Time Variability Patterns of Eutrophication Indicators in the Bay of Algeciras (South Spain)

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
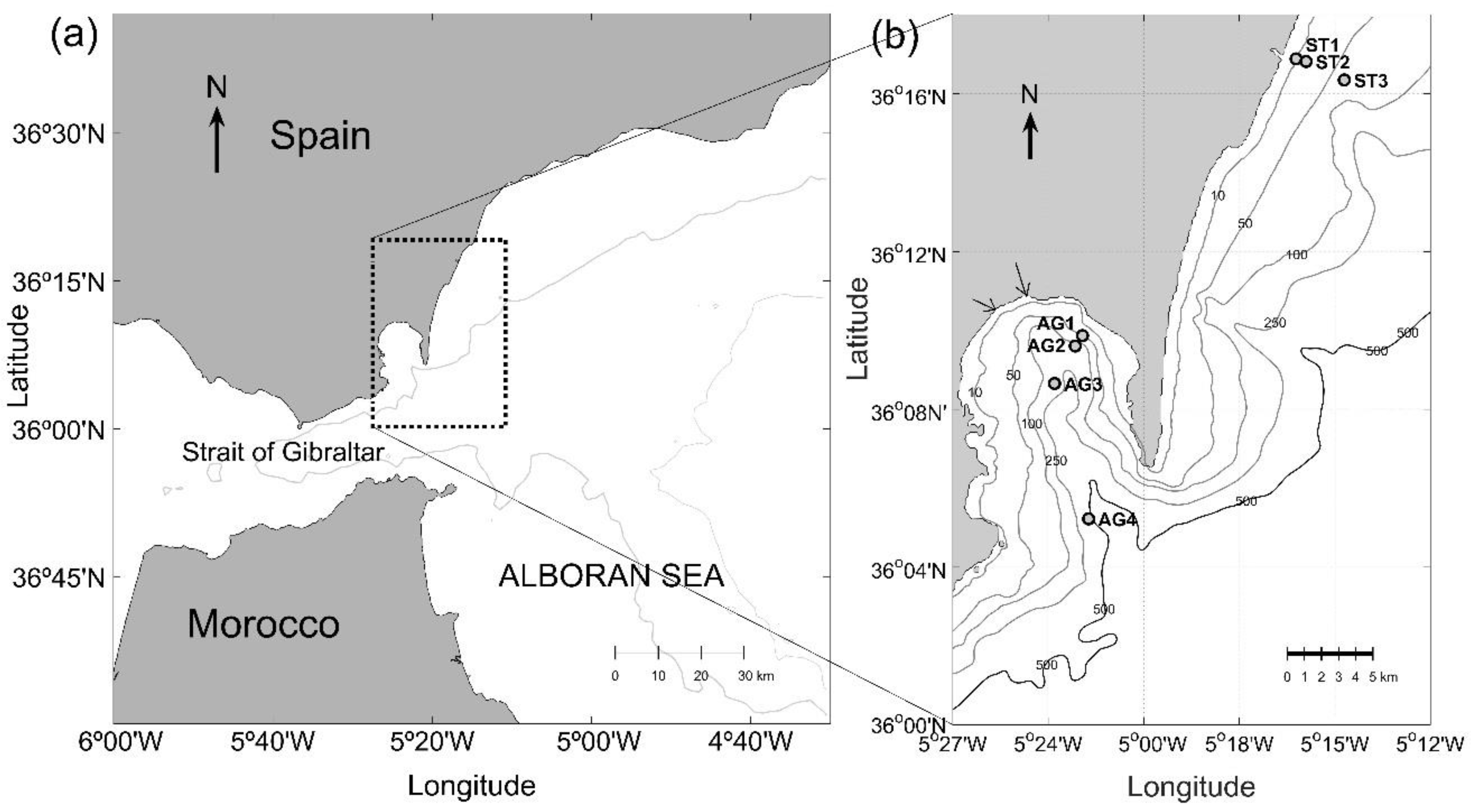
2.1. Study Area and Hydrographical Profiles.
2.2. Sampling Strategy and Analysis Procedures
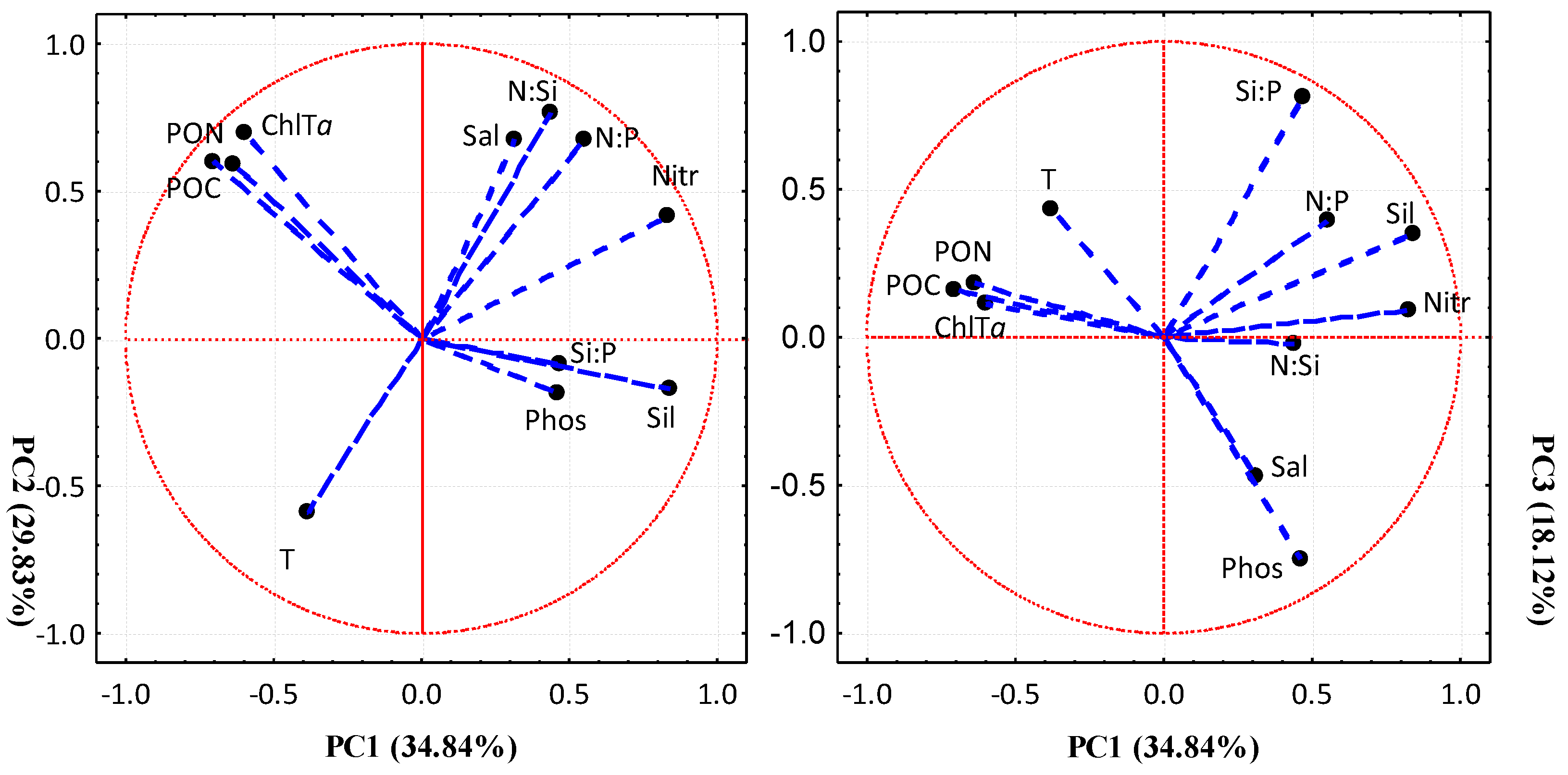
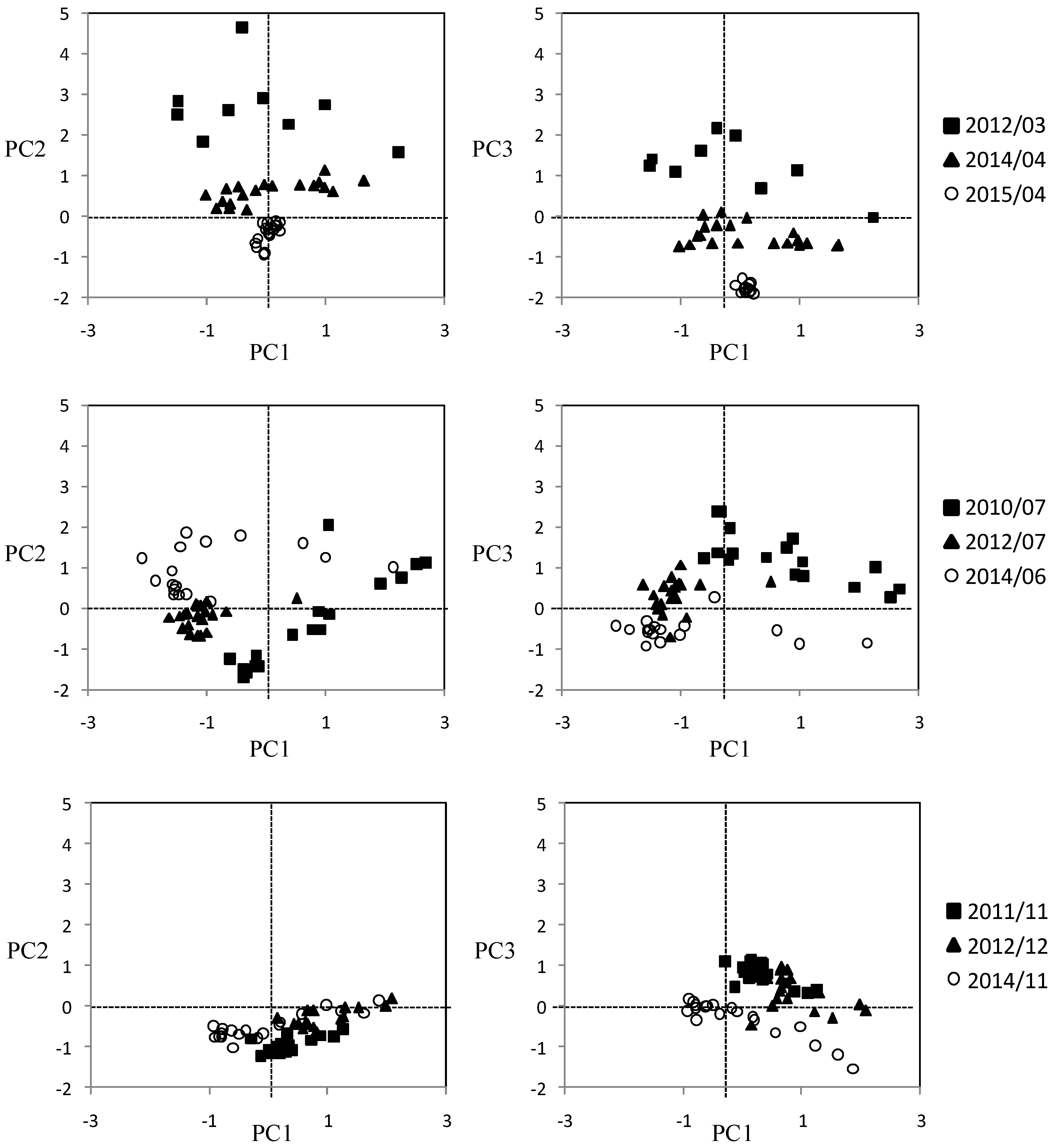
2.3. Statistical Analysis
3. Results
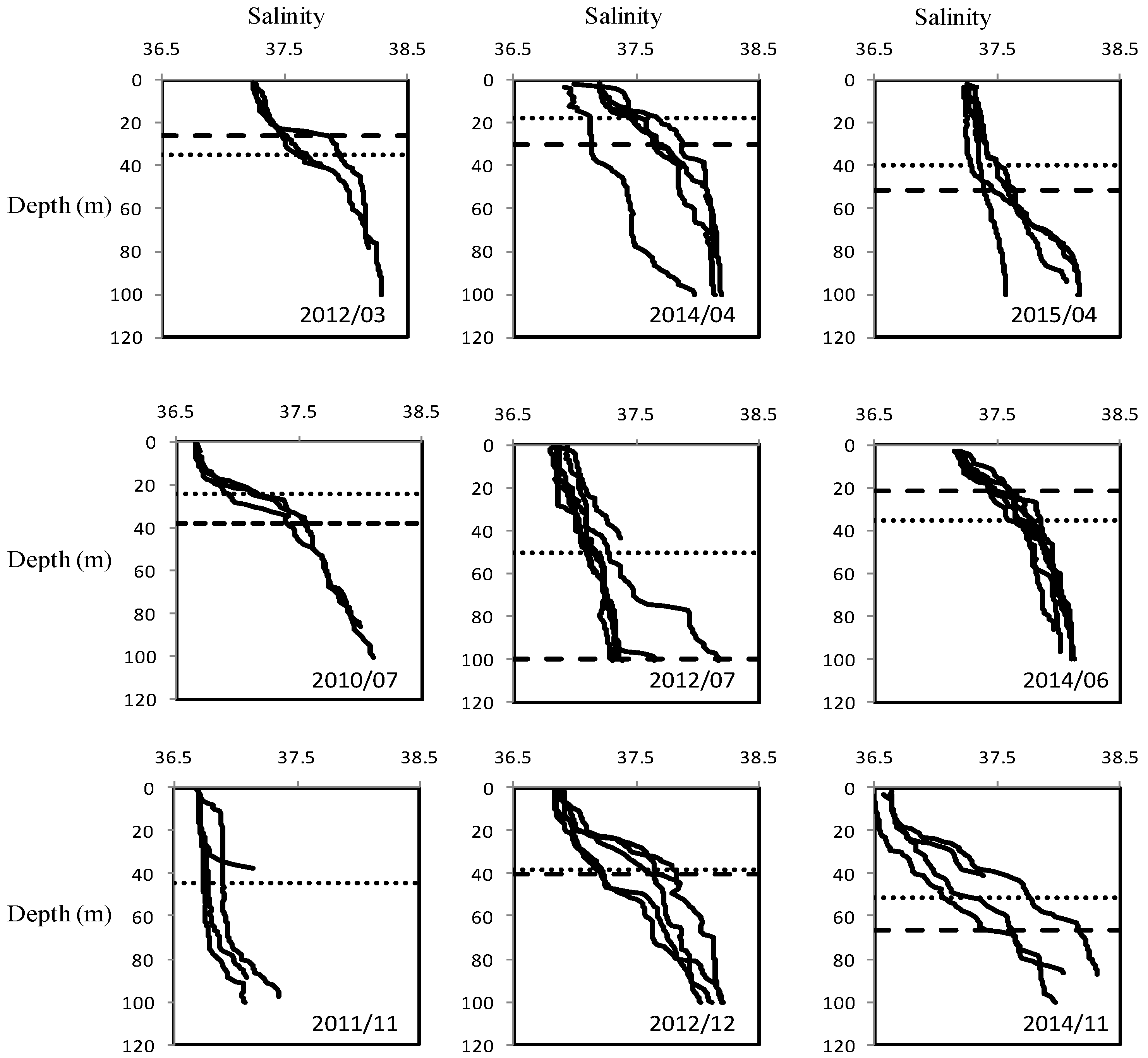
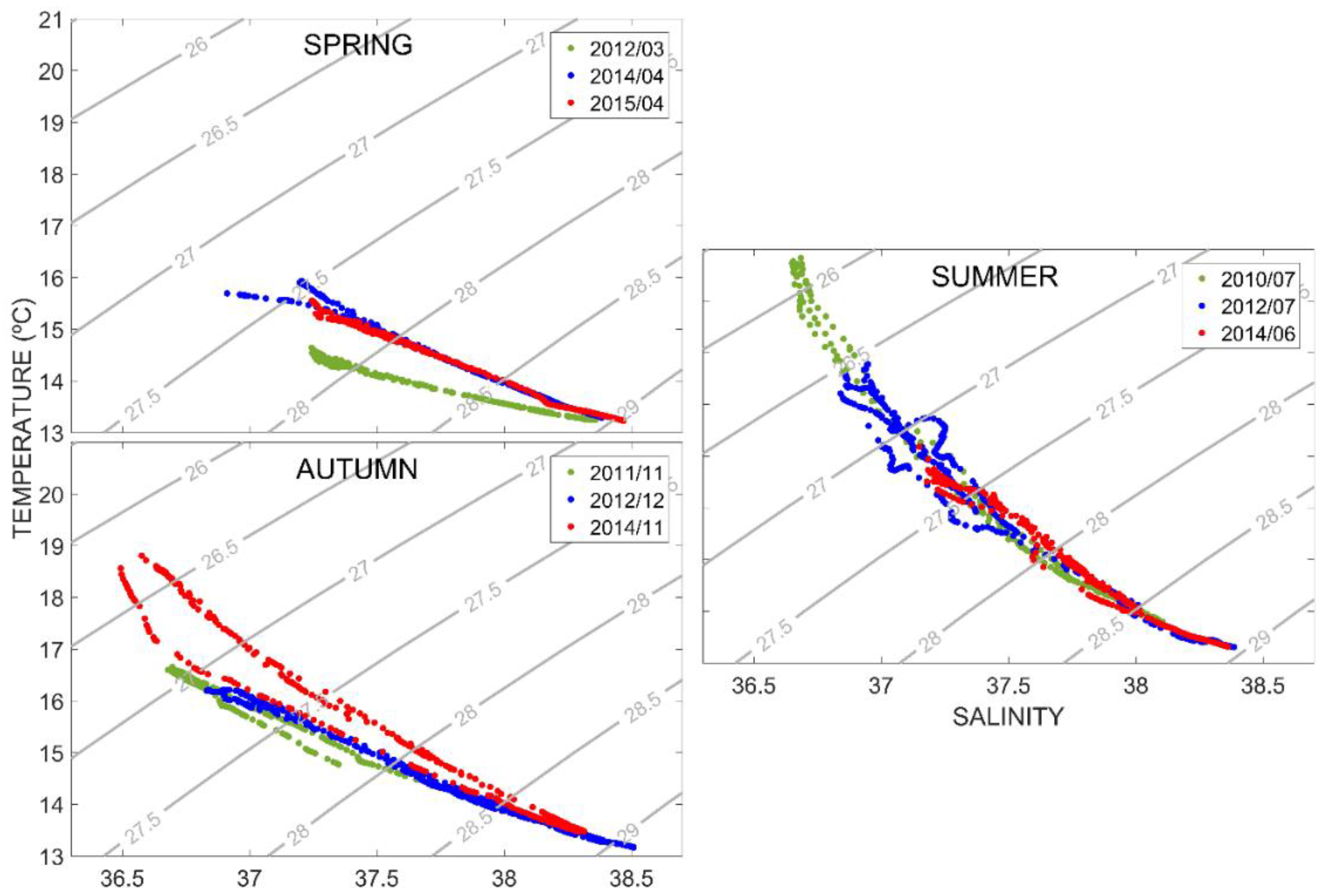
3.1. Temperature and Salinity
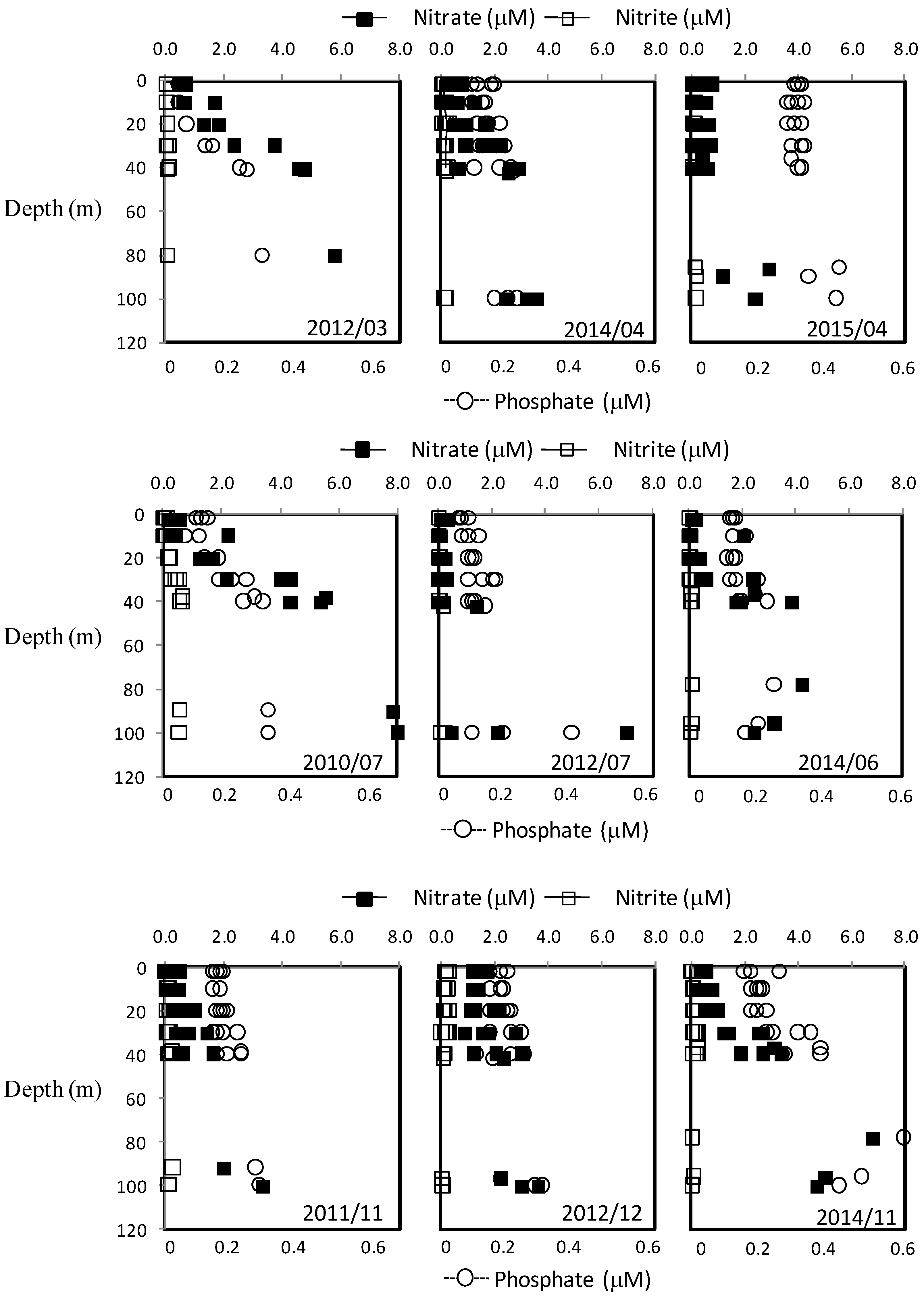
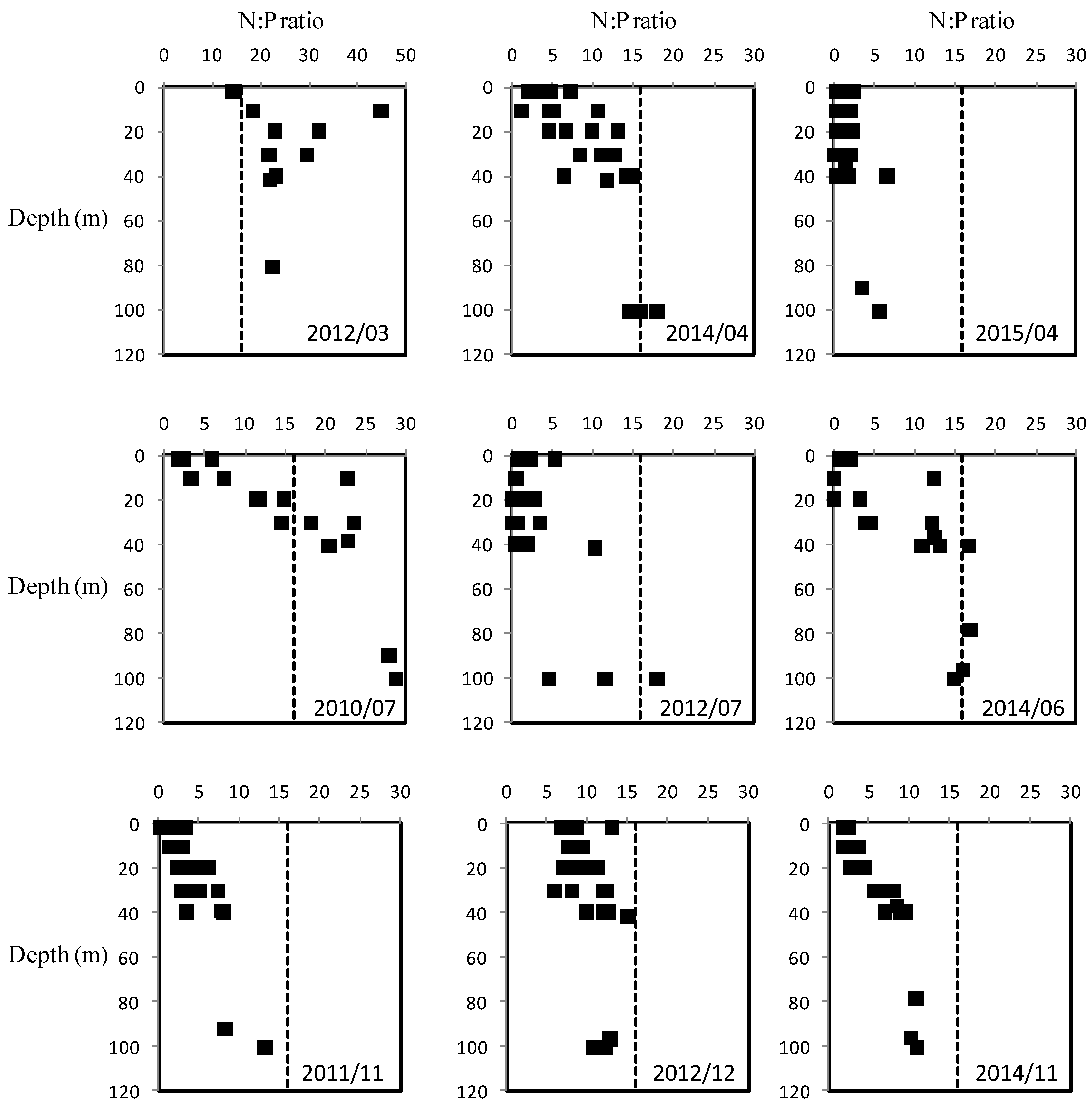
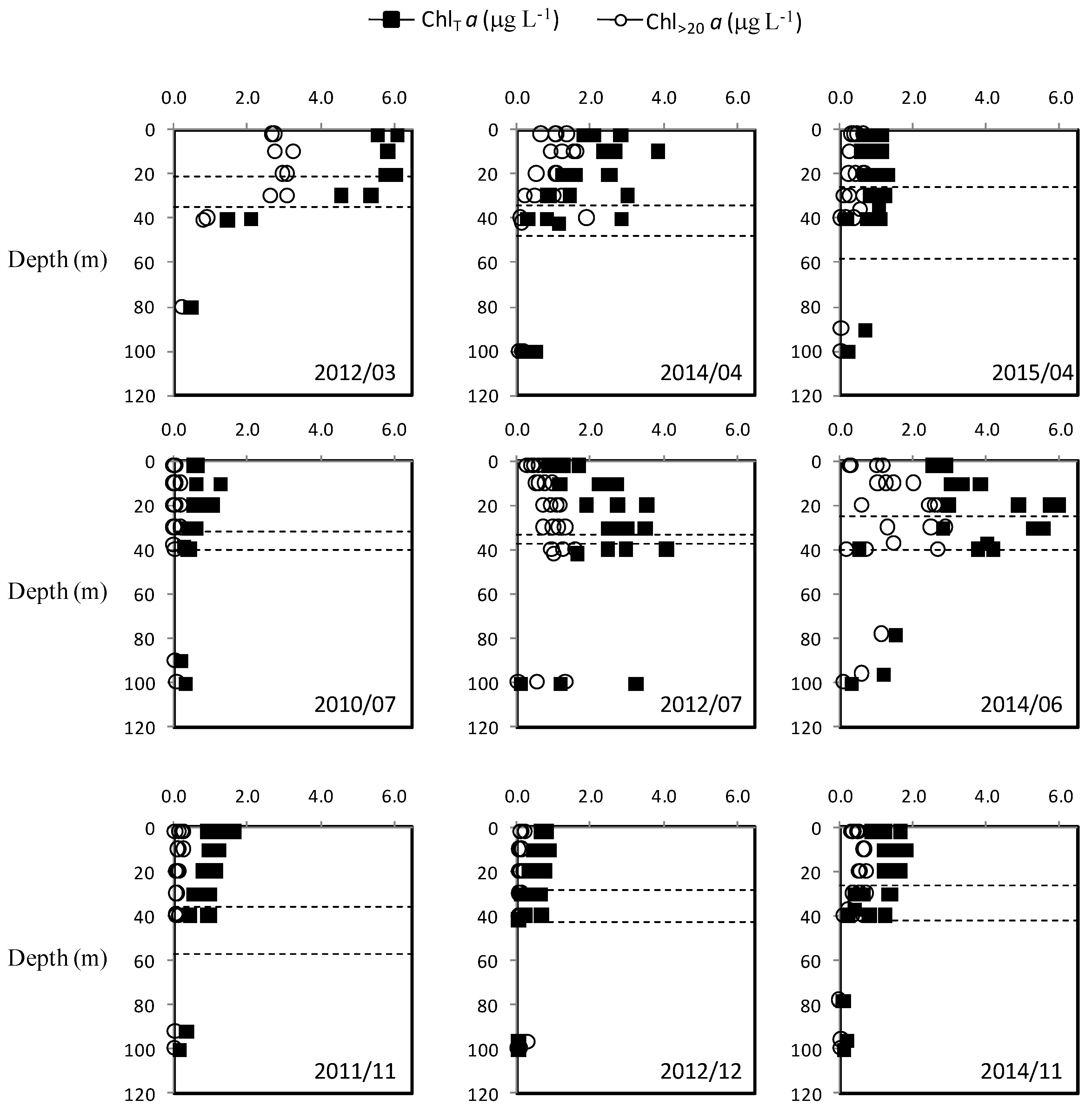
3.2. Vertical Profiles of Nutrients, Chlorophyll a and Particulate Matter
3.3. Seasonal Variability
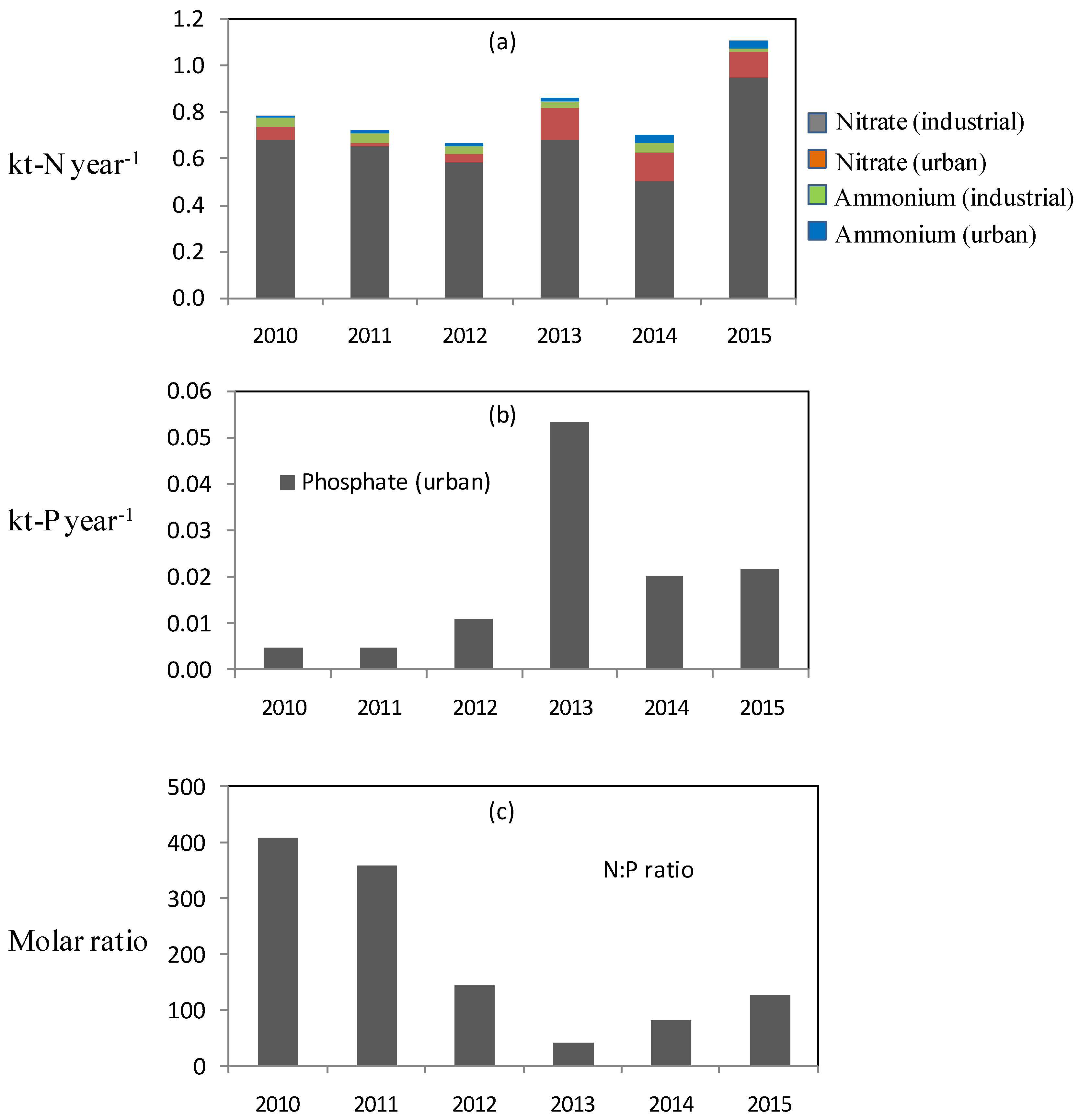
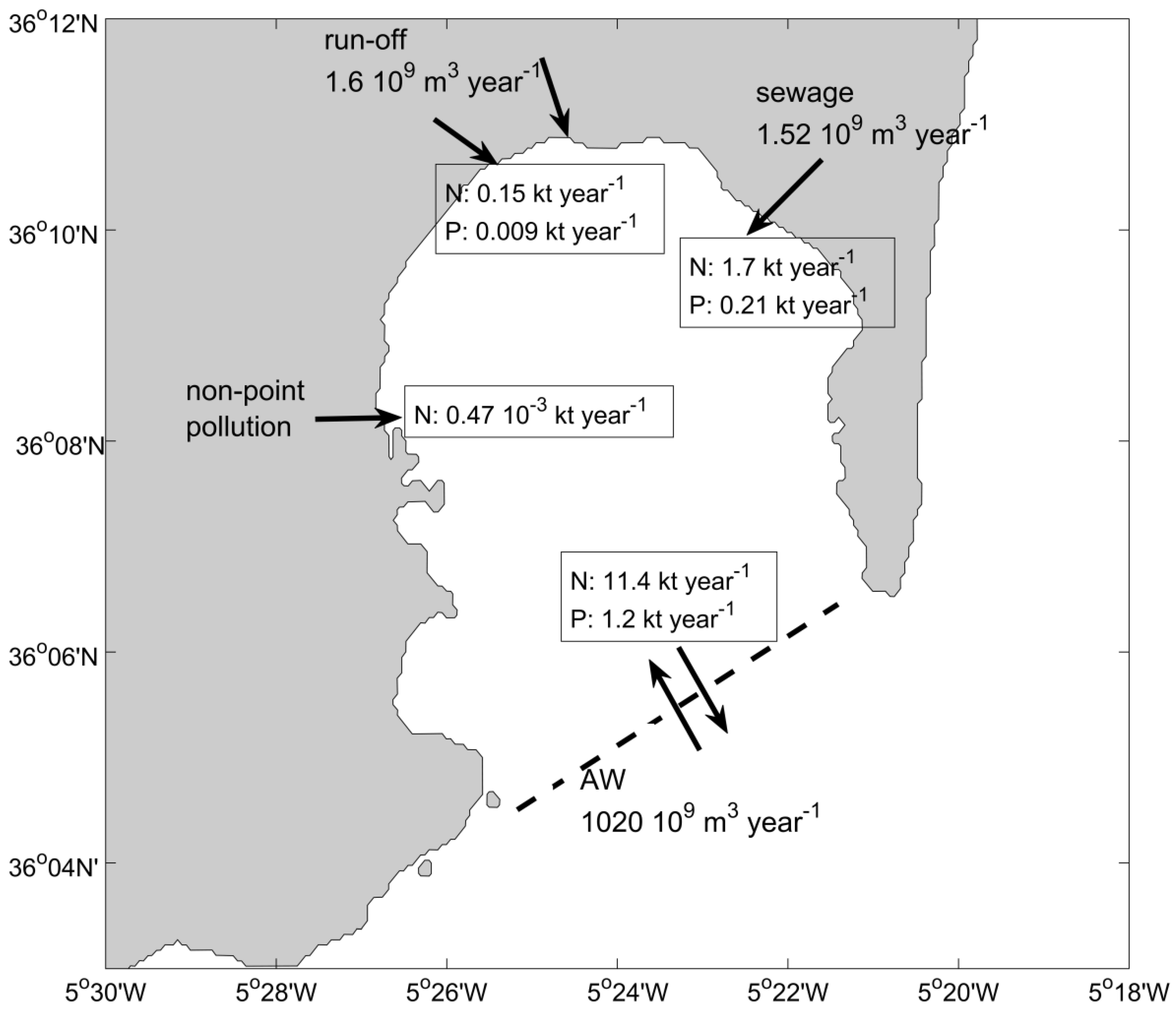
3.4. Nutrient Discharges from Terrestrial Sources
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- European Commission. Commission Decision of 1 September 2010 on criteria and methodological standards on good environmental status of marine waters (notified under document C(2010) 5956); (2010/477/EU)2010. Off. J. Eur. Union 2010, 232, 12–24. [Google Scholar]
- Ferreira, J.G.; Jesper, H.; Andersen, J.H.; Borja, A.; Bricker, S.B.; Camp, J.; da Silva, M.C.; Garcés, E.; Heiskanen, A.S.; Humborg, C.; et al. Overview of eutrophication indicators to assess environmental status within the European Marine Strategy Framework Directive. Estuar. Coast. Shelf Sci. 2011, 93, 117–131. [Google Scholar] [CrossRef]
- Harding, L.W.; Batiuk, R.A.; Fisher, T.R.; Gallegos, C.L.; Malone, T.C.; Miller, W.D.; Mulholland, M.R.; Paerl, H.W.; Perry, E.S.; Tango, P.J. Scientific bases for numerical chlorophyll criteria in Chesapeake Bay. Estuaries Coasts 2014, 37, 134–148. [Google Scholar] [CrossRef]
- Cloern, J.E. Our evolving conceptual model of the coastal eutrophication problem. Mar. Ecol. Prog. Ser. 2001, 210, 223–253. [Google Scholar] [CrossRef] [Green Version]
- McQuatters-Gollop, A.; Gilbert, A.J.; Mee, L.D.; Vermaat, J.E.; Artioli, Y.; Humborg, C.; Wulff, F. How well do ecosystem indicators communicate the effects of anthropogenic eutrophication? Estuar. Coast. Shelf Sci. 2009, 82, 583–596. [Google Scholar] [CrossRef]
- Alpine, A.E.; Cloern, J.E. Trophic interactions and direct physical effects control phytoplankton biomass and production in an estuary. Limnol. Oceanogr. 1992, 37, 946–955. [Google Scholar] [CrossRef] [Green Version]
- Balls, P.W.; Macdonald, A.; Pugh, K.; Edwards, A.C. Long-term nutrient enrichment of an estuarine system: Ythan, Scotland (1958-1993). Environ. Pollut. 1995, 90, 311–321. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Gilabert., J.; Gutiérrez, J.M.; Fernández, A.I.; Marcos, C.; Sabah, S. Evidence of a planktonic food web response to changes in nutrient input dynamics in the Mar Menor coastal lagoon, Spain. Hydrobiologia 2002, 475/476, 359–369. [Google Scholar]
- Pérez-Ruzafa, A.; Fernández, A.I.; Marcos, C.; Gilabert, J.; Quispe, J.I.; García-Charton, J.A. Spatial and temporal variations of hydrological conditions, nutrients and chlorophyll a in a Mediterranean coastal lagoon (Mar Menor, Spain). Hydrobiologia 2005, 550, 11–27. [Google Scholar] [CrossRef]
- Monbet, Y. Control of phytoplankton biomass in estuaries: A comparative analysis of microtidal and macrotidal estuaries. Estuaries 1992, 15, 563–571. [Google Scholar] [CrossRef]
- Li, W.K.W.; Lewis, M.R.; Harrison, W.G. Multiscalarity of the nutrient–chlorophyll relationship in coastal phytoplankton. Estuaries Coasts 2010, 33, 440–447. [Google Scholar] [CrossRef]
- Periáñez, R. Modelling the environmental behaviour of pollutants in Algeciras Bay (South Spain). Mar. Pollut. Bull. 2012, 64, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Garrido, J.C.; García-Lafuente, J.; Sammartino, S.; Naranjo, C.; de los Santos, F.J.; Álvarez-Fanjul, E. Meteorologically-driven circulation and flushing times of the Bay of Algeciras, Strait of Gibraltar. Mar. Pollut. Bull. 2014, 80, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Sammartino, S.; Sánchez-Garrido, J.C.; Naranjo, C.; García Lafuente, J.; Rodríguez Rubio, P.; Sotillo, M. Water renewal in semi-enclosed basins: A high resolution Lagrangian approach with application to the Bay of Algeciras, Strait of Gibraltar. Limnol. Oceanogr. Methods 2018, 16, 106–118. [Google Scholar] [CrossRef]
- Morillo, J.; Usero, J.; El Bakouri, H. Biomonitoring of heavy metals in the coastal waters of two industrialised bays in southern Spain using the barnacle Balanus amphitrite. Chem. Spec. Bioavailab. 2008, 20, 227–237. [Google Scholar] [CrossRef]
- Morales-Caselles, C.; Kalman, J.; Riba, I.; DelValls, T.A. Comparing sediment quality in Spanish littoral areas affected by acute (Prestige, 2002) and chronic (Bay of Algeciras) oil spills. Environ. Pollut. 2007, 146, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Moyano, J.E.; García-Adiego, E.M.; Estacio, F.; García-Gómez, J.C. Effect of environmental factors on the spatial variation of the epifaunal polychaetes of the alga Halopteris scoparia in Algeciras Bay (Strait of Gibraltar). Hydrobiologia 2002, 470, 133–148. [Google Scholar] [CrossRef]
- Guerra-García, J.M.; Baeza-Rojano, E.; Cabezas, M.P.; Díaz-Pavón, J.J.; Pacios, I.; García-Gómez, J.C. The amphipods Caprella penantis and Hyale schmidtii as biomonitors of trace metal contamination in intertidal ecosystems of Algeciras Bay, Southern Spain. Mar. Pollut. Bull. 2009, 58, 765–786. [Google Scholar] [CrossRef] [PubMed]
- Carballo, J.L.; Naranjo, S.A.; García, J.C. Use of marine sponges as stress indicators in marine ecosystems at Algeciras Bay (southern Iberian Peninsula). Mar. Ecol. Prog. Ser. 1996, 135, 109–122. [Google Scholar] [CrossRef] [Green Version]
- Mercado, J.M.; Cortés, D.; García, A.; Ramírez, T. Seasonal and inter-annual changes in the planktonic communities of the northwest Alboran Sea (Mediterranean Sea). Prog. Oceanogr. 2007, 74, 273–293. [Google Scholar] [CrossRef]
- Mercado, J.M.; Cortés, D.; Ramírez, T.; Gómez, F. Decadal weakening of the wind-induced upwelling reduces the impact of nutrient pollution in the Bay of Málaga (western Mediterranean Sea). Hydrobiologia 2012, 680, 91–107. [Google Scholar] [CrossRef]
- Kara, A.B.; Rochford, P.A.; Hulbur, H.E. Efficient and accurate bulk parameterizations of air-sea fluxes for use in general circulation models. J. Atmos. Ocean. Technol. 2000, 17, 1421–1438. [Google Scholar] [CrossRef]
- Mercado, J.M.; Ramírez, T.; Cortés, D.; Sebastián, M.; Liger, E.; Bautista, B. Partitioning the effects of changes in nitrate availability and phytoplankton community structure on the relative nitrate uptake in the Northwest Alboran Sea (Mediterranean Sea). Mar. Ecol. Prog. Ser. 2008, 359, 51–68. [Google Scholar] [CrossRef]
- Consejería de Medio Ambiente y Ordenación del Territorio. Memoria del Plan Hidrológico. Demarcación Hidrológica de las Cuencas Mediterráneas Andaluzas. Ciclo de Planificación Hidrológica 2015/2021; Junta de Andalucía: Seville, Spain, 2016; 458p. (In Spanish) [Google Scholar]
- Ramírez, T.; Cortés, D.; Mercado, J.M.; Vargas-Yañez, M.; Sebastián, M.; Liger, E. Seasonal dynamics of inorganic nutrients and phytoplankton biomass in the NW Alboran Sea. Estuar. Coast. Shelf Sci. 2005, 65, 654–670. [Google Scholar] [CrossRef]
- Savenkoff, C.; Chanut, J.P.; Vezina, A.F.; Gratton, Y. Distribution of biological-activity in the lower St. Lawrence estuary as determined by multivariate analysis. Estuar. Coast. Shelf Sci. 1995, 40, 647–664. [Google Scholar] [CrossRef]
- Minas, H.J.; Coste, B.; le Corre, P.; Minas, M.; Raimbault, P. Biological and geochemical signatures associated with the water circulation through the Strait of Gibraltar and in western Alboran Sea. J. Geophys. Res. 1991, 96, 8755–8771. [Google Scholar] [CrossRef]
- Sarhan, T.; García-Lafuente, J.; Vargas, M.; Vargas, J.M.; Plaza, P. Upwelling mechanisms in the northwestern Alborán Sea. J. Mar. Syst. 2000, 23, 317–331. [Google Scholar] [CrossRef]
- Tintoré, J.; Gomis, D.; Alonso, S.; Parrilla, G. Mesoscale dynamics and vertical motion in the Alboran Sea. J. Phys. Oceanogr. 1991, 21, 811–823. [Google Scholar] [CrossRef]
- Macías, D.; Martin, A.P.; García-Lafuente, J.; García, X.M.; Yool, A.; Brun, M.; Vázquez, A.; Izquierdo, A.; Sein, D.; Echevarría, F. Mixing and biogeochemical effects induced by tides at on the Atlantic-Mediterranean flow in the Strait of Gibraltar. Prog. Oceanogr. 2007, 74, 252–272. [Google Scholar]
- García-Górriz, E.; Carr, M.-E. Physical control of phytoplankton distributions in the Alboran Sea: A numerical and satellite approach. J. Geophys. Res. 2001, 106, 16795–16805. [Google Scholar] [CrossRef] [Green Version]
- Gómez, F.; Gorsky, G.; Garcia-Gorriz, E.; Picheral, M. Control of phytoplankton distribution in the Strait of Gibraltar by wind and fortnightly tides. Estuar. Coast. Shelf Sci. 2004, 59, 85–497. [Google Scholar] [CrossRef]
- Amorim, A.L.; León, P.; Mercado, J.M.; Cortés, D.; Gómez, F.; Putzeys, S.; Salles, S.; Yebra, L. Controls of picophytoplankton abundance and composition in a highly dynamic marine system, the Northern Alboran Sea (Western Mediterranean). J. Sea Res. 2016, 112, 13–22. [Google Scholar] [CrossRef]
- Coste, B.; le Corre, P.; Minas, H.J. Re-evaluation of the nutrient exchanges in the Strait of Gibraltar. Deep Sea Res. Part A 1988, 35, 767–775. [Google Scholar] [CrossRef]
- Dafner, E.V.; Boscolo, R.; Bryden, H.L. The N:Si:P molar ratio in the Strait of Gibraltar. Geophys. Res. Lett. 2003, 20, 1506. [Google Scholar] [CrossRef]
- Gómez, F.; Gorsky, G.; Striby, L.; Vargas, J.M.; González, N.; Picheral, M.; García-Lafuente, J.; Varela, M.; Goutx, M. Small-scale temporal variations in biogeochemical features in the Strait of Gibraltar, Mediterranean side—The role of NACW and the interface oscillation. J. Mar. Syst. 2001, 30, 207–220. [Google Scholar] [CrossRef]
- Béthoux, J.P.; Morin, P.; Ruiz-Pino, D.P. Temporal trends in nutrient ratios: Chemical evidence of Mediterranean ecosystem changes driven by human activity. Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 2007–2016. [Google Scholar] [CrossRef]
- Huertas, I.E.; Ríos, A.F.; García-Lafuente, J.; Navarro, G.; Makaoui, A.; Sánchez-Román, A.; Rodriguez-Galvez, S.; Orbi, A.; Ruíz, J.; Pérez, F.F. Atlantic forcing of the Mediterranean oligotrophy. Glob. Biogeochem. Cycles 2012, 26, GB2022. [Google Scholar] [CrossRef]
- Echevarría, F.; García-Lafuente, J.; Bruno, M.; Gorsky, G.; Goutx, M.; González, N.; García, C.M.; Gómez, F.; Vargas, J.M.; Picheral, M.; et al. Physical and biological coupling in the Strait of Gibraltar. Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 4115–4130. [Google Scholar] [CrossRef]
- Reul, A.; Rodríguez, V.; Jiménez-Gómez, F.; Blanco, J.M.; Bautista, B.; Sarhan, T.; Guerrero, F.; Ruíz, J.; García-Lafuente, J. Variability in the spatio-temporal distribution and size–structure of phytoplankton across an upwelling area in the NW Alboran Sea (W-Mediterranean). Cont. Shelf Res. 2005, 25, 589–608. [Google Scholar] [CrossRef]
- Gómez, F.; González, N.; Echevarría, F.; García, C.M. Distribution and fluxes of dissolved nutrients in the Strait of Gibraltar and its relationships to microphytoplankton biomass. Estuar. Coast. Shelf Sci. 2000, 51, 439–449. [Google Scholar]
- Mercado, J.M.; Ramírez, D.; Cortés, D. Changes in nutrient concentration induced by hydrological variability and its effects on light absorption by phytoplankton in the Alboran Sea (Western Mediterranean Sea). J. Mar. Syst. 2008, 71, 31–45. [Google Scholar] [CrossRef]
- Neale, P.J.; Sobrino, C.; Segovia, M.; Mercado, J.M.; León, P.; Cortés, D.; Tuite, P.; Picazo, A.; Salles, S.; Cabrerizo, M.J.; Prasil, O. Effects of CO2, nutrients and light on coastal plankton. I. Abiotic conditions and biological responses. Aquat. Biol. 2014, 22, 25–41. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Survey | Year | Data | Seasonal Period | Stations |
|---|---|---|---|---|
| EU0710 (10/07) | 2010 | 11 July | Stratification | AG1, AG2, AG3 |
| EU1111 (11/11) | 2011 | 25 November | Destratification | AG1, AG2, AG3, AG4 |
| EU0312 (12/03) | 2012 | 8 March | Bloom | AG1, AG2, AG3 |
| EU0712 (12/07) | 2012 | 14 July | Stratification | AG1, AG2, AG3, AG4 |
| EU1112 (12/12) | 2012 | 30 November | Destratification | AG1, AG2, AG3, AG4 |
| EU0414 (14/04) | 2014 | 19 April | Bloom | AG1, AG2, AG3, AG4 |
| EU0614 (14/06) | 2014 | 3 June | Stratification | AG1, AG2, AG3, AG4 |
| EU1114 (14/11) | 2014 | 27 October | Destratification | AG1, AG2, AG3, AG4 |
| EU0415 (15/04) | 2015 | 21 April | Bloom | AG1, AG2, AG3, AG4 |
| Spring | Summer | Autumn | |
|---|---|---|---|
| Surface temperature (°C) | 15.0 ± 0.8 | 17.8 ± 1.8 | 16.6 ± 0.5 |
| Surface salinity | 37.4 ± 0.3 | 37.0 ± 0.4 | 36.9 ± 0.2 |
| Temperature (°C) | 15.0 ± 0.5 | 17.5 ± 1.2 | 16.7 ± 1.1 |
| Salinity | 37.3 ± 0.1 | 37.1 ± 0.2 | 36.9 ± 0.2 |
| AMI (m) | 31.3 ± 18.7 | 75.4 ± 39.5 | 58.8 ± 32.5 |
| MLD (m) | 31.5 ± 16.9 | 34.3 ± 19.3 | 38.4 ± 20.1 |
| ELD (m) | 34.9 ± 10.1 | 39.3 ± 14.0 | 39.3 ± 8.9 |
| Nitrate (μM) | 0.87 ± 0.43 | 0.92 ± 1.03 | 1.03 ± 0.53 |
| Phosphate (μM) | 0.12 ± 0.13 | 0.12 ± 0.02 | 0.18 ± 0.03 |
| Silicate (μM) | 0.73 ± 0.11 | 0.96 ± 0.75 | 1.73 ± 0.61 |
| N:P molar ratio | 11.2 ± 12.0 | 5.4 ± 5.6 | 5.6 ± 3.3 |
| N:Si molar ratio | 1.2 ± 0.7 | 0.57 ± 0.35 | 0.63 ± 0.33 |
| Si:P molar ratio | 7.2 ± 5.5 | 7.8 ± 5.7 | 10.3 ± 5.0 |
| ChlT a (μg L−1) | 3.0 ± 2.6 | 2.2 ± 1.5 | 0.95 ± 0.34 |
| POC (μg L−1) | 226 ± 139 | 237 ± 106 | 130 ± 27 |
| PON (μg L−1) | 30 ± 16 | 28 ± 12 | 20 ± 4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mercado, J.M.; León, P.; Salles, S.; Cortés, D.; Yebra, L.; Gómez-Jakobsen, F.; Herrera, I.; Alonso, A.; Sánchez, A.; Valcárcel-Pérez, N.; et al. Time Variability Patterns of Eutrophication Indicators in the Bay of Algeciras (South Spain). Water 2018, 10, 938. https://doi.org/10.3390/w10070938
Mercado JM, León P, Salles S, Cortés D, Yebra L, Gómez-Jakobsen F, Herrera I, Alonso A, Sánchez A, Valcárcel-Pérez N, et al. Time Variability Patterns of Eutrophication Indicators in the Bay of Algeciras (South Spain). Water. 2018; 10(7):938. https://doi.org/10.3390/w10070938
Chicago/Turabian StyleMercado, Jesús M., Pablo León, Soluna Salles, Dolores Cortés, Lidia Yebra, Francisco Gómez-Jakobsen, Inma Herrera, Aitor Alonso, Antonio Sánchez, Nerea Valcárcel-Pérez, and et al. 2018. "Time Variability Patterns of Eutrophication Indicators in the Bay of Algeciras (South Spain)" Water 10, no. 7: 938. https://doi.org/10.3390/w10070938
APA StyleMercado, J. M., León, P., Salles, S., Cortés, D., Yebra, L., Gómez-Jakobsen, F., Herrera, I., Alonso, A., Sánchez, A., Valcárcel-Pérez, N., & Putzeys, S. (2018). Time Variability Patterns of Eutrophication Indicators in the Bay of Algeciras (South Spain). Water, 10(7), 938. https://doi.org/10.3390/w10070938