1. Introduction
Deltas and estuaries are complex ecosystems largely recognized for their productivity and importance, both for the economy and the conservation of nature (e.g., [
1,
2,
3]). However, the rapid growth of human population has put these areas under increasing pressure that threatens their ecological integrity and economic value [
4]. In Mediterranean aquatic ecosystems, the impacts produced by anthropogenic pressures are magnified by their increased, and often extreme, natural hydrological variability [
5]. Human response to such hydrological fluctuations includes flow regulation and water extraction that frequently disrupt aquatic ecosystems and produce accentuated environmental stress [
6,
7,
8].
There is a need to harmonize nature conservation with socioeconomic activities. The provision of environmental flow regimes (hereafter, e-flows) in the context of the Brisbane Declaration [
9,
10] “to sustain aquatic ecosystems which, in turn, support human cultures, economies, sustainable livelihoods, and well-being” constitutes an essential tool towards this direction. The 2012 European Commission’s “Blueprint to Safeguard Europe’s Water Resources” proposed the development of a guidance document [
11] in the framework of a Common Implementation Strategy (CIS) that would provide a European definition of e-flows and a common understanding of how they should be calculated in general terms. Such a document constituted a complement for the EU Water Framework Directive (WFD) [
12], which established the objective of achieving a good ecological status (which categories may be high, good, moderate, poor, or bad) in European water courses.
The ecological status of rivers in the European Union must be assessed through ecological indices (Water Framework Directive; [
12]). Those based on fish offer the best sensitiveness to hydromorphological pressures [
13,
14], such as hydrological variations. The official index in Spain is the Mediterranean Index of Biotic Integrity (IBIMED), based on the Index of Biotic Integrity for Catalan rivers (IBICAT2010) [
15] and intercalibrated in Europe [
16].
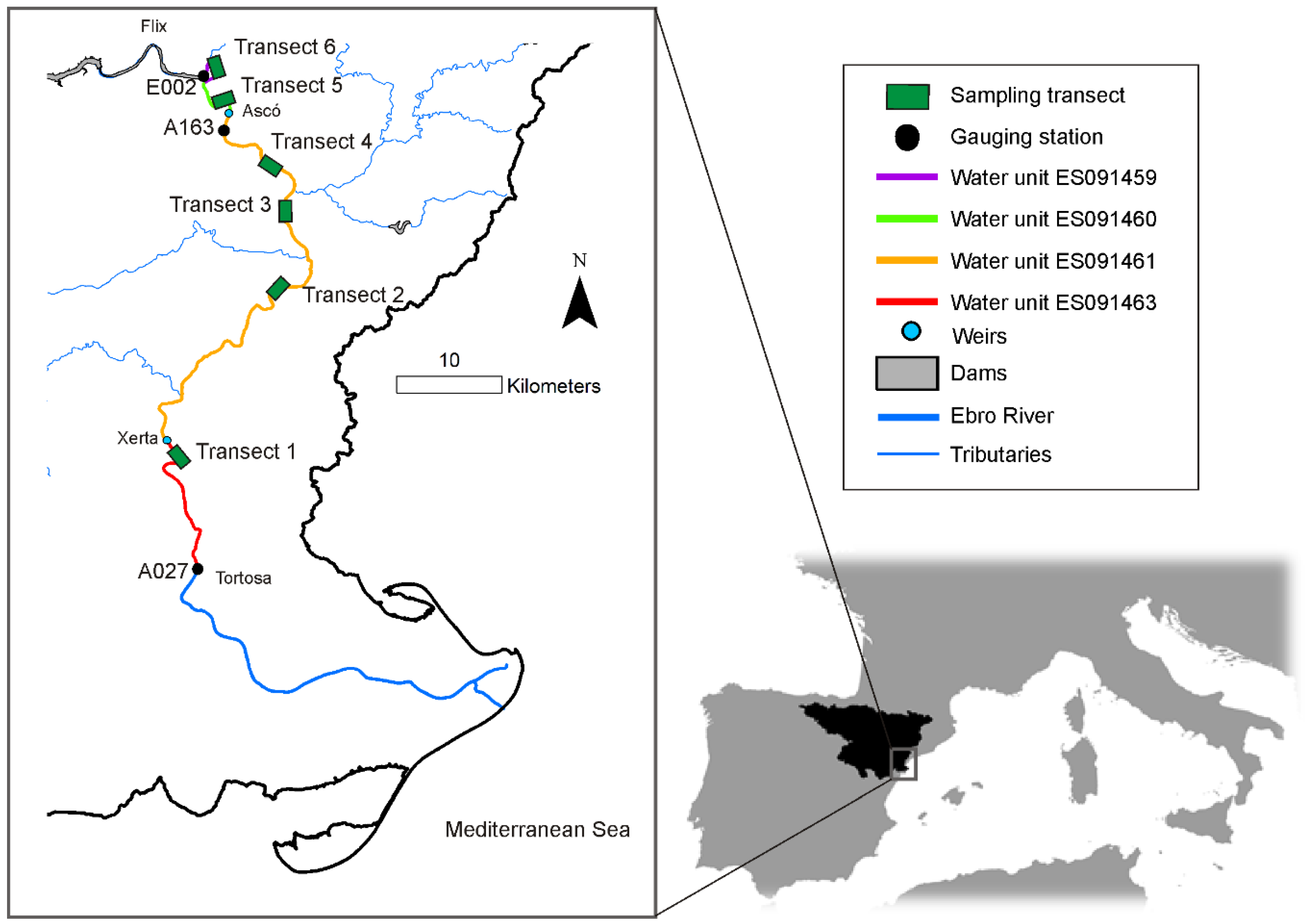
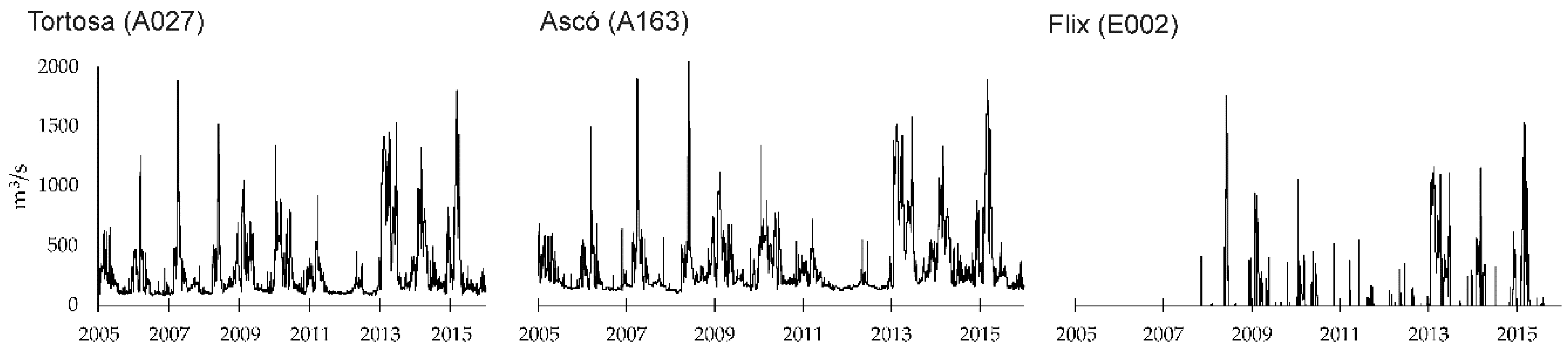
Studies developed in the lower Ebro have linked fish-based ecological quality or alien fish species with flow regimes. Using data from summer 2016, Caiola et al. [
17] found that the success (establishment and dispersal) of alien fishes is enhanced by flow reduction, which resulted in decreased flow velocity in the littoral zone. They determined that water velocities lower than 0.4 m/s are associated with an impacted community dominated by alien species (i.e., alien-to-native species ratio greater than 0.5). Belmar et al. [
18] used interannual field data sampled between 2006 and 2015 in the lower Ebro and found relationships between hydrological indices associated with the magnitude and variability of flow regimes (using daily and hourly flow records) and ecological quality, assessed using different fish-based indices. They concluded that the index IBICAT2010 [
15] is more suitable than IBICAT2b (its variant) [
15] and the Improvement and Spatial extension of the European Fish Index (EFI+) [
19,
20] to detect ecohydrological relationships in the lower Ebro River, depending on the spatial and temporal scales considered. On one side, this further suggested that the time period considered to characterize hydrologic regimes determined the ability to observe relationships between flow indices and ecological quality. On the other side, the ability to detect such ecohydrological relationships depended on the location of the transect, even with those located within the same water unit (“masa de agua”; subdivision of surface waters to implement the WFD in Spain).
The e-flows proposed for the lower Ebro (
Table 1) have little in common with the requirements of natural ecosystems or natural flow regimes, as they are not an exception to the common tendency of water management to set minimum flows that are constant for long periods [
21]. The Hydrological Plan published in 1996 established such e-flows as the 10% of the natural mean interannual runoff (5% when the mean flow was greater than 80 m
3/s) in the river, maintaining a provision of 100 m
3/s for the mouth. In 2001, the National Hydrological Plan (PHN, in Spanish) developed the Integral Protection Plan of the Ebro Delta (PIPDE, in Spanish) to maintain its “special ecological conditions” but e-flows were defined using the same criteria as in 1996. This simplistic approach was criticized arguing that the classical methods of determining environmental flows in rivers are neither designed nor adequate for the objective of maintaining the deltas and estuaries in a good ecological status [
22]. In 2007, the Commission for the Sustainability of the Ebro Land (CSTE, in Catalan), including representatives from the Catalan and Spanish governments, issued a report proposal of monthly e-flows for the river. In addition, the Royal Decree of Hydrological Planning [
23] and the Technical Instruction of Hydrological Planning [
24] developed normative contents regarding e-flow assessments. The Ebro’s Hydrological Plan for 2010–2015, approved in 2014, defined e-flows for the lower part of the river, much lower than those proposed by CSTE in 2007 and 2015 [
25]. Finally, the Hydrological Plan for the period 2016–2021 used the same environmental regime (
Table 1). Therefore, the e-flows for the lower Ebro are based solely on defining flow magnitudes. Although such magnitudes may change among months and types of year, there are not explicit rules to adjust flow variability (e.g., coefficient of variation).
Advances in measuring flow-ecology relationships allow inferring conclusions on the ecological impacts of specific flow regimes on communities, including e-flows, but further research is still necessary in order to determine the patterns of habitat complexity that may explain differences in ecohydrological relationships (
sensu [
26]) among river transects (and water units). Exceptional rises in magnitude, together with a general reduction in flow magnitude and variability, are common in Mediterranean main stems (such as the lower Ebro) because of dam management (with consequences on river habitats) [
27]. Given the relevance of flow magnitude in e-flow regimes (
Table 1), basing such flow-ecology relationships on mean discharges results relevant from a water management perspective (see examples in [
28,
29]). In addition, the sustainability of deltas cannot be guaranteed only with the allocation of e-flows for the fish inhabiting the low section of the river, which is the current practice in Spain and most countries. E-flows must be determined not only for the river ecosystem but also for the associated coastal and marine systems, which represents a challenge for scientists and water managers [
22]. The possible effects of water quality (e.g., nutrient content) have also to be considered. In this context, a relationship between freshwater inputs from the Ebro River and coastal fishery species such as anchovy (
Engraulis encransicolus) has been highlighted [
30]. Major river outflows are one of the nutrient enrichment processes that maintain fishery production worldwide [
31], and similar results have been obtained in other areas in Mexico [
32] and Australia [
33]. In this context, the littoral of the Ebro delta is suitable to study the relationship between river outflows and two shellfish fishery species: prawn (
Penaeus kerathurus) and mantis shrimp (
Squilla mantis). Due to their commercial value and their dependence on deltaic habitats, the delta is the only fishing ground of these species exploited by the corresponding fishermen’s society (the most important in Catalonia in terms of income). Therefore, significant relationships between river outflows and these species would have socioeconomic implications relevant for water management at catchment scale (particularly, for e-flow assessment). In addition, the delta is characterized as one of the main feeding and breeding areas of several endangered bird species, amongst which Audouin’s gull (
Larus audouinii) stands out. Determining relationships between flow regimes and bird communities in the delta and in similar Mediterranean areas, as it has been done in (scarce) studies developed in other areas [
34], may be essential for nature conservation.
The present study, developed in the lower Ebro and its delta, aimed to (i) deepen into the ecohydrological relationships found in Belmar et al. [
18] in order to obtain additional conclusions for water managers; and (ii) complete such conclusions using other groups of organisms relevant for ecological conservation and socioeconomic activities at the delta and its littoral zone [
22]. This will also provide conclusions useful for water management. Specific tasks were planned to:
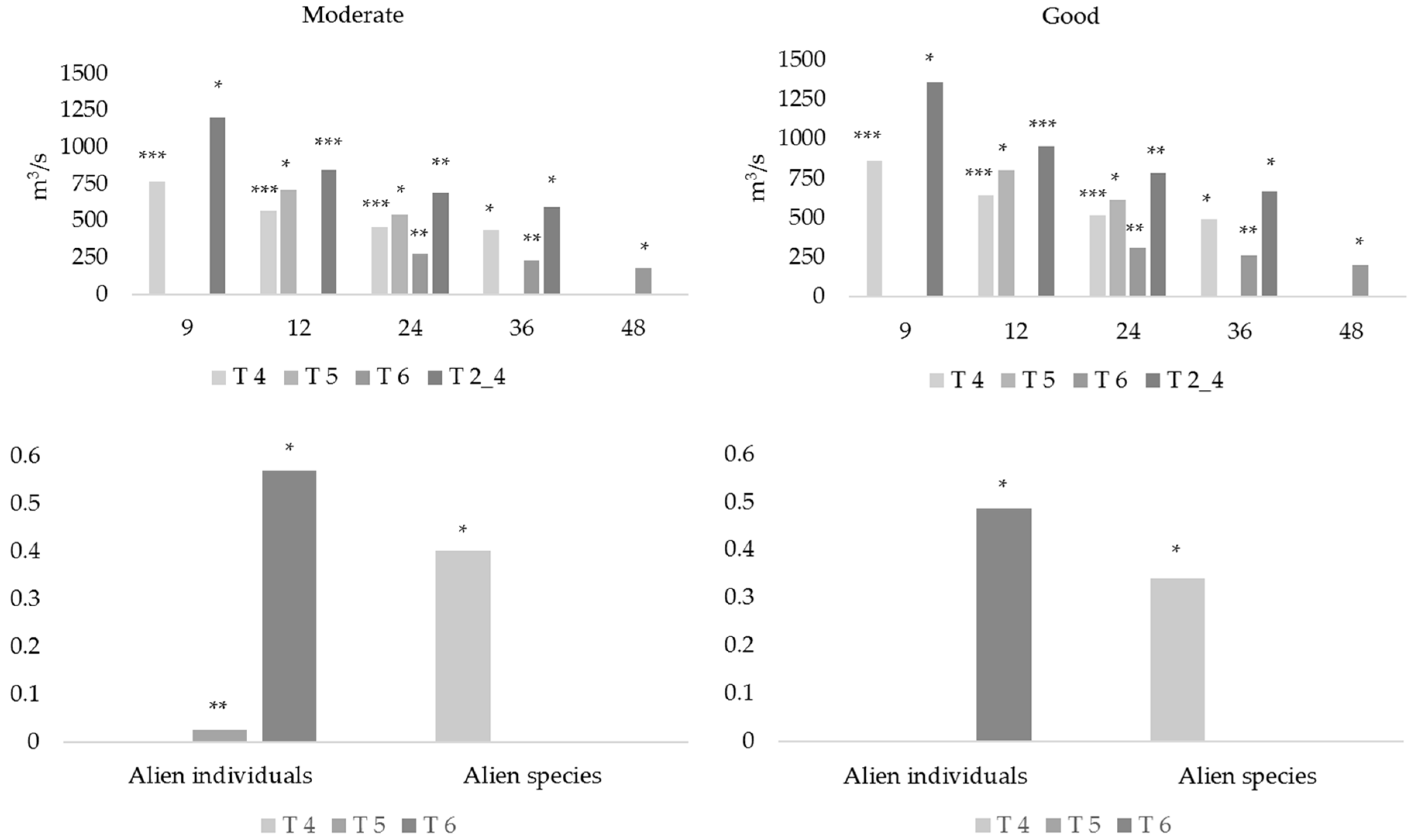
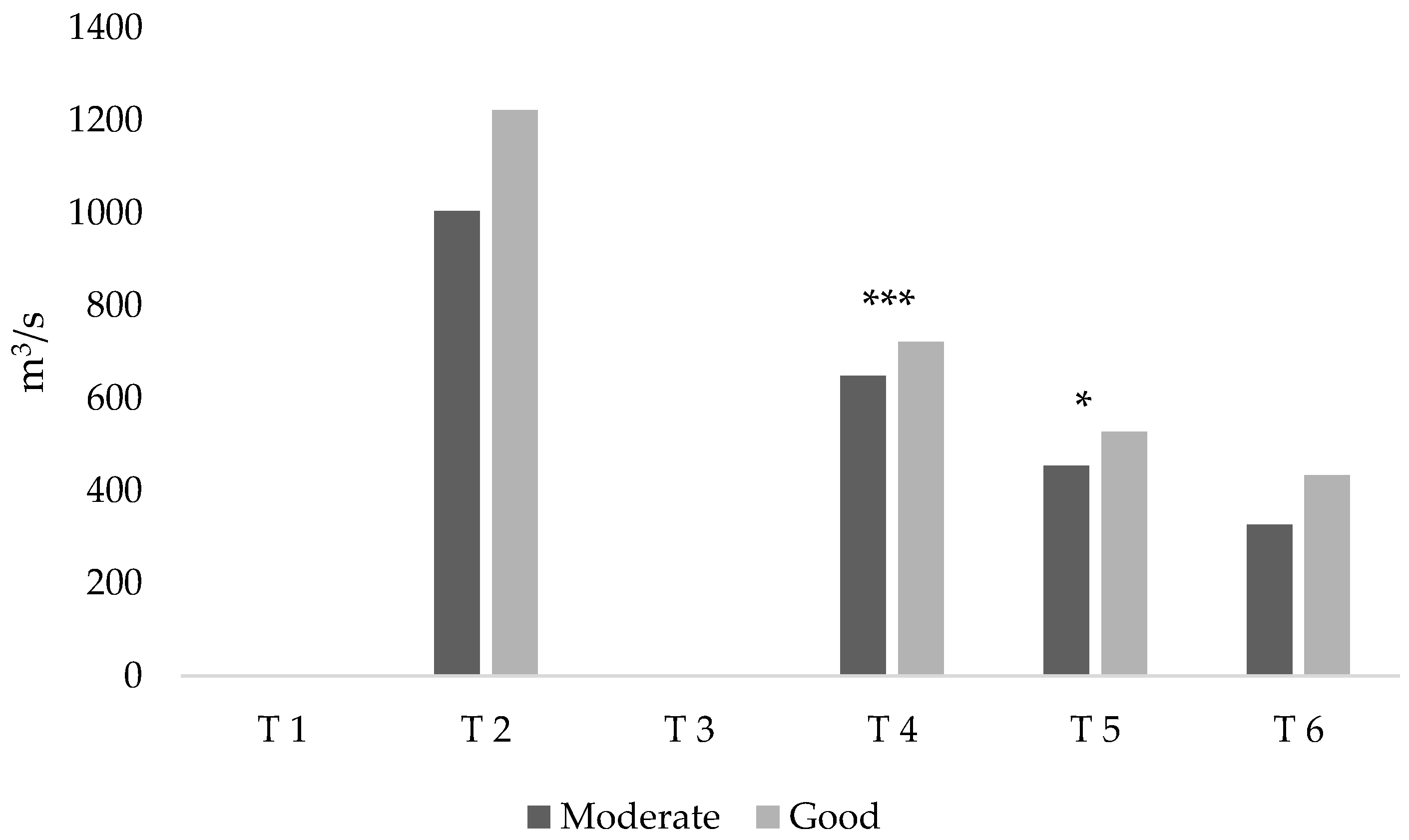
Determine the relationship between mean discharge (instead of the original set of hydrological indices in Belmar et al. [
18]) averaged along the same time periods and ecological status in order to use the obtained models to calculate e-flows to preserve the good ecological status in the lower Ebro (assessing the suitability of the proposals presented up-to-date). Fish communities were characterized using not only ecological quality indices (IBICAT2010, IBICAT2b, and EFI+) but also the ratio of alien species (both in terms of richness and abundance). This allowed determination of the relationship between ecological quality and alien species, in order to validate the 0.5 threshold used by Caiola et al. [
17] to consider a community as impacted.
Introduce habitat and riparian characteristics in the models to determine the influence of habitat complexity.
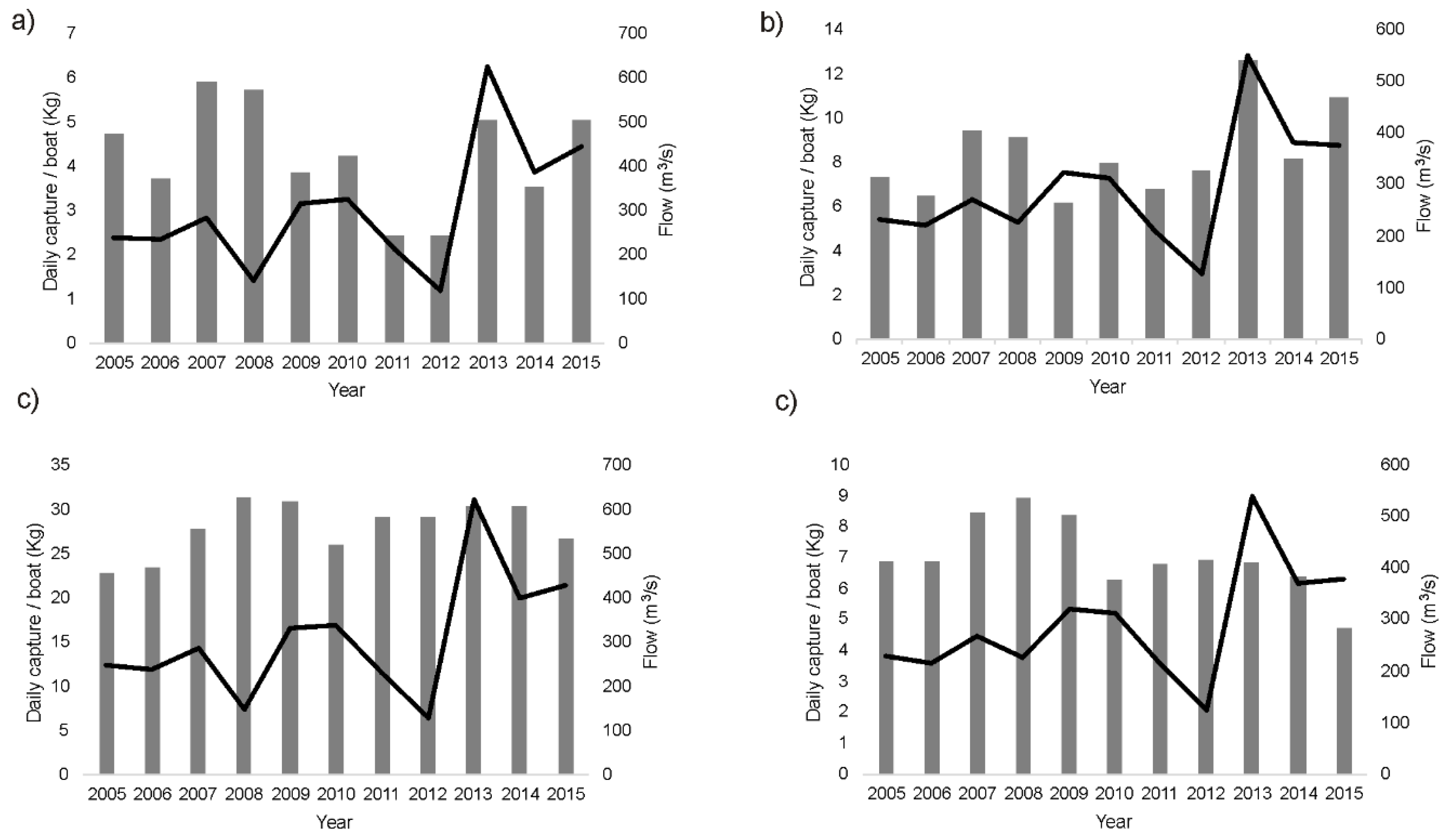
Identify potential relationships among discharge, water quality, and two (shellfish) fishery species with socioeconomic relevance (prawn, or Penaeus kerathurus, and mantis shrimp, or Squilla mantis), as well as between flows and bird populations (ecological relevance) at the delta.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}