Beta Diversity Partitioning and Drivers of Variations in Fish Assemblages in a Headwater Stream: Lijiang River, China

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
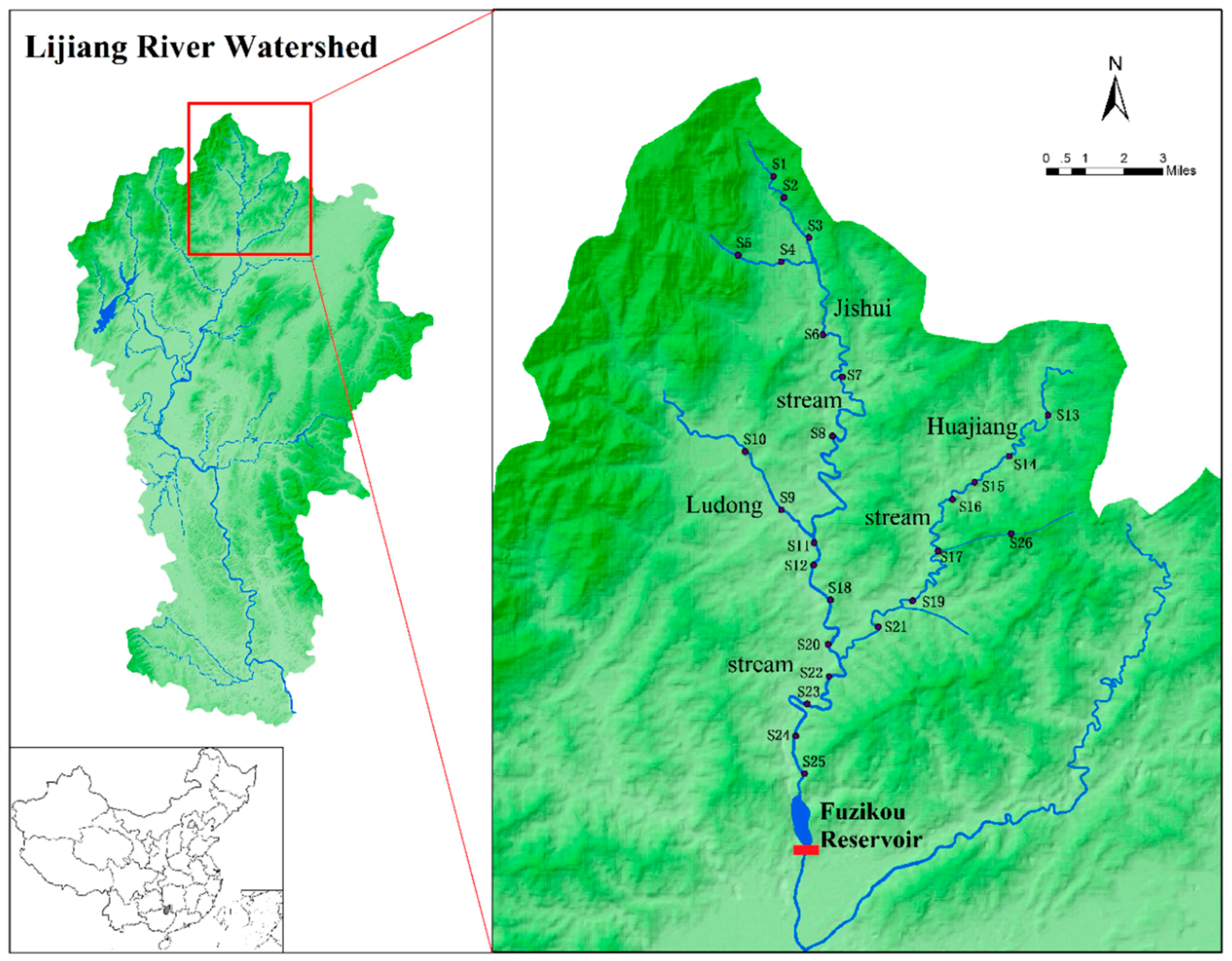
2.1. Study Area
2.2. Field Sample Collection
2.3. Biotic and Abiotic Datasets
2.4. Abiotic Datasets
2.5. Data Analysis
3. Results
3.1. Variability of Abiotic Variables
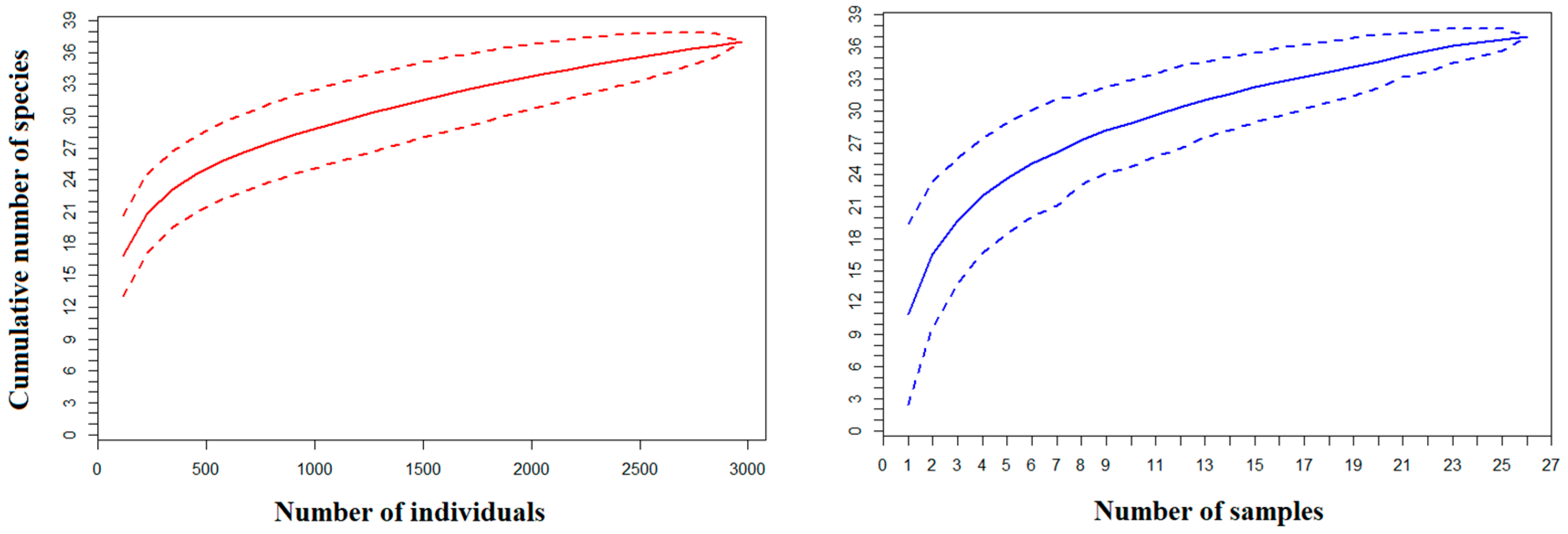
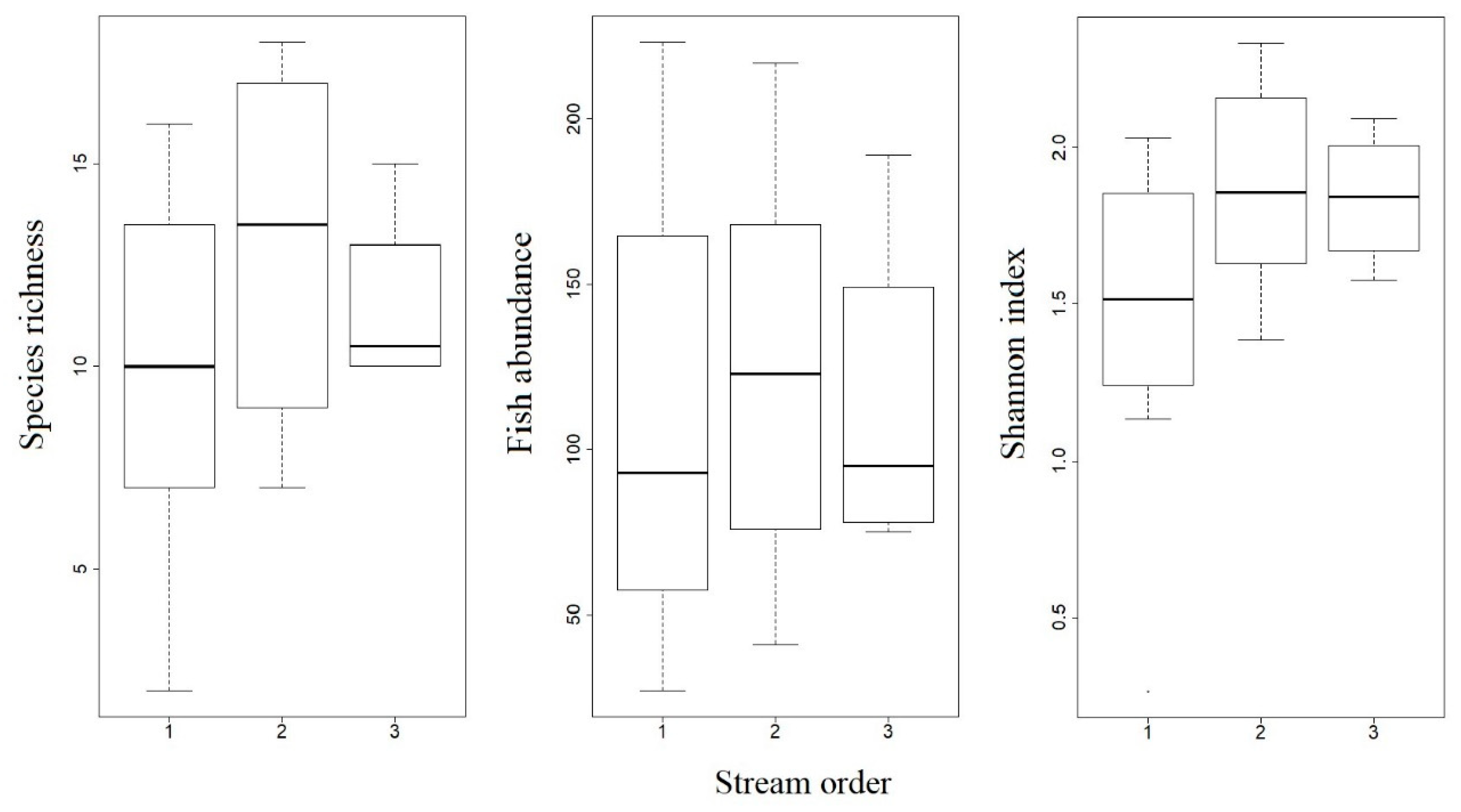
3.2. Fish Assemblage Composition
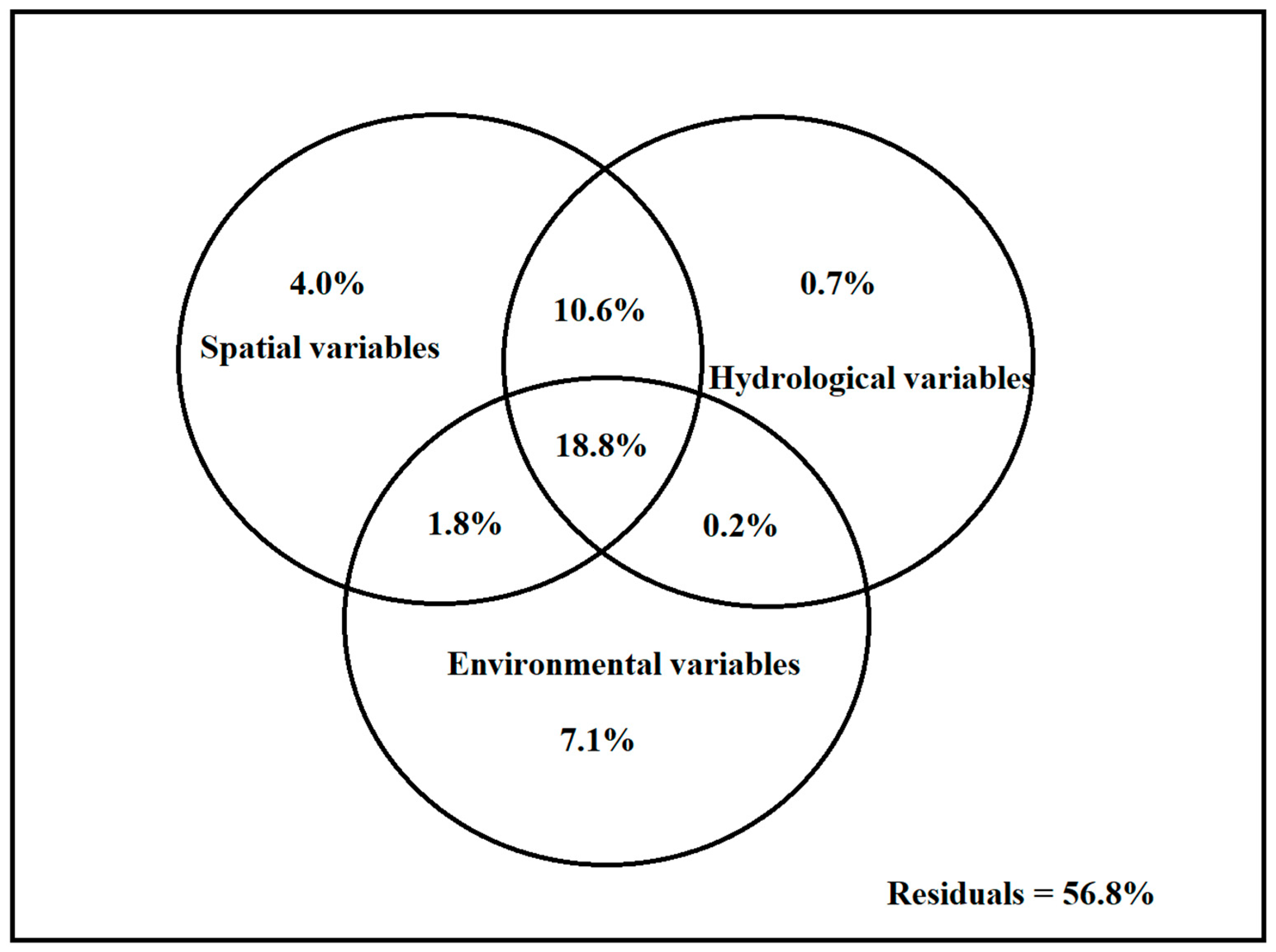
3.3. Driver of Species Composition
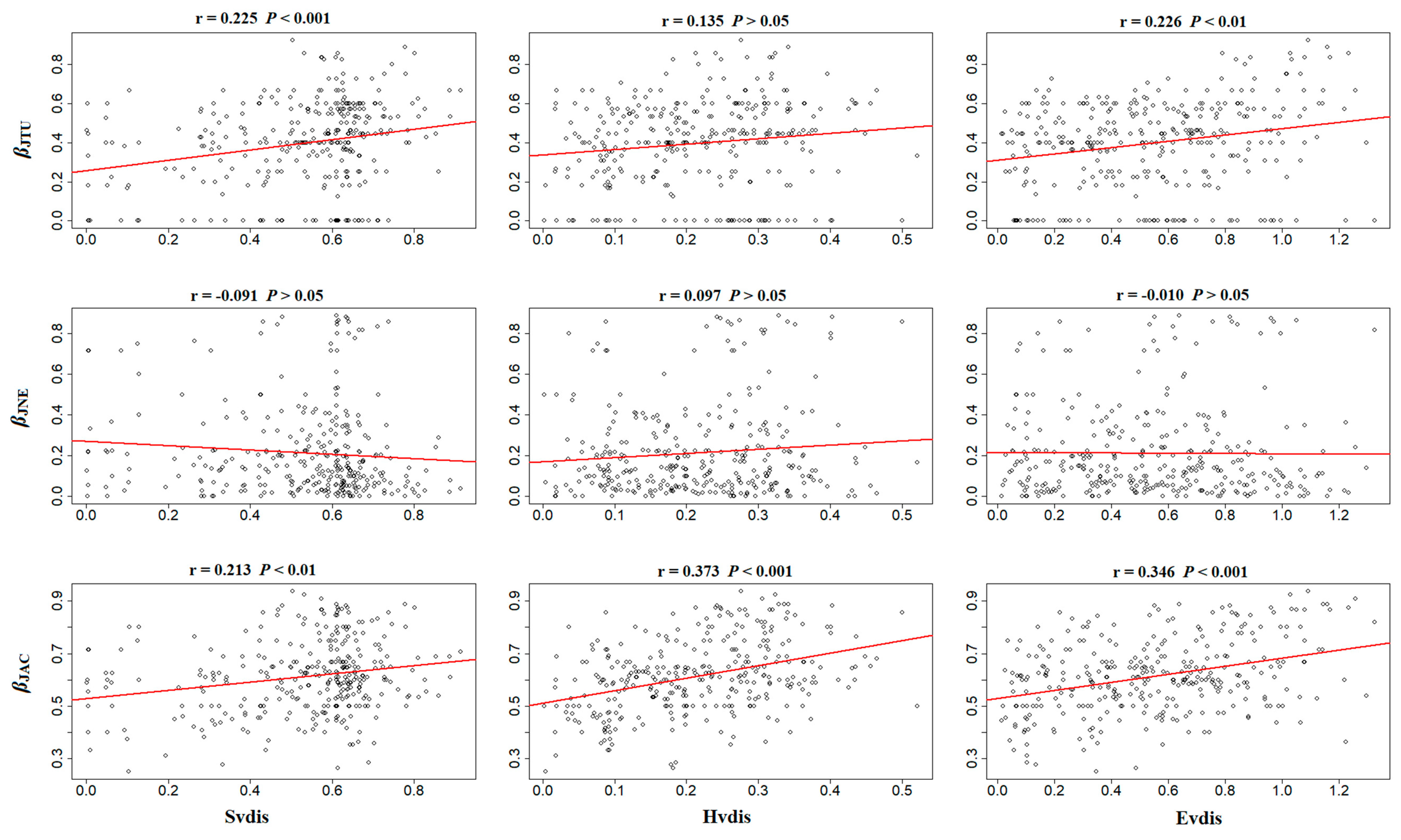
3.4. Main Driver of ß-Diversity
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Jowett, I.G.; Richardson, J. Fish communities in New Zealand rivers and their relationship to environmental variables. N. Z. J. Mar. Freshw. Res. 2003, 37, 347–366. [Google Scholar] [CrossRef] [Green Version]
- Zeni, J.O.; Casatti, L. The influence of habitat homogenization on the trophic structure of fish fauna in tropical streams. Hydrobiologia 2014, 726, 259–270. [Google Scholar] [CrossRef]
- Silvano, R.A.M.; do Amaral, B.D.; Oyakawa, O.T. Spatial and temporal patterns of diversity and distribution of the Upper Jurua River fish community (Brazilian Amazon). Environ. Biol. Fishes 2000, 57, 25–35. [Google Scholar] [CrossRef]
- Chick, J.H.; Pegg, M.A.; Koel, T.M. Spatial patterns of fish communities in the Upper Mississippi River System: Assessing fragmentation by low-head dams. River Res. Appl. 2006, 22, 413–427. [Google Scholar] [CrossRef]
- Li, J.; Huang, L.; Zou, L.; Kano, Y.; Sato, T.; Yahara, T. Spatial and temporal variation of fish assemblages and their associations to habitat variables in a mountain stream of north Tiaoxi River, China. Environ. Biol. Fishes 2012, 93, 403–417. [Google Scholar] [CrossRef]
- Bennett, M.G.; Kozak, J.P. Spatial and temporal patterns in fish community structure and abundance in the largest US river swamp, the Atchafalaya River floodplain, Louisiana. Ecol. Freshw. Fish 2016, 25, 577–589. [Google Scholar] [CrossRef]
- Espirito-Santo, H.M.V.; Zuanon, J. Temporary pools provide stability to fish assemblages in Amazon headwater streams. Ecol. Freshw. Fish 2017, 26, 475–483. [Google Scholar] [CrossRef]
- Pegg, M.A.; McClelland, M.A. Spatial and temporal patterns in fish communities along the Illinois River. Ecol. Freshw. Fish 2004, 13, 125–135. [Google Scholar] [CrossRef]
- Senay, C.; Taranu, Z.E.; Bourque, G.; Macnaughton, C.J.; Lanthier, G.; Harvey-Lavoie, S.; Boisclair, D. Effects of river scale flow regimes and local scale habitat properties on fish community attributes. Aquat. Sci. 2017, 79, 13–26. [Google Scholar] [CrossRef]
- Daufresne, M.; Boet, P. Climate change impacts on structure and diversity of fish communities in rivers. Glob. Chang. Biol. 2007, 13, 2467–2478. [Google Scholar] [CrossRef]
- Wyzga, B.; Amirowicz, A.; Radecki-Pawlik, A.; Zawiejska, J. Hydromorphological Conditions, Potential Fish Habitats and the Fish Community in a Mountain River Subjected to Variable Human Impacts, the Czarny Dunajec, Polish Carpathians. River Res. Appl. 2009, 25, 517–536. [Google Scholar] [CrossRef]
- Fischer, J.; Paukert, C.; Daniels, M. Fish Community Response to Habitat Alteration: Impacts of Sand Dredging in the Kansas River. Trans. Am. Fish. Soc. 2012, 141, 1532–1544. [Google Scholar] [CrossRef] [Green Version]
- Shukla, R.; Bhat, A. Beta-diversity partitioning and drivers of variations in tropical fish community structure in central India. Aquat. Sci. 2018, 80, 18. [Google Scholar] [CrossRef]
- Zbinden, Z.D.; Matthews, W.J. Beta diversity of stream fish assemblages: Partitioning variation between spatial and environmental factors. Freshw. Biol. 2017, 62, 1460–1471. [Google Scholar] [CrossRef]
- Fausch, K.D.; Torgersen, C.E.; Baxter, C.V.; Li, H.W. Landscapes to riverscapes: Bridging the gap between research and conservation of stream fishes. Bioscience 2002, 52, 483–498. [Google Scholar] [CrossRef]
- Leite, G.F.M.; Silva, F.T.C.; Goncalves, J.F.J.; Salles, P. Effects of conservation status of the riparian vegetation on fish assemblage structure in neotropical headwater streams. Hydrobiologia 2015, 762, 223–238. [Google Scholar] [CrossRef]
- Legendre, P. Interpreting the replacement and richness difference components of beta diversity. Glob. Ecol. Biogeogr. 2014, 23, 1324–1334. [Google Scholar] [CrossRef] [Green Version]
- Koleff, P.; Gaston, K.J.; Lennon, J.J. Measuring beta diversity for presence–absence data. J. Anim. Ecol. 2003, 72, 367–382. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Baselga, A.; Leprieur, F. Comparing methods to separate components of beta diversity. Methods Ecol. Evol. 2015, 6, 1069–1079. [Google Scholar] [CrossRef]
- Griffiths, D. Connectivity and vagility determine beta diversity and nestedness in North American and European freshwater fish. J. Biogeogr. 2017, 44, 1723–1733. [Google Scholar] [CrossRef]
- Vitorino, O.B.; Fernandes, R.; Agostinho, C.S.; Pelicice, F.M. Riverine networks constrain beta-diversity patterns among fish assemblages in a large Neotropical river. Freshw. Biol. 2016, 61, 1733–1745. [Google Scholar] [CrossRef]
- Leprieur, F.; Tedesco, P.A.; Hugueny, B.; Beauchard, O.; Durr, H.H.; Brosse, S.; Oberdorff, T. Partitioning global patterns of freshwater fish beta diversity reveals contrasting signatures of past climate changes. Ecol. Lett. 2011, 14, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.; He, D.M.; Perrett, L.; Wang, H.Y.; Hu, W.X.; Deng, W.D.; Wu, Y.F. Fish and fisheries in the Upper Mekong: Current assessment of the fish community, threats and conservation. Rev. Fish Biol. Fish. 2009, 19, 465–480. [Google Scholar] [CrossRef]
- Xie, J.Y.; Tang, W.J.; Yang, Y.H. Fish assemblage changes over half a century in the Yellow River, China. Ecol. Evol. 2018, 8, 4173–4182. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.C.; Zhang, C.G.; Fan, E.Y.; Zhao, Y.H. Freshwater fishes of China: Species richness, endemism, threatened species and conservation. Divers. Distrib. 2016, 22, 358–370. [Google Scholar] [CrossRef]
- Shuai, F.M.; Li, X.H.; Chen, F.C.; Li, Y.F.; Lek, S. Spatial patterns of fish assemblages in the Pearl River, China: Environmental correlates. Fundam. Appl. Limnol. 2017, 189, 329–340. [Google Scholar] [CrossRef]
- Yan, Y.Z.; Xiang, X.Y.; Chu, L.; Zhan, Y.J.; Fu, C.Z. Influences of local habitat and stream spatial position on fish assemblages in a dammed watershed, the Qingyi Stream, China. Ecol. Freshw. Fish 2011, 20, 199–208. [Google Scholar] [CrossRef]
- Kang, B.; Deng, J.M.; Wu, Y.F.; Chen, L.Q.; Zhang, J.; Qiu, H.Y.; Lu, Y.; He, D.M. Mapping China’s freshwater fishes: Diversity and biogeography. Fish Fish. 2014, 15, 209–230. [Google Scholar] [CrossRef]
- Kang, B.; Huang, X.X.; Yan, Y.Z.; Yan, Y.R.; Lin, H.D. Continental-scale analysis of taxonomic and functional fish diversity in the Yangtze river. Glob. Ecol. Conserv. 2018, 15, e00442. [Google Scholar] [CrossRef]
- Li, R.; Chen, Q.; Tonina, D.; Cai, D. Effects of upstream reservoir regulation on the hydrological regime and fish habitats of the Lijiang River, China. Ecol. Eng. 2015, 76, 75–83. [Google Scholar] [CrossRef]
- Wu, Z.; Huang, L. The Relationship between Fish Diversity and Environmental Variables; Science Press: Beijing, China, 2017. [Google Scholar]
- Zhu, Z.; Wu, Z.; Huang, L.; Feng, W.; Ding, Y. Species composition and diversity of fish in the upper reach of Lijiang River. Sichuan J. Zool. 2015, 34, 126–132. [Google Scholar]
- Anonymous. Freshwater Fishes of Guangxi, China, 2nd ed.; Guangxi People’s Publishing House: Nanning, China, 2006. [Google Scholar]
- Barbour, M.T.; Gerritsen, J.; Snyder, B.D.; Stribling, J.B. Rapid Bioassessment Protocols for Use in Streams and Wadable Rivers: Periphyton, Benthic Invertebrates and Fish, 2nd ed.; U.S. Environmental Protection Agency, Office of Water: Washington, DC, USA, 1999.
- Harrelson, C.C.; Rawlins, C.L.; Potyondy, J.P. Stream Channel Reference Sites: An Illustrated Guide to Field Technique; General Technical Report RM-245; U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1994.
- Baselga, A.; Orme, C.D.L. betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Svenning, J.C.; Baktoft, K.H.; Balslev, H. Land-use history affects understorey plant species distributions in a large temperate-forest complex, Denmark. Plant Ecolgy 2009, 201, 221–234. [Google Scholar] [CrossRef]
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Wu, N.; Qu, Y.; Guse, B.; Makarevičiūtė, K.; To, S.; Riis, T.; Fohrer, N. Hydrological and environmental variables outperform spatial factors in structuring species, trait composition, and beta diversity of pelagic algae. Ecol. Evol. 2018, 8, 2947–2961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Varation partitioning of species data matrices: Estimation and comparison of fractions. Ecology 2006, 87, 2614–2625. [Google Scholar] [CrossRef]
- Legendre, P. Studying beta diversity: Ecological variation partitioning by multiple regression and canonical analysis. J. Plant Ecol. 2008, 1, 3–8. [Google Scholar] [CrossRef]
- Baselga, A. The relationship between species replacement, dissimilarity derived from nestedness, and nestedness. Glob. Ecol. Biogeogr. 2012, 21, 1223–1232. [Google Scholar] [CrossRef] [Green Version]
- Sály, P.; Takács, P.; Kiss, I.; Bíró, P.; Erős, T. The relative influence of spatial context and catchment- and site-scale environmental factors on stream fish assemblages in a human-modified landscape. Ecol. Freshw. Fish 2011, 20, 251–262. [Google Scholar] [CrossRef]
- Godinho, F.N.; Ferreira, M.T.; Santos, J.M. Variation in fish community composition along an Iberian river basin from low to high discharge: Relative contributions of environmental and temporal variables. Ecol. Freshw. Fish 2000, 9, 22–29. [Google Scholar] [CrossRef]
- Magalhães, M.F.; Batalha, D.C.; Collares-Pereira, M.J. Gradients in stream fish assemblages across a Mediterranean landscape: Contributions of environmental factors and spatial structure. Freshw. Biol. 2002, 47, 1015–1031. [Google Scholar] [CrossRef]
- Matthews, W.J. Patterns in Freshwater Fish Ecology; Chapman & Hall: New York, NY, USA, 1998. [Google Scholar]
- Jackson, D.A.; Peres-Neto, P.R.; Olden, J.D. What controls who is where in freshwater fish communities—The roles of biotic, abiotic, and spatial factors. Can. J. Fish. Aquat. Sci. 2001, 58, 157–170. [Google Scholar]
- Fu, C.Z.; Wu, J.H.; Chen, J.K.; Qu, Q.H.; Lei, G.C. Freshwater fish biodiversity in the Yangtze River basin of China: Patterns, threats and conservation. Biodivers. Conserv. 2003, 12, 1649–1685. [Google Scholar] [CrossRef]
- Li, Y.R.; Tao, J.; Chu, L.; Yan, Y.Z. Effects of anthropogenic disturbances on alpha and beta diversity of fish assemblages and their longitudinal patterns in subtropical streams, China. Ecol. Freshw. Fish 2018, 27, 433–441. [Google Scholar] [CrossRef]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The River Continuum Concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Colwell, R.K.; Chao, A.; Gotelli, N.J.; Lin, S.-Y.; Mao, C.X.; Chazdon, R.L.; Longino, J.T. Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. J. Plant Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef] [Green Version]
- Chao, A. Non-parametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853. [Google Scholar] [CrossRef]
- Araújo, E.S.; Marques, E.E.; Freitas, I.S.; Neuberger, A.L.; Fernandes, R.; Pelicice, F.M. Changes in distance decay relationships after river regulation: Similarity among fish assemblages in a large Amazonian river. Ecol. Freshw. Fish 2013, 22, 543–552. [Google Scholar] [CrossRef]
- Pelicice, F.M.; Pompeu, P.S.; Agostinho, A.A. Large reservoirs as ecological barriers to downstream movements of Neotropical migratory fish. Fish Fish. 2015, 16, 697–715. [Google Scholar] [CrossRef]
- Cooper, A.R.; Infante, D.M.; Daniel, W.M.; Wehrly, K.E.; Wang, L.; Brenden, T.O. Assessment of dam effects on streams and fish assemblages of the conterminous USA. Sci. Total Environ. 2017, 586, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Socolar, J.B.; Gilroy, J.J.; Kunin, W.E.; Edwards, D.P. How Should Beta-Diversity Inform Biodiversity Conservation? Trends Ecol. Evol. 2016, 31, 67–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elements | Abbreviation |
|---|---|
| Environmental variables | Ev |
| Hydrological variables | Hv |
| Spatial variables | Sv |
| Dissimilarity matrices based on Euclidean distances for environmental variables | Evdis |
| Dissimilarity matrices based on Euclidean distances for hydrological variables | Hvdis |
| Dissimilarity matrices based on Euclidean distances for spatial variables | Svdis |
| Sørensen dissimilarity | ßSOR |
| Simpson dissimilarity (=turnover component of Sørensen dissimilarity) | ßSIM |
| Nestedness-resultant component of Sørensen dissimilarity | ßNES |
| Jaccard dissimilarity | ßJAC |
| Turnover component of Jaccard dissimilarity | ßJTU |
| Nestedness-resultant component of Jaccard dissimilarity | ßJNE |
| Abiotic Variables | Mean | Min. | Max. |
|---|---|---|---|
| Environmental variables (Ev) | |||
| Water temperature (°C) | 22.1 | 18.8 | 25.1 |
| pH | 7.8 | 6.9 | 9.7 |
| Dissolved oxygen (mg/L) | 8.4 | 6.1 | 9.8 |
| Conductivity (μS/cm) | 62.4 | 16.1 | 157.4 |
| Turbidity (NTU) | 3.1 | 0.2 | 18.4 |
| Substrate size (cm) | 10.2 | 7.2 | 15.9 |
| % Hydrostatic habitat (pool, plunge) | 28.4 | 0.0 | 76.9 |
| % Fastflow habitat (riffle, cascade) | 36.3 | 7.5 | 67.0 |
| % Slowflow habitat (run, glide) | 35.4 | 0.0 | 75.0 |
| Hydrological variables (Hv) | |||
| Wetted width (m) | 14.2 | 2.4 | 63.5 |
| Depth (cm) | 34.3 | 16.1 | 53.5 |
| Flow velocity (cm/s) | 31.2 | 14.2 | 59.4 |
| Distance from the dam of Fuzikou Reservoir (km) | 20.4 | 2.2 | 35.6 |
| Altitude (m) | 324.4 | 226.6 | 469.8 |
| Spatial variables (Sv) | |||
| Latitude (°N) | 25.78 | 25.70 | 25.88 |
| Longitude (°E) | 110.46 | 110.50 | 110.57 |
| PCNM1: Principal coordinates of neighborhood matrix 1 | 0.00 | −0.25 | 0.32 |
| PCNM2: Principal coordinates of neighborhood matrix 2 | 0.00 | −0.29 | 0.34 |
| PCNM3: Principal coordinates of neighborhood matrix 3 | 0.00 | −0.38 | 0.37 |
| PCNM4: Principal coordinates of neighborhood matrix 4 | 0.00 | −0.42 | 0.37 |
| PCNM5: Principal coordinates of neighborhood matrix 5 | 0.00 | −0.25 | 0.48 |
| PCNM6: Principal coordinates of neighborhood matrix 6 | 0.00 | −0.42 | 0.39 |
| PCNM7: Principal coordinates of neighborhood matrix 7 | 0.00 | −0.62 | 0.42 |
| PCNM8: Principal coordinates of neighborhood matrix 8 | 0.00 | −0.37 | 0.40 |
| PCNM9: Principal coordinates of neighborhood matrix 9 | 0.00 | −0.5 | 0.36 |
| PCNM10: Principal coordinates of neighborhood matrix 10 | 0.00 | −0.43 | 0.39 |
| PCNM11: Principal coordinates of neighborhood matrix 11 | 0.00 | −0.61 | 0.45 |
| PCNM12: Principal coordinates of neighborhood matrix 12 | 0.00 | −0.71 | 0.70 |
| PCNM13: Principal coordinates of neighborhood matrix 13 | 0.00 | −0.30 | 0.56 |
| PCNM14: Principal coordinates of neighborhood matrix 14 | 0.00 | −0.50 | 0.76 |
| PCNM15: Principal coordinates of neighborhood matrix 15 | 0.00 | −0.40 | 0.50 |
| PCNM16: Principal coordinates of neighborhood matrix 16 | 0.00 | −0.61 | 0.43 |
| PCNM17: Principal coordinates of neighborhood matrix 17 | 0.00 | −0.42 | 0.82 |
| Fish Species | Fish Abundance (N) |
|---|---|
| Bagridae | |
| 1. Tachysurus adiposalis (Oshima, 1919) | 7 |
| 2. Tachysurus albomarginatus (Rendahl, 1928) | 34 |
| Balitoridae | |
| 3. Erromyzon sinensis (Chen, 1980) | 312 |
| 4. Pseudogastromyzon fangi (Nichols, 1931) | 736 |
| 5. Vanmanenia lineata (Fang, 1935) | 26 |
| 6. Vanmanenia pingchowensis (Fang, 1935) | 86 |
| Cichlidae | |
| 7. Oreochromis niloticus (Linnaeus, 1758) * | 2 |
| Cobitidae | |
| 8. Misgurnus anguillicaudatus (Cantor, 1842) | 36 |
| Cyprinidae | |
| 9. Acrossocheilus kreyenbergii (Regan, 1908) | 19 |
| 10. Acrossocheilus parallens (Nichols, 1931) | 381 |
| 11. Carassius auratus (Linnaeus, 1758) | 1 |
| 12. Cyprinus carpio Linnaeus, 1758 | 5 |
| 13. Microphysogobio chenhsienensis (Fang, 1938) | 29 |
| 14. Microphysogobio kiatingensis (Wu, 1930) | 3 |
| 15. Onychostoma barbatulum (Pellegrin, 1908) | 1 |
| 16. Onychostoma gerlachi (Peters, 1881) | 1 |
| 17. Opsariichthys bidens Günther, 1873 | 88 |
| 18. Parasinilabeo assimilis Wu & Yao, 1977 | 16 |
| 19. Pseudorasbora parva (Temminck & Schlegel, 1846) | 12 |
| 20. Rhodeus ocellatus (Kner, 1866) | 1 |
| 21. Squalidus atromaculatus (Nichols & Pope, 1927) | 27 |
| 22. Zacco platypus (Temminck & Schlegel, 1846) | 623 |
| Gobiidae | |
| 23. Rhinogobius duospilus (Herre, 1935) | 110 |
| 24. Rhinogobius filamentosus (Wu, 1939) | 4 |
| 25. Rhinogobius similis Gill, 1859 | 143 |
| 26. Rhinogobius leavelli (Herre, 1935) | 183 |
| Mastacembelidae | |
| 27. Macrognathus aculeatus (Bloch, 1786) | 2 |
| Nemacheilidae | |
| 28. Oreonectes platycephalus Günther, 1868 | 2 |
| 29. Schistura fasciolata (Nichols & Pope, 1927) | 22 |
| 30. Schistura incerta (Nichols, 1931) | 9 |
| 31. Traccatichthys pulcher (Nichols & Pope, 1927) | 1 |
| Percichthyidae | |
| 32. Coreoperca whiteheadi Boulenger, 1900 | 25 |
| 33. Siniperca scherzeri Steindachner, 1892 | 1 |
| 34. Siniperca undulata Fang & Chong, 1932 | 1 |
| Siluridae | |
| 35. Pterocryptis anomala (Herre, 1933) | 2 |
| 36. Pterocryptis cochinchinensis (Valenciennes, 1840) | 1 |
| Sisoridae | |
| 37. Glyptothorax fokiensis (Rendahl, 1925) | 16 |
| Abiotic Variables | F | P | Abiotic Variables | F | P |
|---|---|---|---|---|---|
| Environmental variables (Ev) | 4.52 | <0.001 | Spatial variables (Sv) | 4.39 | <0.001 |
| Turbidity | 4.55 | 0.005 | PCNM1 | 9.61 | 0.005 |
| Conductivity | 3.93 | 0.005 | PCNM2 | 3.81 | 0.01 |
| Hydrological variables (Hv) | 4.89 | <0.001 | PCNM3 | 2.77 | 0.025 |
| Altitude | 7.93 | 0.005 | PCNM8 | 3.16 | 0.015 |
| Depth | 1.95 | 0.03 |
| Index | Evdis | Hvdis | Svdis | Evdis a | Hvdis a | Svdis a |
|---|---|---|---|---|---|---|
| ßSIM | 0.269 ** | 0.155 * | 0.232 *** | 0.188 * | 0.034 | 0.128 |
| ßSNE | 0.033 | 0.150 | −0.058 | 0.047 | 0.162 | −0.085 |
| ßSOR | 0.356 *** | 0.366 *** | 0.200 ** | 0.281 * | 0.255 * | 0.042 |
| ßJTU | 0.226 ** | 0.135 | 0.225 *** | 0.154 * | 0.028 | 0.139 |
| ßJNE | −0.010 | 0.097 | −0.091 | 0.018 | 0.126 | −0.099 |
| ßJAC | 0.346 *** | 0.373 *** | 0.213 ** | 0.264 * | 0.264 * | 0.061 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, L.; Huang, J.; Wu, Z.; Mo, Y.; Zou, Q.; Jeppesen, E.; Wu, N. Beta Diversity Partitioning and Drivers of Variations in Fish Assemblages in a Headwater Stream: Lijiang River, China. Water 2019, 11, 680. https://doi.org/10.3390/w11040680
Huang L, Huang J, Wu Z, Mo Y, Zou Q, Jeppesen E, Wu N. Beta Diversity Partitioning and Drivers of Variations in Fish Assemblages in a Headwater Stream: Lijiang River, China. Water. 2019; 11(4):680. https://doi.org/10.3390/w11040680
Chicago/Turabian StyleHuang, Liangliang, Jian Huang, Zhiqiang Wu, Yuanmin Mo, Qi Zou, Erik Jeppesen, and Naicheng Wu. 2019. "Beta Diversity Partitioning and Drivers of Variations in Fish Assemblages in a Headwater Stream: Lijiang River, China" Water 11, no. 4: 680. https://doi.org/10.3390/w11040680
APA StyleHuang, L., Huang, J., Wu, Z., Mo, Y., Zou, Q., Jeppesen, E., & Wu, N. (2019). Beta Diversity Partitioning and Drivers of Variations in Fish Assemblages in a Headwater Stream: Lijiang River, China. Water, 11(4), 680. https://doi.org/10.3390/w11040680