 | 0 | 0 | 0 |
| Ulva rigida | 0 | 5.9 | 2 | 0 | 2.4 | 3.0 | 0 | 0 | 1.8 |
| Zostera noltii | 27.8 | 4.5 | 0 | 2.3 | 0 | 0 | 2.8 | 0 | 0 |
| Average Dissimilarity (%) between Lagoon Types | Spring | Summer | Autumn | ||||||
| Av. Dis. (I–II) = 83.6 Av. Dis. (I–III) = 96.44 Av. Dis. (II–III) = 87.6 | Av. Dis. (I–II) = 82.4 Av. Dis. (I–III) = 95.9 Av. Dis. (II–III) = 80.1 | Av. Dis. (I-II) = 81.2 Av. Dis. (I-III) = 96.9 Av. Dis. (II-III) = 83.2 | |||||||
| Type I | Type II | Type III | Type I | Type II | Type III | Type I | Type II | Type III | |
| Species | Av. Abund. | Av. Abund. | Av. Abund. | Av. Abund. | Av. Abund. | Av. Abund. | Av. Abund. | Av. Abund. | Av. Abund. |
| Acanthophora nayadiformis | 0 | 2.4 | 0 | 0 | 4.7 | 0 | 0 | 3.1 | 0 |
| Acetabularia acetabulum | 0 | 8.4 | 0 | 0 | 8.0 | 1.3 | 0 | 6.9 | 0 |
| Ceramium diaphanum | 0 | 0 | 0 | 0 | 1.5 | 4 | 0 | 1.3 | 2.4 |
| Chaetomorpha linum | 1.5 | 2.8 | 1.5 | 6.2 | 3.3 | 0.8 | 1.6 | 5.0 | 1.5 |
| Chaetomorpha sp. | 0 | 0 | 2.4 | 0 | 0 | 0 | 0 | 0 | 0 |
| Chondria capillaries | 0 | 1.4 | 3.6 | 0 | 2.9 | 5.1 | 0 | 1.8 | 8.4 |
| Cladophora glomerata | 2.6 | 1.7 | 0 | 5.1 | 3.6 | 0 | 5.9 | 3.6 | 0 |
| Cladophora prolifera | 0 | 1.4 | 3.3 | 0 | 1 | 2.3 | 0 | 1.4 | 1 |
| Cladophora sp. | 0 | 1.5 | 2.1 | 0 | 0.6 | 2.3 | 0 | 0 | 0 |
| Cymodocea nodosa | 0 | 13.2 | 24.2 | 0 | 20.9 | 35.5 | 0 | 17.9 | 37.6 |
| Cystoseira barbata | 0 | 2.1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Gracilaria bursa-pastoris | 0 | 3.1 | 0 | 0 | 3.3 | 0 | 0 | 4.1 | 0 |
| Gracilariopsis longissima | 0 | 0.5 | 18.9 | 0 | 2.7 | 8.9 | 1 | 1.7 | 10.4 |
| Lamprothamnium papulosum | 4.1 | 9.3 | 0 | 17 | 10.4 | 0 | 19.6 | 12.2 | 0 |
| Ruppia cirrhosa | 22.5 | 23.9 | 0.4 | 31.7 | 15.8 | 0.5 | 29.7 | 15.6 | 0.3 |
| Ulva lactuca | 3 | 0.4 | 9.6 | 9.2 | 0.7 | 5.2 | 5.7 | 0.4 | 10.2 |
| Ulva rigida | 0 | 2.9 | 1.9 | 0 | 3.7 | 2.3 | 0 | 2.3 | 2.4 |
| Zostera noltii | 16.3 | 0.46 | 0 | 12 | 1.7 | 0 | 6.4 | 1.1 | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Christia, C.; Giordani, G.; Papastergiadou, E. Environmental Variability and Macrophyte Assemblages in Coastal Lagoon Types of Western Greece (Mediterranean Sea). Water 2018, 10, 151. https://doi.org/10.3390/w10020151
Christia C, Giordani G, Papastergiadou E. Environmental Variability and Macrophyte Assemblages in Coastal Lagoon Types of Western Greece (Mediterranean Sea). Water. 2018; 10(2):151. https://doi.org/10.3390/w10020151
Chicago/Turabian StyleChristia, Chrysoula, Gianmarco Giordani, and Eva Papastergiadou. 2018. "Environmental Variability and Macrophyte Assemblages in Coastal Lagoon Types of Western Greece (Mediterranean Sea)" Water 10, no. 2: 151. https://doi.org/10.3390/w10020151
APA StyleChristia, C., Giordani, G., & Papastergiadou, E. (2018). Environmental Variability and Macrophyte Assemblages in Coastal Lagoon Types of Western Greece (Mediterranean Sea). Water, 10(2), 151. https://doi.org/10.3390/w10020151