Ammonium Transformation in 14 Lakes along a Trophic Gradient




Abstract
:
1. Introduction
2. Materials and Methods
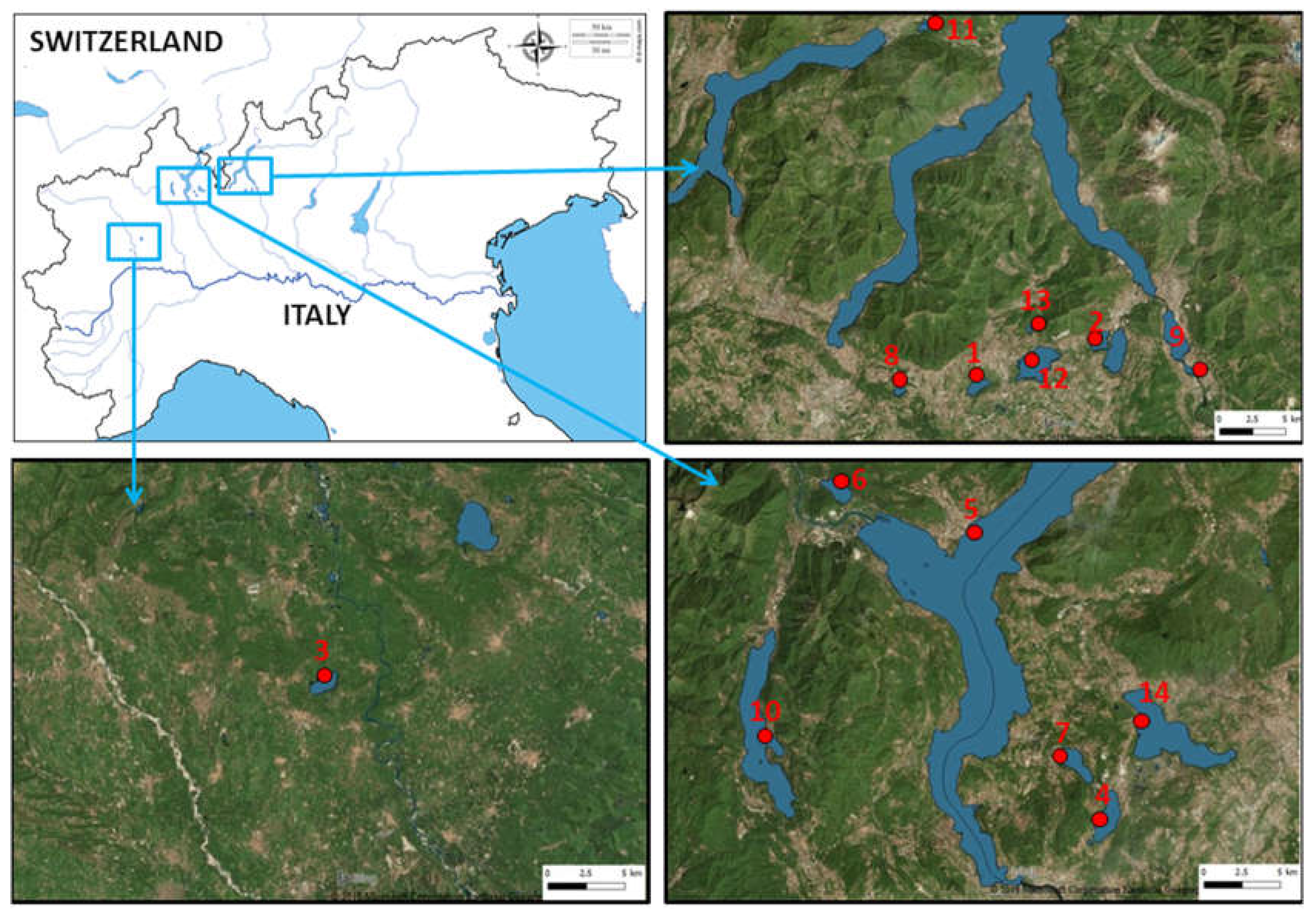
2.1. Study Sites
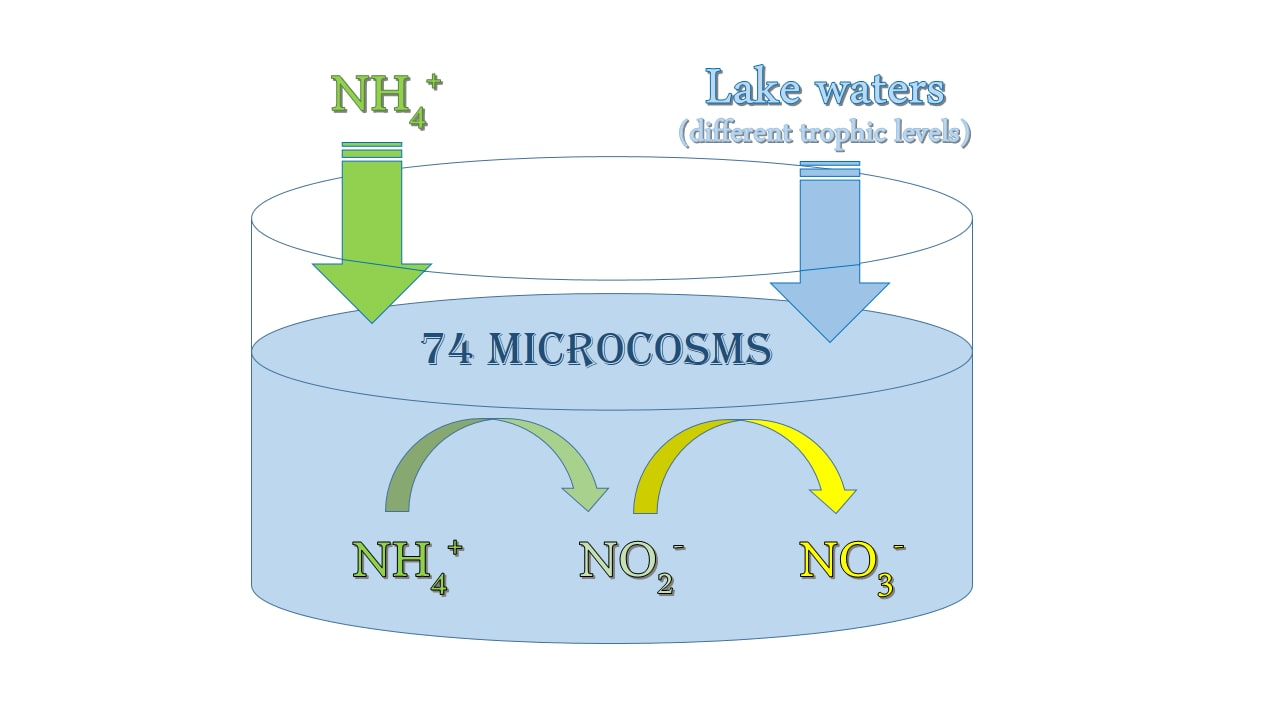
2.2. Sampling Method and Experimental Procedure
2.3. Data Analyses
3. Results
3.1. Lakes Characteristics
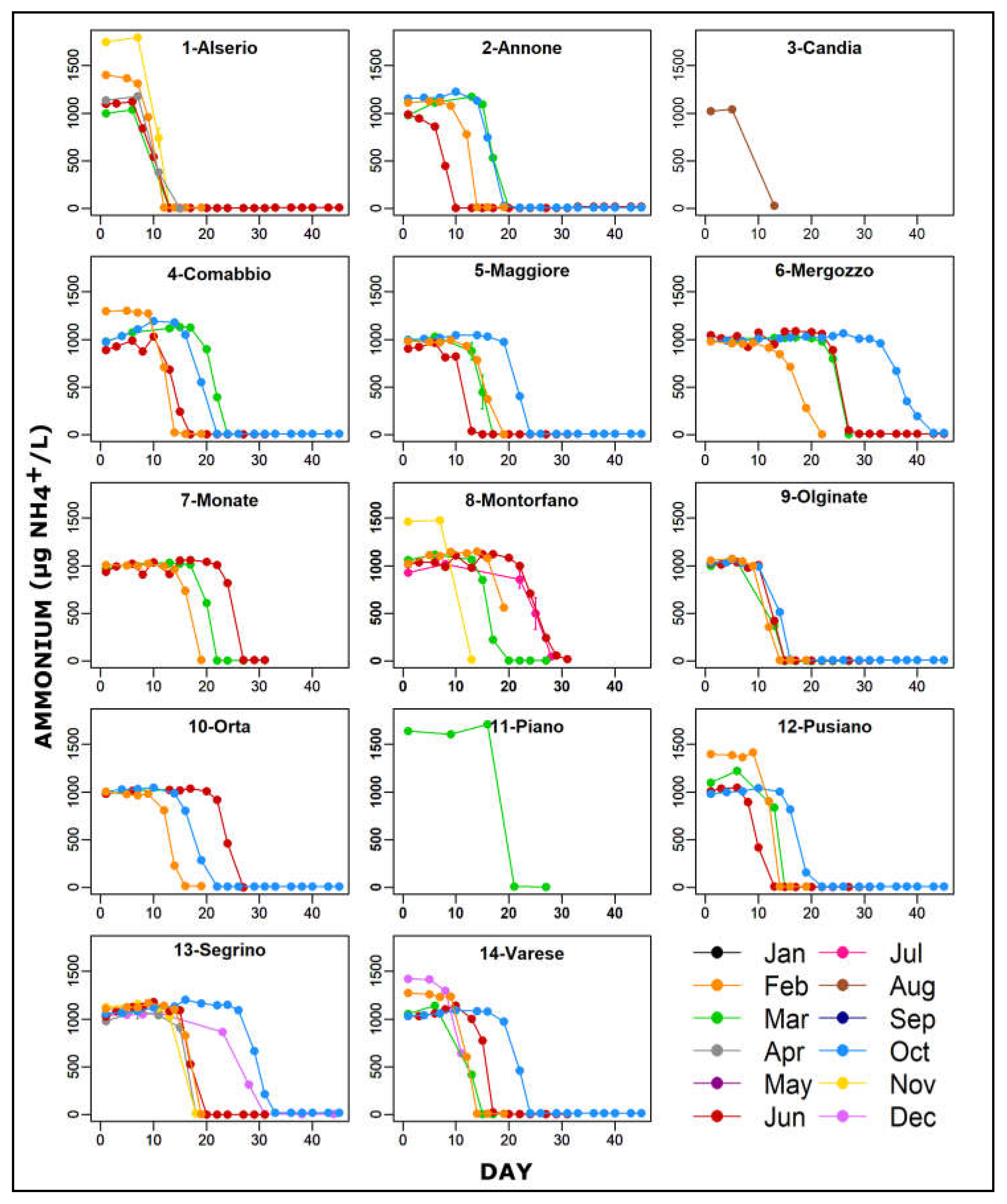
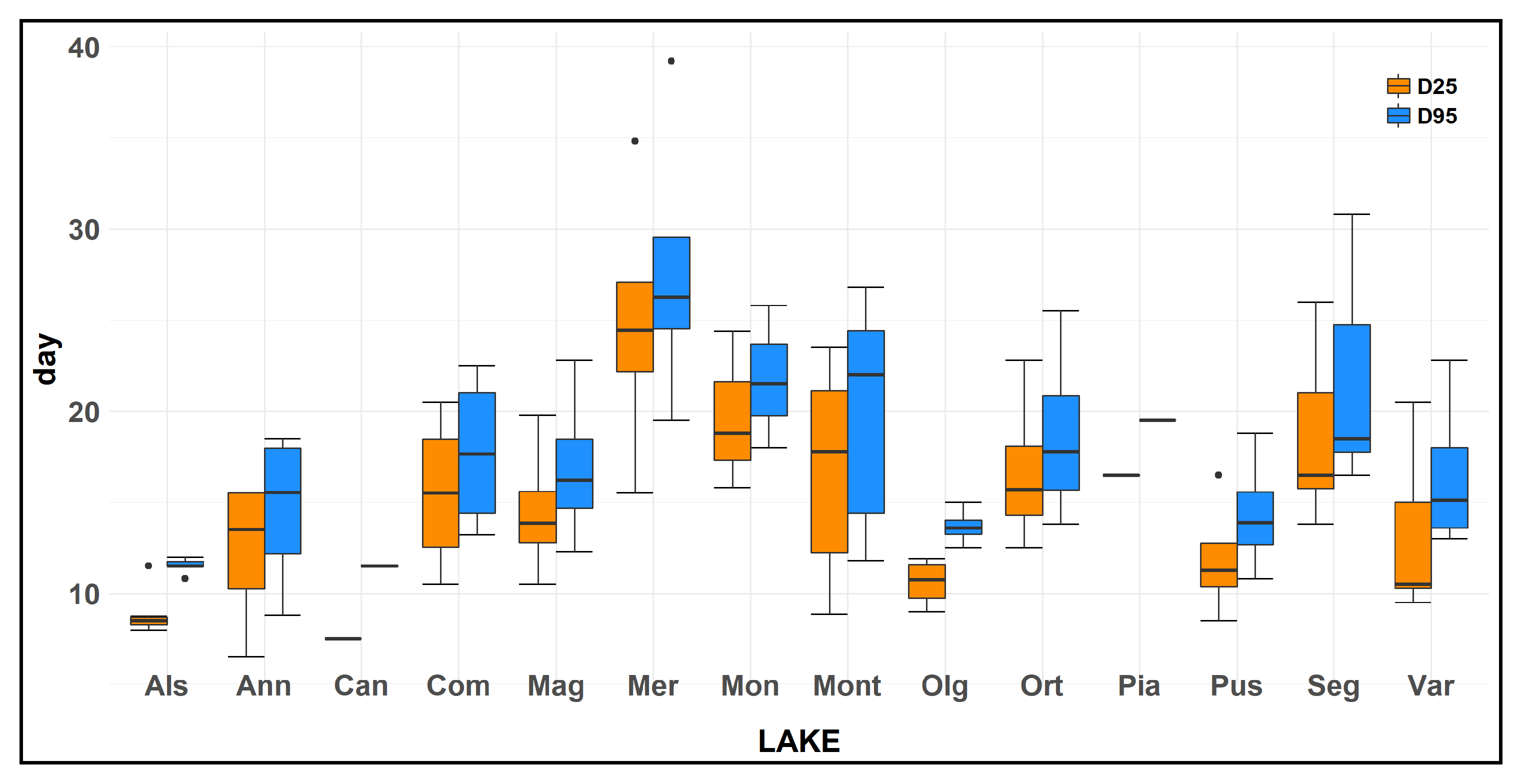
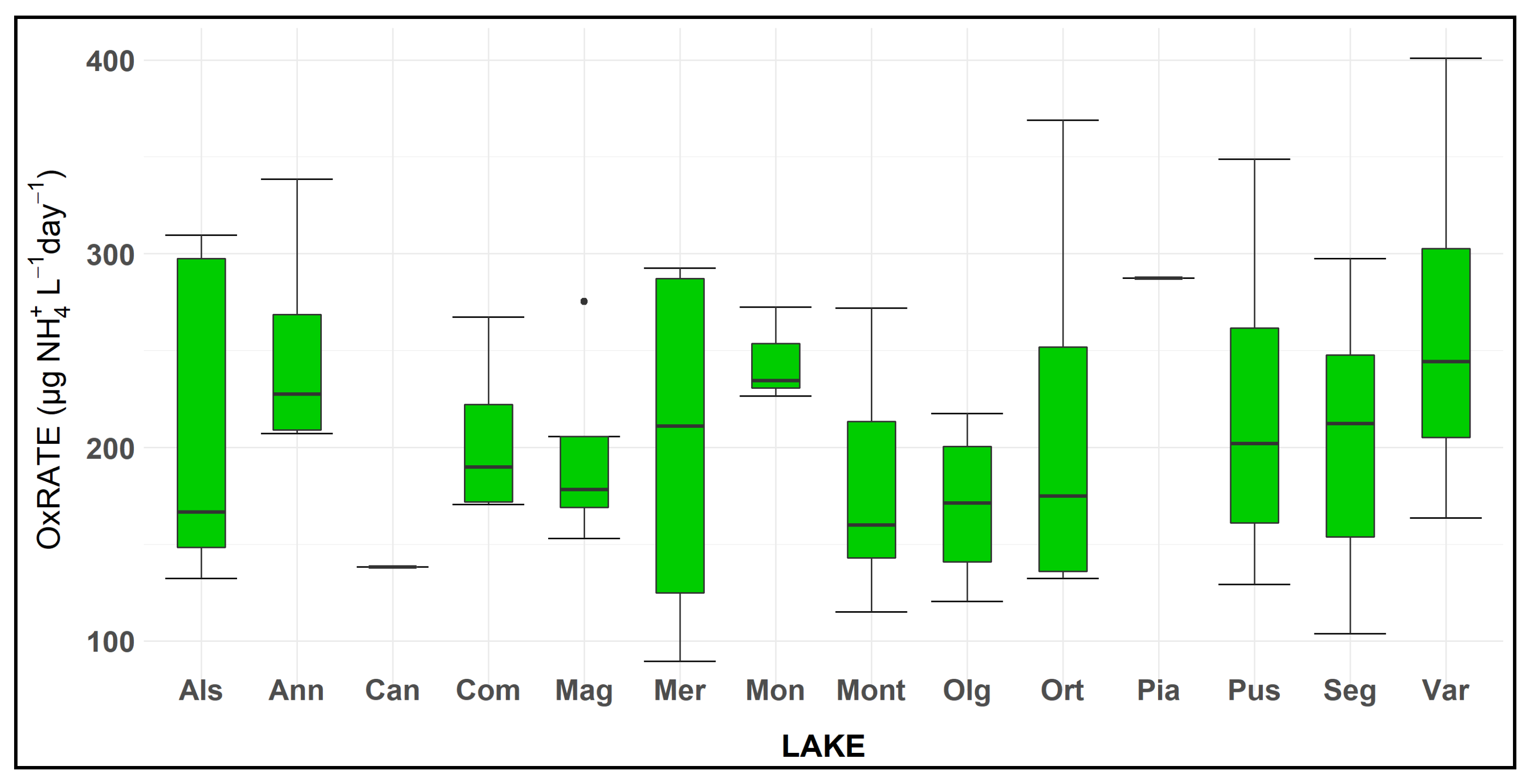
3.2. Ammonium Oxidation
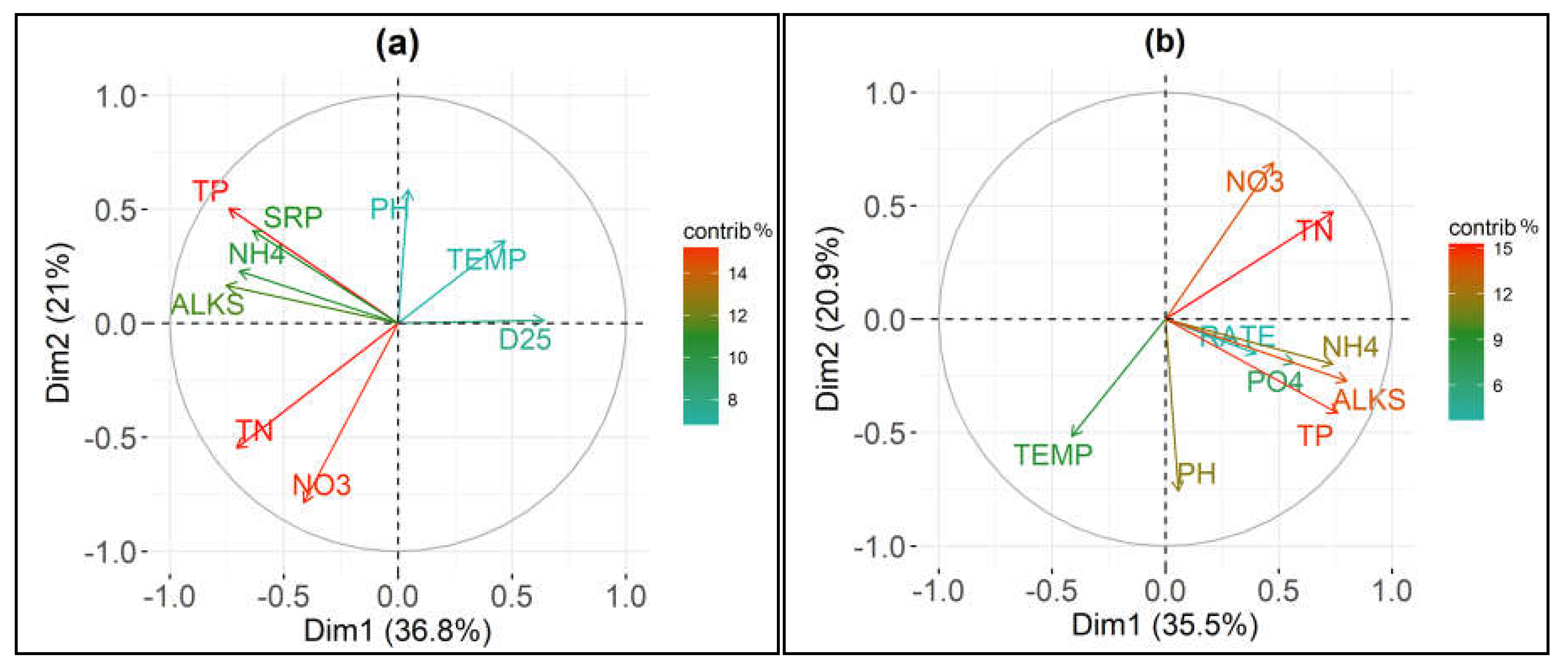
3.3. Relationship between Ammonium Oxidation and Chemico-Physical Parameters
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Benli, A.Ç.K.; Köksal, G.; Özkul, A. Sublethal ammonia exposure of Nile tilapia (Oreochromis niloticus L.): Effects on gill, liver and kidney histology. Chemosphere 2008, 72, 1355–1358. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yu, N.; Qin, J.G.; Li, E.; Du, Z.; Chen, L. Effects of ammonia stress, dietary linseed oil and Edwardsiella ictaluri challenge on juvenile darkbarbel catfish Pelteobagrus vachelli. Fish Shellfish Immunol. 2014, 38, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Collos, Y.; Harrison, P.J. Acclimation and toxicity of high ammonium concentrations to unicellular algae. Mar. Pollut. Bull. 2014, 80, 8–23. [Google Scholar] [CrossRef] [PubMed]
- Zekker, I.; Rikmann, E.; Tenno, T.; Kroon, K.; Vabamäe, P.; Salo, E.; Tenno, T.; Loorits, L.; Dc Rubin, S.S.C.; Vlaeminck, S.E. Deammonification process start-up after enrichment of anammox microorganisms from reject water in a moving-bed biofilm reactor. Environ. Technol. (United Kingdom) 2013, 34, 3095–3101. [Google Scholar] [CrossRef] [PubMed]
- Zekker, I.; Rikmann, E.; Tenno, T.; Vabamäe, P.; Kroon, K.; Loorits, L.; Saluste, A.; Tenno, T. Effect of concentration on anammox nitrogen removal rate in a moving bed biofilm reactor. Environ. Technol. 2012, 33, 2263–2271. [Google Scholar] [CrossRef] [PubMed]
- Leoni, B.; Garibaldi, L. Population dynamics of Chaoborus flavicans and Daphnia spp.: Effects on a zooplankton community in a volcanic eutrophic lake with naturally high metal concentrations (L. Monticchio Grande, Southern Italy). J. Limnol. 2009, 68, 37–45. [Google Scholar] [CrossRef]
- Wajsbrot, N.; Gasith, A.; Diamant, A.; Popper, D.M. Chronic toxicity of ammonia to juvenile gilthead seabream Sparus aurata and related histopathological effects. J. Fish Biol. 1993, 42, 321–328. [Google Scholar] [CrossRef]
- Hargreaves, J.A.; Tucker, C.S. Managing Ammonia in Fish Ponds; SRAC Publication: Stoneville, MS, USA, 2004. [Google Scholar]
- Brandes, J.A.; Devol, A.H.; Deutsch, C. New developments in the marine nitrogen cycle. Chem. Rev. 2007, 107, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Zekker, I.; Rikmann, E.; Kroon, K.; Mandel, A.; Mihkelson, J. Ameliorating nitrite inhibition in a low-temperature nitritation–anammox MBBR using bacterial intermediate nitric oxide. Int. J. Environ. Sci. Technol. 2017, 14, 2343–2356. [Google Scholar] [CrossRef]
- Hayden, C.J.; Beman, J.M. High abundances of potentially active ammonia-oxidizing bacteria and archaea in oligotrophic, high-altitude lakes of the Sierra Nevada, California, USA. PLoS ONE 2014, 9, e111560. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, M.; Ray, A.; Post, A.F.; McKay, R.M.; Bullerjahn, G.S. Identification, enumeration and diversity of nitrifying planktonic archaea and bacteria in trophic end members of the Laurentian Great Lakes. J. Great Lakes Res. 2016, 1, 39–49. [Google Scholar] [CrossRef]
- Bernhard, A.E.; Tucker, J.; Giblin, A.E.; Stahl, D.A. Functionally distinct communities of ammonia-oxidizing bacteria along an estuarine salinity gradient. Environ. Microbiol. 2007, 9, 1439–1447. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, J.; Wang, Z.; Pei, Y. Nitrification in lake sediment with addition of drinking water treatment residuals. Water Res. 2014, 56, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Randall, D.J.; Tsui, T.K.N. Ammonia toxicity in fish. Mar. Pollut. Bull. 2002, 45, 17–23. [Google Scholar] [CrossRef]
- Sinha, A.K.; Liew, H.J.; Diricx, M.; Blust, R.; De Boeck, G. The interactive effects of ammonia exposure, nutritional status and exercise on metabolic and physiological responses in gold fish (Carassius auratus L.). Aquat. Toxicol. 2012, 109, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.H.; Yang, F.F.; Ling, R.Z.; Liao, S.A.; Miao, Y.T.; Ye, C.X.; Wang, A.L. Effects of ammonia exposure on apoptosis, oxidative stress and immune response in pufferfish (Takifugu obscurus). Aquat. Toxicol. 2015, 164, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Rikmann, E.; Zekker, I.; Tenno, T.; Saluste, A.; Tenno, T. Inoculum-free start-up of biofilm- and sludge-based deammonification systems in pilot scale. Int. J. Environ. Sci. Technol. 2018, 15, 133–148. [Google Scholar] [CrossRef]
- Pauer, J.J.; Auer, M.T. Nitrification in the water column and sediment of a hypereutrophic lake and adjoining river system. Water Res. 2000, 34, 1247–1254. [Google Scholar] [CrossRef]
- Beaulieu, J.J.; Tank, J.L.; Hamilton, S.K.; Wollheim, W.M.; Hall, R.O.; Mulholland, P.J.; Peterson, B.J.; Ashkenas, L.R.; Cooper, L.W.; Dahm, C.N.; et al. Nitrous oxide emission from denitrification in stream and river networks. Proc. Natl. Acad. Sci. USA 2011, 108, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wei, Z.; Zhao, Y.; Xi, B.; Wang, X.; Zhao, T.; Zhang, X.; Wei, Y. Environmental factors influencing the distribution of ammonifying and denitrifying bacteria and water qualities in 10 lakes and reservoirs of the Northeast, China. Microb. Biotechnol. 2015, 8, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Smith, V.; Wood, S.; McBride, C.; Atalah, J.; Hamilton, D. Phosphorus and nitrogen loading restraints are essential for successful eutrophication control of Lake Rotorua, New Zealand. Inland Waters 2016, 6, 273–283. [Google Scholar] [CrossRef]
- Lipschultz, F.; Wofsy, S.C.; Fox, L.E. The effects of light and nutrients on rates of ammonium transformation in a eutrophic river. Mar. Chem. 1985, 16, 329–341. [Google Scholar] [CrossRef]
- Usui, T.; Koike, I.; Ogura, N. N2O production, nitrification and denitrification in an estuarine sediment. Estuar. Coast. Shelf Sci. 2001, 52, 769–781. [Google Scholar] [CrossRef]
- Strauss, E.A.; Richardson, W.B.; Bartsch, L.A.; Cavanaugh, J.C.; Bruesewitz, D.A.; Imker, H.; Heinz, J.A.; Soballe, D.M. Nitrification in the Upper Mississippi River: Patterns, controls, and contribution to the NO3− budget. J. N. Am. Benthol. Soc. 2004, 23, 1–14. [Google Scholar] [CrossRef]
- Small, G.E.; Bullerjahn, G.S.; Sterner, R.W.; Beall, B.F.N.; Brovold, S.; Finlay, J.C.; McKay, R.M.L.; Mukherjee, M. Rates and controls of nitrification in a large oligotrophic lake. Limnol. Oceanogr. 2013, 58, 276–286. [Google Scholar] [CrossRef]
- Leoni, B.; Nava, V.; Patelli, M. Relationship among climate variability, Cladocera phenology and pelagic food web in deep lakes in different trophic state. Mar. Freshw. Res. 2017. accepted for publication. [Google Scholar]
- Kowalczewska-Madura, K.; Godyn, R.; Dera, M. Spatial and seasonal changes of phosphorus internal loading in two lakes with different trophy. Ecol. Eng. 2015, 74, 187–195. [Google Scholar] [CrossRef]
- Poste, A.E.; Muir, D.C.G.; Guildford, S.J.; Hecky, R.E. Bioaccumulation and biomagnification of mercury in African lakes: The importance of trophic status. Sci. Total Environ. 2015, 506–507, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Premazzi, G.; Chiaudani, G. Ecological Quality of Surface Waters: Quality Assessment Schemes for European Community Lakes; European Commission: Brussels, Luxembourg, 1992; Volume 14563.
- Ambrosetti, W.; Barbanti, L. Physical limnology of Italian lakes. 1. Relationship between morphometry and heat content. J. Limnol. 2003, 61, 147–157. [Google Scholar] [CrossRef]
- Leoni, B.; Morabito, G.; Rogora, M.; Pollastro, D.; Mosello, R.; Arisci, S.; Forasacco, E.; Garibaldi, L. Response of planktonic communities to calcium hydroxide addition in a hardwater eutrophic lake: Results from a mesocosm experiment. Limnology 2007, 8, 121–130. [Google Scholar] [CrossRef]
- Riccardi, N.; Mangoni, M. Considerations on the biochemical composition of some freshwater zooplankton species. J. Limnol. 1999, 58, 58–65. [Google Scholar] [CrossRef]
- Volta, P.; Jeppesen, E.; Leoni, B.; Campi, B.; Sala, P.; Garibaldi, L.; Lauridsen, T.L.; Winfield, I.J. Recent invasion by a non-native cyprinid (common bream Abramis brama) is followed by major changes in the ecological quality of a shallow lake in southern Europe. Biol. Invasions 2013, 15, 2065–2079. [Google Scholar] [CrossRef] [Green Version]
- Rogora, M.; Mosello, R.; Kamburska, L.; Salmaso, N.; Cerasino, L.; Leoni, B.; Garibaldi, L.; Soler, V.; Lepori, F.; Colombo, L.; et al. Recent trends in chloride and sodium concentrations in the deep subalpine lakes (Northern Italy). Environ. Sci. Pollut. Res. 2015, 22, 19013–19026. [Google Scholar] [CrossRef] [PubMed]
- American Public Health Association (APHA). Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 1998; p. 552. [Google Scholar]
- Marti, C.M.; Imberger, J.; Garibaldi, L.; Leoni, B. Using time scales to characterize phytoplankton assemblages in a deep subalpine lake during the thermal stratification period: Lake Iseo, Italy. Water Resour. Res. 2015, 52, 1762–1780. [Google Scholar] [CrossRef]
- Van Der Maaten, L.J.P.; Postma, E.O.; Van Den Herik, H.J. Dimensionality Reduction: A Comparative Review. J. Mach. Learn. Res. 2009, 10, 1–41. [Google Scholar] [CrossRef]
- Jollife, I.T.; Cadima, J. Principal component analysis: A review and recent developments. Philos. Trans. A Math. Phys. Eng. Sci. 2016, 374, 20150202. [Google Scholar] [CrossRef] [PubMed]
- Madsen, H.; Thyregod, P. Introduction to General and Generalized Linear Models; CRC Press: Boca Raton, FL, USA, 2010; ISBN 9781420091557. [Google Scholar]
- Leoni, B.; Marti, C.L.; Imberger, J.; Garibaldi, L. Summer spatial variations in phytoplankton composition and biomass in surface waters of a warm-temperate, deep, oligo-holomictic lake: Lake Iseo, Italy. Inland Waters 2014, 4, 303–310. [Google Scholar] [CrossRef]
- Miller, A.J. Subset Selection in Regression; Chapman & Hall/CRC: Boca Raton, FL, USA, 2002; ISBN 9781584881711. [Google Scholar]
- Schützenmeister, A.; Jensen, U.; Piepho, H.P. Checking normality and homoscedasticity in the general linear model using diagnostic plots. Commun. Stat. Simul. Comput. 2012, 41, 141–154. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2017; ISBN 3_900051_00_3. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2009; Volume 35, ISBN 9780387981406. [Google Scholar]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix (Version 0.84). Available online: https://github.com/taiyun/corrplot (accessed on 14 November 2017).
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses (Version 1.0.5). Available online: https://CRAN.R-project.org/package=factoextra (accessed on 22 August 2017).
- Camargo, J.A.; Alonso, Á. Ecological and toxicological effects of inorganic nitrogen pollution in aquatic ecosystems: A global assessment. Environ. Int. 2006, 32, 831–849. [Google Scholar] [CrossRef] [PubMed]
- Rabalais, N.N. Nitrogen in Aquatic Ecosystems. Ambio 2002, 31, 102–112. [Google Scholar] [CrossRef] [PubMed]
- U.S. Environmental Protection Agency. Aquatic Life Ambient Water Quality Criteria for Ammonia—Freshwater; U.S. Environmental Protection Agency: Washington, DC, USA, 2013; Volume 78, pp. 67–79.
- Netten, J.J.C.; van der Heide, T.; Smolders, A.J.P. Interactive effects of pH, temperature and light during ammonia toxicity events in Elodea canadensis. Chem. Ecol. 2013, 29, 448–458. [Google Scholar] [CrossRef]
- Loken, L.C.; Small, G.E.; Finlay, J.C.; Sterner, R.W.; Stanley, E.H. Nitrogen cycling in a freshwater estuary. Biogeochemistry 2016, 127, 199–216. [Google Scholar] [CrossRef]
- Gobler, C.J.; Burkholder, J.M.; Davis, T.W.; Harke, M.J.; Johengen, T.; Stow, C.A.; Van De Waal, D.B. The dual role of nitrogen supply in controlling the growth and toxicity of cyanobacterial blooms. Harmful Algae 2016, 54, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Nava, V.; Patelli, M.; Soler, V.; Leoni, B. Interspecific relationship and ecological requirements of two potentially harmful cyanobacteria in a Deep South-Alpine Lake (L. Iseo, I). Water (Switzerland) 2017, 9, 993. [Google Scholar] [CrossRef]
- Glibert, P.M.; Wilkerson, F.P.; Dugdale, R.C.; Raven, J.A.; Dupont, C.L.; Leavitt, P.R.; Parker, A.E.; Burkholder, J.M.; Kana, T.M. Pluses and minuses of ammonium and nitrate uptake and assimilation by phytoplankton and implications for productivity and community composition, with emphasis on nitrogen-enriched conditions. Limnol. Oceanogr. 2016, 61, 165–197. [Google Scholar] [CrossRef]
- Nimptsch, J.; Pflugmacher, S. Ammonia triggers the promotion of oxidative stress in the aquatic macrophyte Myriophyllum mattogrossense. Chemosphere 2007, 66, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Gerletti, M.; Provini, A. Effect of nitrification in Lake Orta. In Ninth International Conference on Water Pollution Research, Proceedings of the 9th International Conference, Stockholm, Sweden, 12–16 June 1978; Elsevier: Amsterdam, The Netherlands, 1979; pp. 839–851. ISBN 9780080229393. [Google Scholar]
- Kim, J.H.; Guo, X.; Park, H.S. Comparison study of the effects of temperature and free ammonia concentration on nitrification and nitrite accumulation. Process Biochem. 2008, 43, 154–160. [Google Scholar] [CrossRef]
- Anthonisen, A.; Loehr, R.; Prakasam, T.; Srinath, E. Inhibition of Nitrification by Ammonia and Nitrous Acid. J. Water Pollut. Control Fed. 1976, 48, 835–852. [Google Scholar] [CrossRef] [PubMed]
- Hira, D.; Aiko, N.; Yabuki, Y.; Fujii, T. Impact of aerobic acclimation on the nitrification performance and microbial community of landfill leachate sludge. J. Environ. Manag. 2018, 209, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, J.; Zhao, Q.; Zhou, Q.; Li, N.; Wang, Y.; Xie, S.; Liu, Y. Sediment Ammonia-Oxidizing Microorganisms in Two Plateau Freshwater Lakes at Different Trophic States. Microb. Ecol. 2016, 71, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, G.; Jeison, D.; Chamy, R. Nitrification with high nitrite accumulation for the treatment of wastewater with high ammonia concentration. Water Res. 2003, 37, 1371–1377. [Google Scholar] [CrossRef]
- Biesterfeld, S.; Farmer, G.; Russell, P.; Figueroa, L. Effect of alkalinity type and concentration on nitrifying biofilm activity. Water Environ. Res. 2003, 75, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Shanahan, J.W.; Semmens, M.J. Alkalinity and pH effects on nitrification in a membrane aerated bioreactor: An experimental and model analysis. Water Res. 2015, 74, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Irvin, S. The comparison of alkalinity and ORP as indicators for nitrification and denitrification in a sequencing batch reactor (SBR). Biochem. Eng. J. 2007, 34, 248–255. [Google Scholar] [CrossRef]
- Lawson, T.B. Fundamentals of Aquacultural Engineering; Springer: Boston, MA, USA, 1995; ISBN 978-1-4612-7578-7. [Google Scholar]
- Isnansetyo, A.; Getsu, S.; Seguchi, M.; Koriyama, M. Independent Effects of Temperature, Salinity, Ammonium Concentration and pH on Nitrification Rate of the Ariake Seawater Above Mud Sediment. HAYATI J. Biosci. 2014, 21, 21–30. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lake | Area (km2) | Volume (m3 × 106) | Depthmax (m) | Depthmean (m) | Trophic Status |
|---|---|---|---|---|---|
| 1-Alserio | 1.23 | 6.55 | 8.1 | 5.3 | Hypereutrophic |
| 2-Annone (W) | 1.75 | 7.60 | 10.0 | 4.5 | Eutrophic |
| 3-Candia | 1.49 | 8.10 | 7.7 | 3.8 | Meso-eutrophic |
| 4-Comabbio | 3.58 | 16.40 | 7.7 | 4.6 | Hypereutrophic |
| 5-Maggiore | 213 | 37500 | 370 | 176 | Oligotrophic |
| 6-Mergozzo | 1.83 | 83.00 | 73.0 | 45.4 | Oligotrophic |
| 7-Monate | 2.51 | 45.00 | 34.0 | 14.4 | Oligotrophic |
| 8-Montorfano | 1.90 | 1.90 | 6.8 | 4.0 | Meso-eutrophic |
| 9-Olginate | 0.58 | 7.00 | 17.0 | 8.0 | Meso-eutrophic |
| 10-Orta | 18.10 | 1286 | 143 | 70.9 | Oligotrophic |
| 11-Piano | 0.63 | 4.03 | 12.5 | 5.1 | Eutrophic |
| 12-Pusiano | 4.93 | 69.00 | 24.3 | 14.0 | Eutrophic |
| 13-Segrino | 0.38 | 1.20 | 9.0 | 4.6 | Oligotrophic |
| 14-Varese | 14.90 | 162 | 26.0 | 9.9 | Hypereutrophic |
| Lake | n | TP (µg/L) | SRP (µg/L) | TN (µg/L) | NO3− (µg/L) | NH4+ (µg/L) | Temperature (°C) | Alkalinity (meq/L) | pH (Units) |
|---|---|---|---|---|---|---|---|---|---|
| 1-Alserio | 5 | 31.9 ± 2.4 | 17.5 ± 3.4 | 2930 ± 333 | 1680 ± 543 | 320 ± 137 | 13.7 ± 2.8 | 3.8 ± 0.2 | 8.0 ± 0.1 |
| 2-Annone (W) | 4 | 27.1 ± 2.2 | 8.7 ± 0.5 | 870 ± 79 | 171 ± 82 | 87 ± 43 | 15.1 ± 4.0 | 2.9 ± 0.1 | 8.3 ± 0.1 |
| 3-Candia | 1 | 18.0 | 2.0 | 1060 | 170 | 17 | 27 | 1.18 | 7.5 |
| 4-Comabbio | 4 | 30.5 ± 5.5 | 16.2 ± 3.6 | 860 ± 105 | 158 ± 88 | 89 ± 75 | 15.2 ± 4.0 | 2.0 ± 0.1 | 8.4 ± 0.1 |
| 5-Maggiore | 4 | 7.6 ± 0.7 | 3.5 ± 0.6 | 885 ± 53 | 648 ± 61 | 10 ± 5 | 14.4 ± 3.5 | 0.9 ± 0.2 | 7.9 ± 0.2 |
| 6-Mergozzo | 4 | 5.0 ± 0.7 | 2.5 ± 0.3 | 851 ± 64 | 595 ± 36 | 15 ± 10 | 14.8 ± 3.8 | 0.3 ± 0.1 | 7.3 ± 0.3 |
| 7-Monate | 3 | 8.0 ± 1.3 | 2.0 ± 0.6 | 459 ± 13 | 158 ± 27 | 16 ± 6 | 13.6 ± 4.0 | 1.1 ± 0.2 | 7.9 ± 0.5 |
| 8-Montorfano | 7 | 18.3 ± 2.5 | 6.8 ± 0.6 | 936 ± 84 | 129 ± 50 | 245 ± 94 | 16.3 ± 2.9 | 1.9 ± 0.1 | 8.3 ± 0.1 |
| 9-Olginate | 4 | 18.0 ± 1.9 | 5.7 ± 1.8 | 942 ± 60 | 648 ± 79 | 39 ± 7 | 13.5 ± 3.5 | 1.4 ± 0.3 | 8.2 ± 0.3 |
| 10-Orta | 4 | 7.2 ± 1.4 | 3.2 ± 0.5 | 1537 ± 19 | 1313 ± 24 | 16 ± 7 | 14.7 ± 3.7 | 0.3 ± 0.1 | 7.2 ± 0.3 |
| 11-Piano | 1 | 63.0 | 8.0 | 2069 | 1424 | 677 | 11 | 3.8 | 8.4 |
| 12-Pusiano | 4 | 26.0 ± 4.6 | 3.6 ± 0.5 | 1313 ± 256 | 667 ± 189 | 133 ± 89 | 14.8 ± 4.4 | 2.6 ± 0.2 | 8.1 ± 0.2 |
| 13-Segrino | 7 | 10.7 ± 0.7 | 4.0 ± 0.5 | 1311 ± 195 | 751 ± 198 | 85 ± 16 | 11.4 ± 2.5 | 2.4 ± 0.1 | 7.9 ± 0.1 |
| 14-Varese | 5 | 50.4 ± 19.1 | 38.2 ± 18.3 | 901 ± 104 | 220 ± 71 | 117 ± 88 | 12.7 ± 3.1 | 2.5 ± 0.1 | 7.9 ± 0.1 |
| Model | Response Variable | Predictor | t | p |
|---|---|---|---|---|
| (a) | D25 | TP | −3.133 | 0.003 ** |
| TN | −2.974 | 0.004 ** | ||
| (b) | OxRate | NH4+ | 3.422 | 0.002 ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leoni, B.; Patelli, M.; Soler, V.; Nava, V. Ammonium Transformation in 14 Lakes along a Trophic Gradient. Water 2018, 10, 265. https://doi.org/10.3390/w10030265
Leoni B, Patelli M, Soler V, Nava V. Ammonium Transformation in 14 Lakes along a Trophic Gradient. Water. 2018; 10(3):265. https://doi.org/10.3390/w10030265
Chicago/Turabian StyleLeoni, Barbara, Martina Patelli, Valentina Soler, and Veronica Nava. 2018. "Ammonium Transformation in 14 Lakes along a Trophic Gradient" Water 10, no. 3: 265. https://doi.org/10.3390/w10030265
APA StyleLeoni, B., Patelli, M., Soler, V., & Nava, V. (2018). Ammonium Transformation in 14 Lakes along a Trophic Gradient. Water, 10(3), 265. https://doi.org/10.3390/w10030265