Reducing the Phytoplankton Biomass to Promote the Growth of Submerged Macrophytes by Introducing Artificial Aquatic Plants in Shallow Eutrophic Waters


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
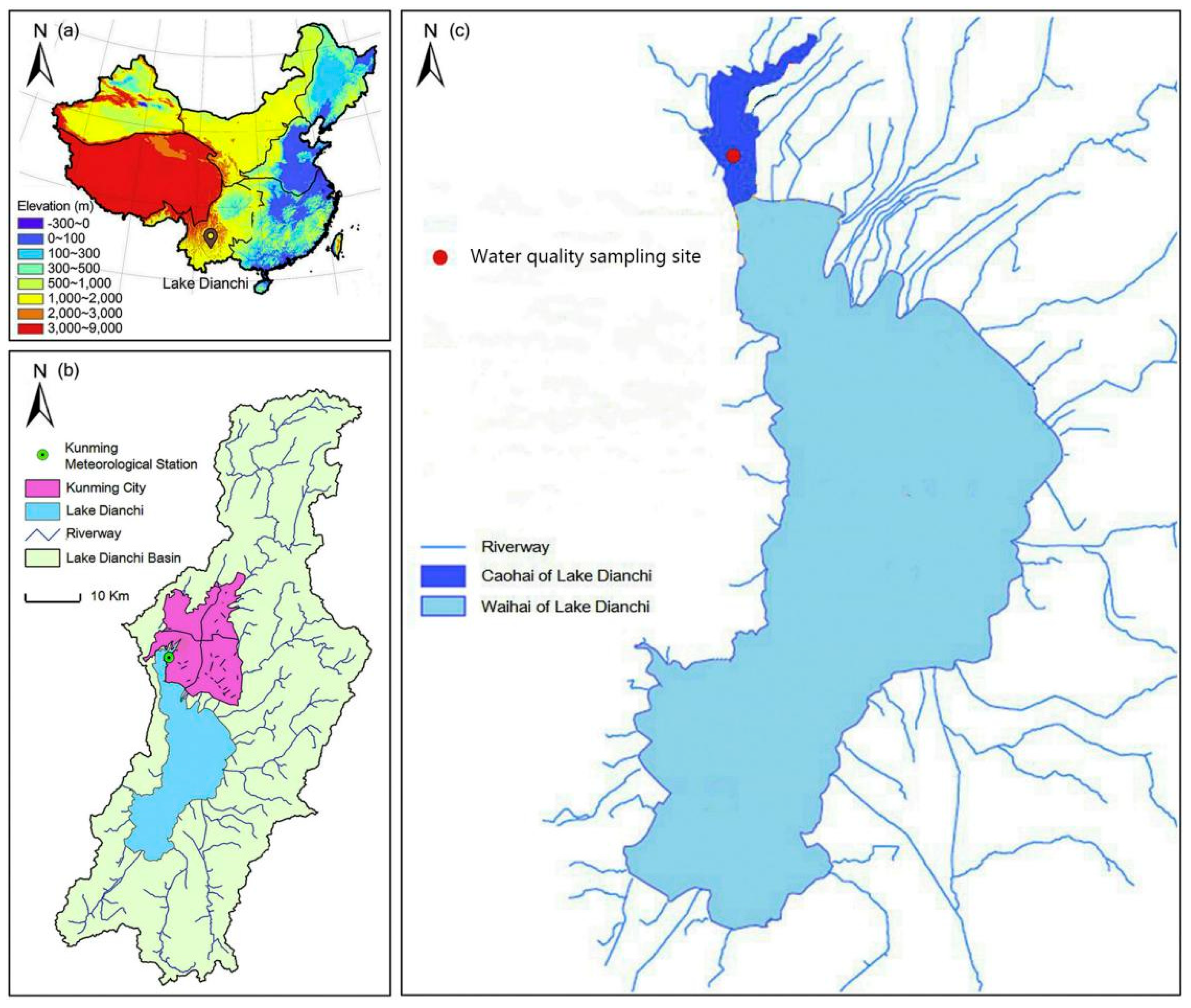
2.1. Study Site
2.2. Experimental Design
2.3. Sample Preparation
2.4. Environmental Parameters
2.5. Phytoplankton Community and Total Phosphorous (TP) Associated with Periphyton Biofilms
2.6. DNA Extraction and Sequencing
2.7. Submerged Macrophyte—Elodea Nuttallii
2.8. Data Analysis
3. Results and Discussion
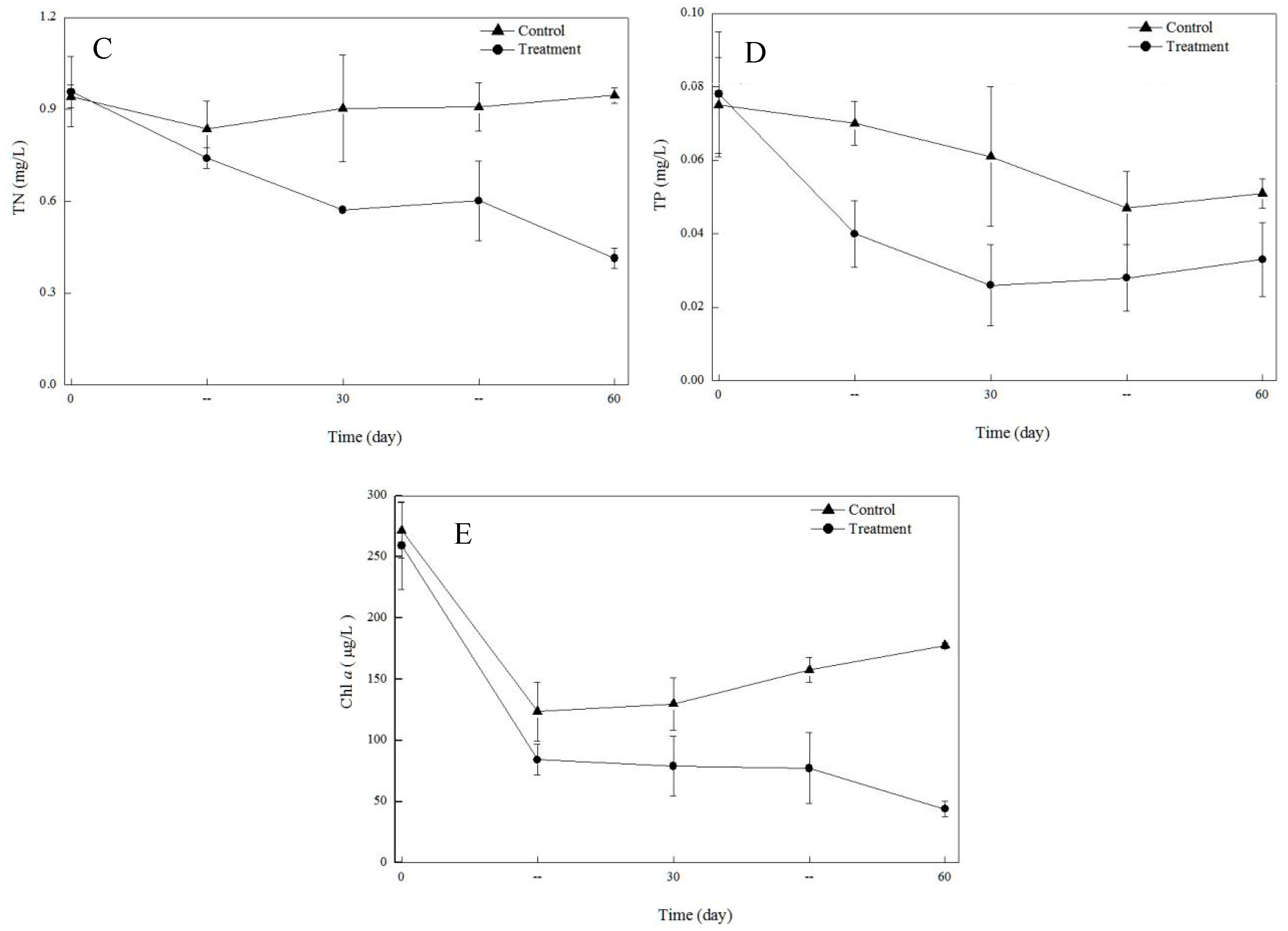
3.1. Variation of Water Quality in the Enclosures
3.2. Variation of TP Concentrations in Periphyton Biofilms Attached to Artificial Aquatic Plants (AAPs)
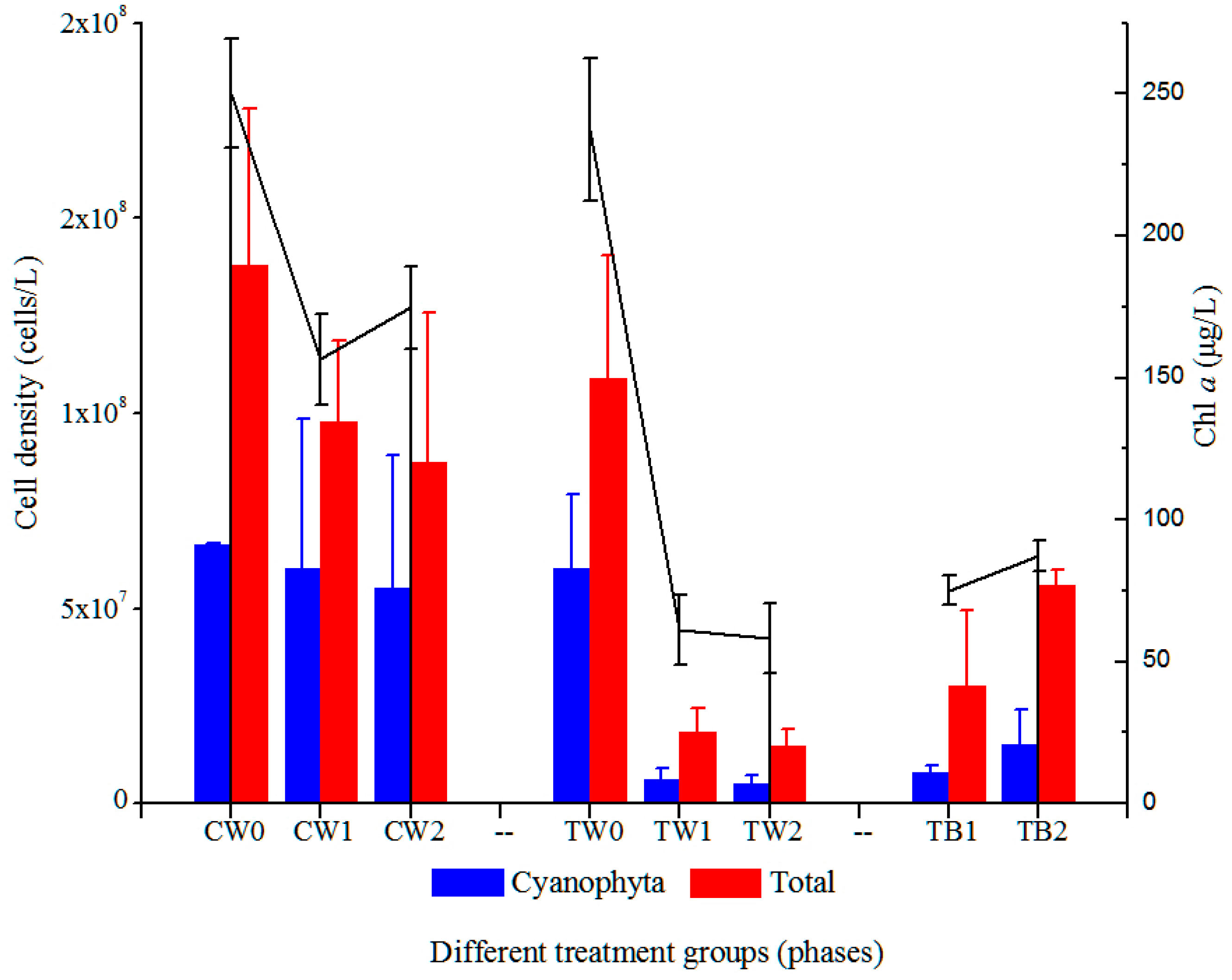
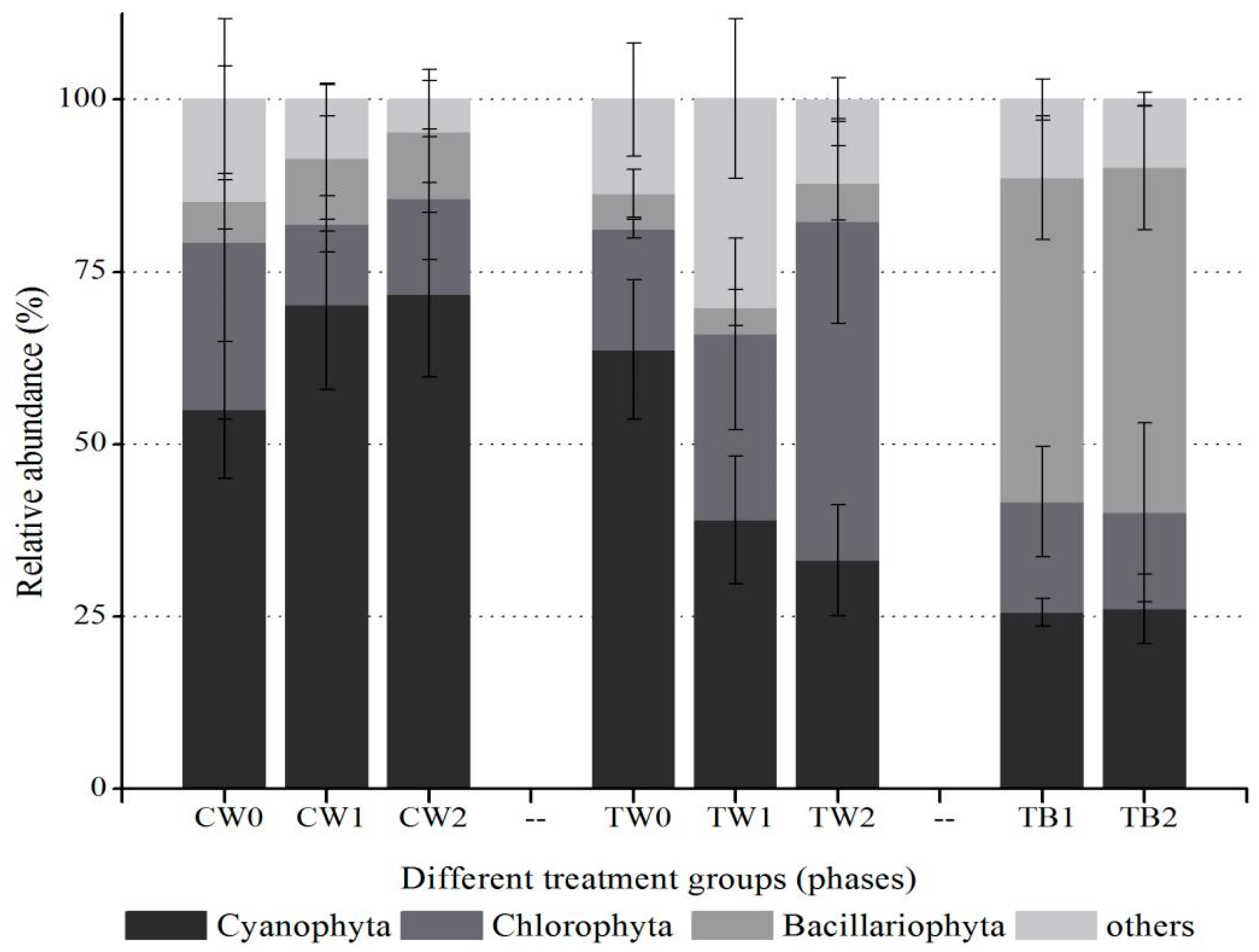
3.3. Dynamics of Phytoplankton in Water Column with the Development of Periphyton Biofilms Attached to AAPs
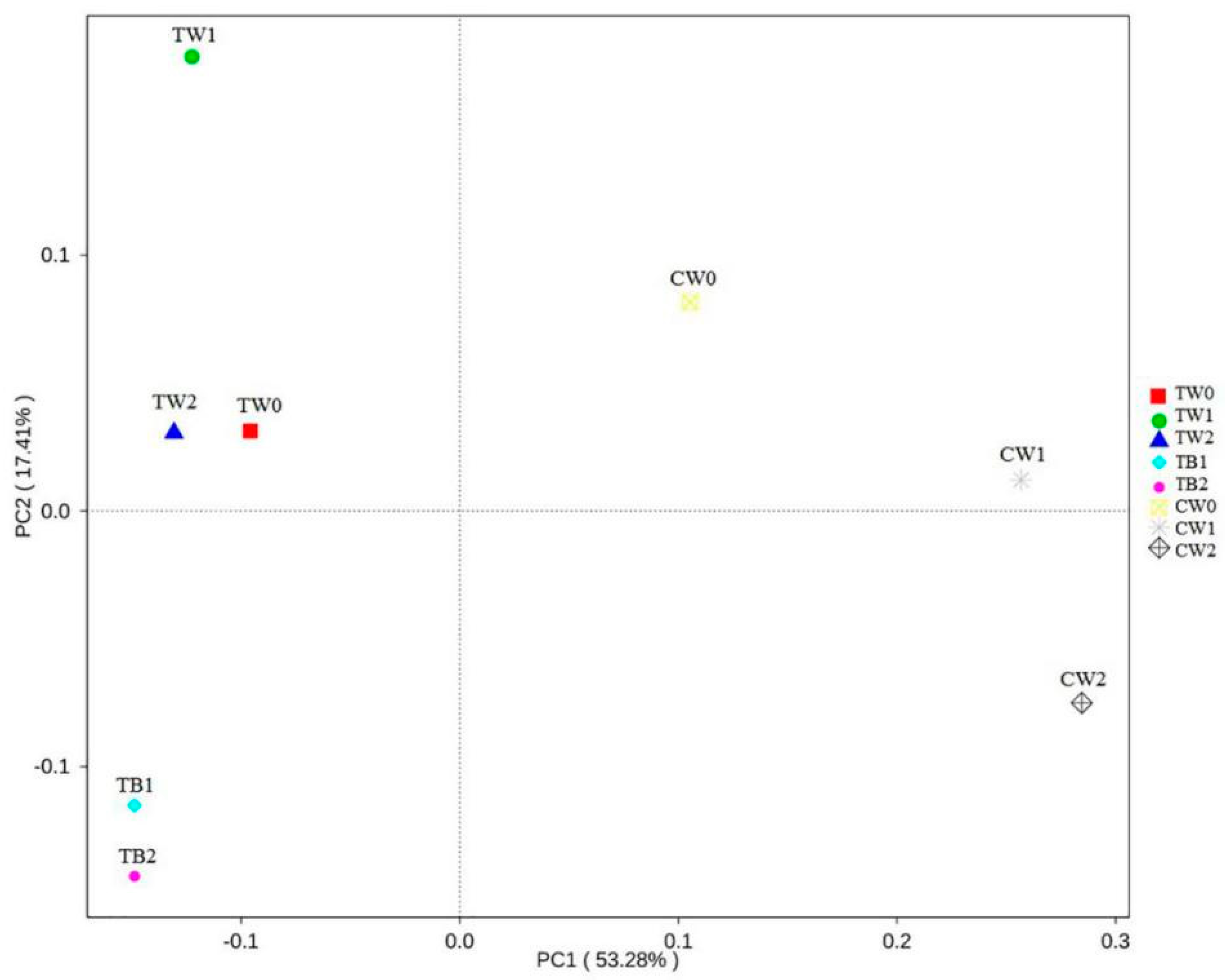
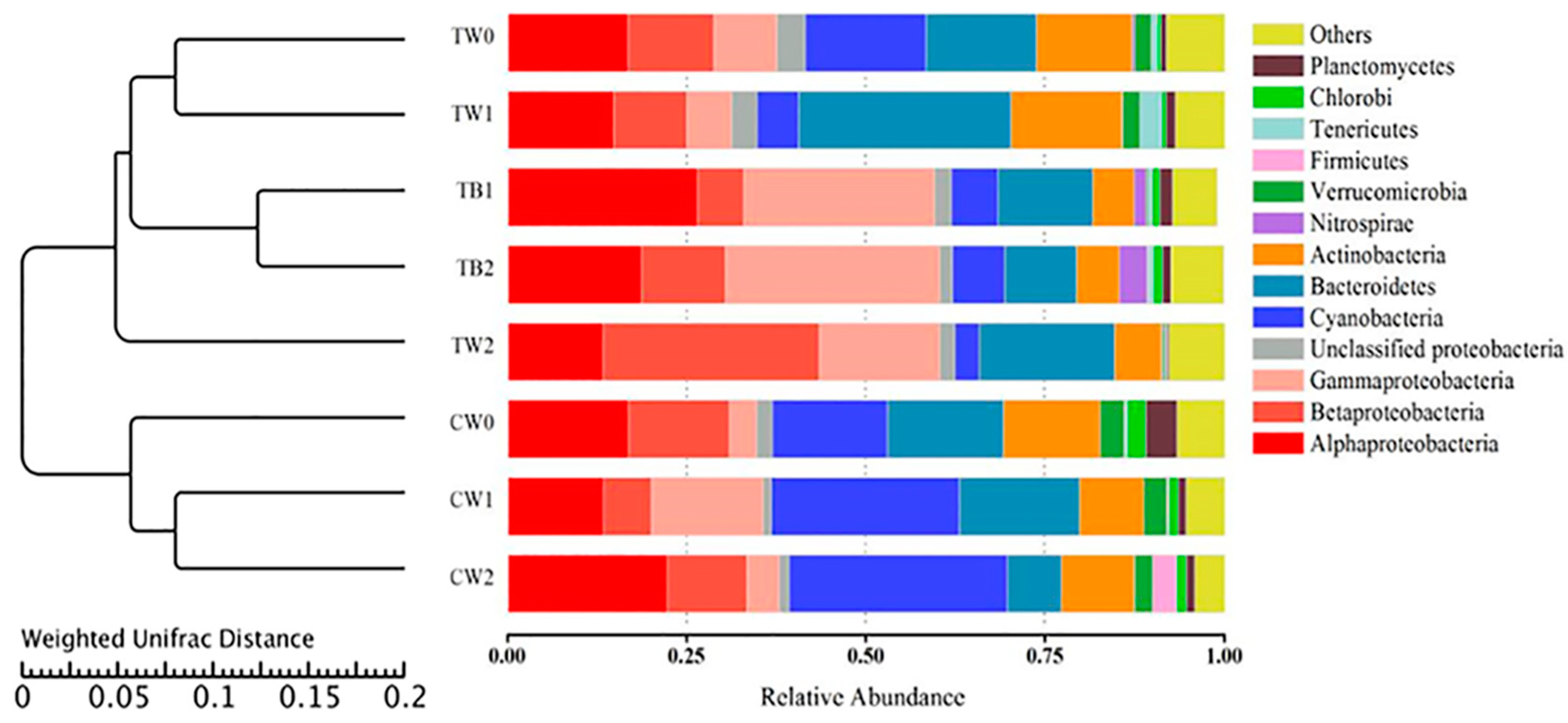
3.4. Dynamics of Bacterial Communities with the Development of Periphyton Biofilms Attached to AAPs
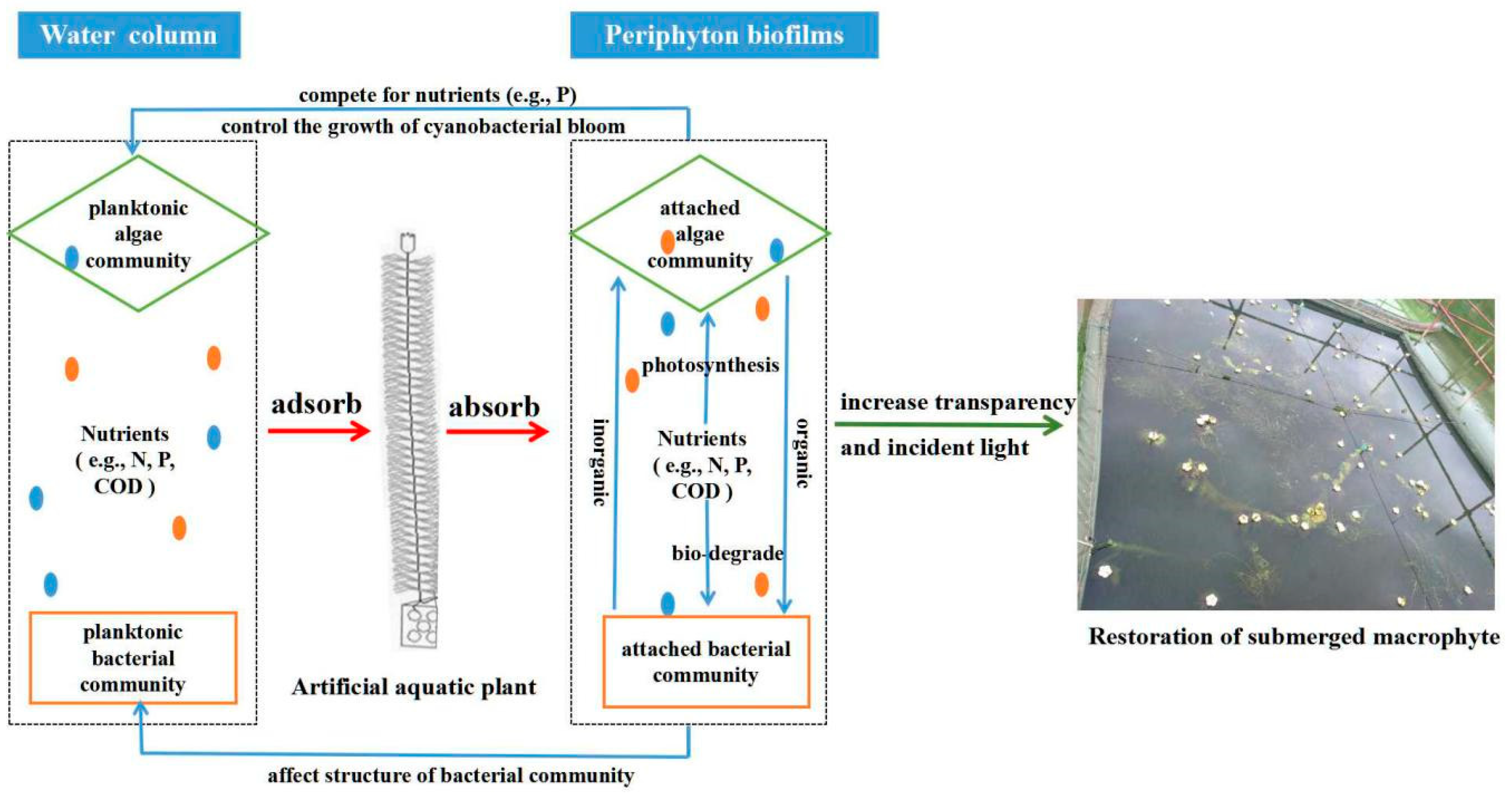
3.5. Restoration of Submerged Macrophytes with Possible Mechanisms
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carpenter, S.R.; Lodge, D.M. Effects of submersed macrophytes on ecosystem processes. Aquat. Bot. 1986, 26, 341–370. [Google Scholar] [CrossRef]
- Maberly, S.C. Ecology of Shallow Lakes; Marten Scheffer, Chapman & Hall: London, UK, 1998; 357p, ISBN 0-412-74920-3. [Google Scholar]
- Gulati, R.D.; Donk, E. Biomanipulation in the Netherlands: Applications in fresh water ecosystems and estuarine water—An introduction. Hydrobiol. Bull. 1989, 23, 1–4. [Google Scholar] [CrossRef]
- Hosper, S.H. Biomanipulation, new perspective for restoring shallow, eutrophic lakes in The Netherlands. Hydrobiol. Bull. 1989, 23, 11–19. [Google Scholar] [CrossRef]
- Scheffer, M.; Hosper, S.H.; Meijer, M.L.; Moss, B.; Jeppesen, E. Alternative equilibria in shallow lakes. Trends Ecol. Evol. 1993, 8, 275–279. [Google Scholar] [CrossRef]
- Waters, M.N.; Schelske, C.L.; Brenner, M. Cyanobacterial dynamics in shallow Lake Apopka (Florida, USA) before and after the shift from a macrophyte-dominated to a phytoplankton-dominated state. Freshwater Biol. 2015, 60, 1571–1580. [Google Scholar] [CrossRef]
- Smith, V.H. Low Nitrogen to phosphorus ratios favor dominance by blue-green algae in lake phytoplankton. Science 1983, 221, 669–671. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. 2009, 1, 27–37. [Google Scholar] [CrossRef]
- Waters, M.N.; Piehler, M.F.; Smoak, J.M.; Martens, C.S. The development and persistence of alternative ecosystem states in a large, shallow lake. Freshwater Biol. 2010, 55, 1249–1261. [Google Scholar] [CrossRef]
- Brock, T.C.M.; Bongaerts, M.C.M.; Heijnen, G.J.M.A.; Heijthuijsen, J.H.F.G. Nitrogen and phosphorus accumulation and cycling by Nymphoides peltata (Gmel.) O. Kuntze (Menyanthaceae). Aquat. Bot. 1983, 17, 189–214. [Google Scholar] [CrossRef]
- Spence, D.H.N. The zonation of plants in freshwater lakes. Adv. Ecol. Res. 1982, 12, 37–125. [Google Scholar]
- Wu, Y.H.; Feng, M.Y.; Liu, J.T.; Zhao, Y. Effects of polyaluminium chloride and copper sulfate on phosphorus and UV254 under different anoxic levels. Fresenius Environ. Bull. 2005, 14, 406–412. [Google Scholar]
- Stüben, D.; Walpersdorf, E.; Voss, K.; Rönick, H.; Schimmele, M.; Baborowski, M.; Luther, G.; Elsner, W. Application of lake marl at lake Arendsee, NE Germany: First results of a geochemical monitoring during the restoration experiment. Sci. Total Environ. 1998, 218, 33–44. [Google Scholar] [CrossRef]
- Hullebusch, E.V.; Deluchat, V.; Chazal, P.M.; Baudu, M. Environmental impact of two successive chemical treatments in a small shallow eutrophied lake: Part I. Case of Aluminium Sulphate. Environ. Pollut. 2002, 120, 617–626. [Google Scholar] [CrossRef]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water. A Guide to their Public Health Consequences, Monitoring and Management; F & FN Spon: London, UK, 1999; p. 416. [Google Scholar]
- Larned, S.T. A prospectus for periphyton: Recent and future ecological research. J. N. Am. Benthol. Soc. 2010, 29, 182–206. [Google Scholar] [CrossRef]
- Underwood, G.J.C.; Perkins, R.G.; Consalvey, M.C.; Hanlon, A.R.M.; Oxborough, K.; Baker, N.R.; Paterson, D.M. Patterns in Microphyto benthic primary productivity: Species-specific variation in migratory rhythms and photosynthetic efficiency in mixed-species biofilms. Limnol. Oceanogr. 2005, 50, 755–767. [Google Scholar] [CrossRef]
- Sukačová, K.; Trtílek, M.; Rataj, T. Phosphorus removal using a microalgal biofilm in a new biofilm photobioreactor for tertiary wastewater treatment. Water Res. 2015, 71, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Liu, X.; Wu, C.; Wu, Y. Nutrient capture and recycling by periphyton attached to modified agrowaste carriers. Environ. Sci. Pollut. Res. 2016, 23, 8035–8043. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, S.; Zhao, H.; Yang, L. Environmentally benign periphyton bioreactors for controlling cyanobacterial growth. Bioresour. Technol. 2010, 101, 9681–9687. [Google Scholar] [CrossRef]
- Wu, Y.; Xia, L.; Yu, Z.; Shabbir, S.; Kerr, P.G. In situ bioremediation of surface waters by periphytons. Bioresour. Technol. 2014, 151, 367–372. [Google Scholar] [CrossRef]
- Huang, L.C. Ecological Restoration of Eutrophic Lake with Artificial Grass—Take Dianchi Lake for Example. Master’s Thesis, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, China, June 2016. [Google Scholar]
- Prieto, D.M.; Rubinos, D.A.; Piñeiro, V.; Díaz-Fierros, F.; Barral, M.T. Influence of epipsammic biofilm on the biogeochemistry of arsenic in freshwater environments. Biogeochemistry 2016, 129, 291–306. [Google Scholar] [CrossRef]
- Teng, Q.X.; Pang, Y.; Hu, X.Z.; Wang, Y.T.; Huang, T.Y. Application of Artificial Plants in Farmland Drainage Ditches of Boluo River. J. Environ. Eng. Technol. 2016, 6, 65–71. [Google Scholar]
- Yang, Y.; Chen, W.; Yi, Z.; Pei, G. The integrative effect of periphyton biofilm and tape grass (Vallisneria natans) on internal loading of shallow eutrophic lakes. Environ. Sci. Pollut. R. 2017, 25, 1773–1783. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Luo, X.; Jin, H.; Gu, J.; Jeppesen, E.; Liu, Z.; Li, K. Effects of exposed artificial substrate on the competition between phytoplankton and benthic algae: Implications for shallow lake restoration. Water 2017, 9, 24. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, W.; Li, D.; Shen, Y.; Li, G.; Liu, Y. First report of aphantoxins in China—Water blooms of toxigenic Aphanizomenon flos-aquae in Lake Dianchi. Ecotoxicol. Environ. Saf. 2006, 65, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Yang, C.; He, L.Q.S.; Dao, G.H.; Du, J.S.; Han, Y.P.; Hu, H.Y. Meteorological factors and water quality changes of Plateau Lake Dianchi in China (1990–2015) and their joint influences on cyanobacterial blooms. Sci. Total Environ. 2019, 665, 406–418. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Du, R.; Zhang, X.; Dong, S.; Sun, S. Succession and seasonal variation in epilithic biofilms on artificial reefs in culture waters of the sea cucumber Apostichopus japonicus. Chin. J. Ocean. Limnol. 2016, 35, 132–152. [Google Scholar] [CrossRef]
- SEPA. Standard Methods for Water and Wastewater Monitoring and Analysis, 4th ed.; State Environmental Protection Administration: Beijing, China, 2002. (In Chinese) [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 1998. [Google Scholar]
- Hu, H.; Wei, Y. The Freshwater Algae of China: Systematics, Taxonomy and Ecology; Science Press: Beijing, China, 2006; 1023p. [Google Scholar]
- Riis, V.; Lorbeer, H.; Babel, W. Extraction of Microorganisms from Soil: Evaluation of the Efficiency by Counting Methods and Activity Measurements. Soil Biol. Biochem. 1998, 30, 1573–1581. [Google Scholar] [CrossRef]
- Nusch, E.A. Comparison of different methods for chlorophyll and phaeopigments determination. Arch. Hydrobiol. Beih. Ergebn. Limnol. 1980, 14, 14–36. [Google Scholar]
- Rao, C.R.M.; Reddi, G.S. Decomposition procedure with aqua regia and hydrofluoric acid at room temperature for the spectrophotometric determination of phosphorus in rocks and minerals. Analyt. Chim. Acta 1990, 237, 251–252. [Google Scholar] [CrossRef]
- Kunii, H. Characteristics of the winter growth of detached Elodea nuttallii (planch.) St John in Japan. Aquat. Bot. 1981, 11, 57–66. [Google Scholar] [CrossRef]
- Kunii, H. Seasonal growth and profile structure development of Elodea nuttallii in pond Ojaga-Ike, Japan (Planch.) St John. Aquat. Bot. 1984, 18, 239–247. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Knight, R. QIIME allows analysis of high-throughput community sequencing data. Nat. Med. 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.L.; Zhou, L.; Li, S.M.; Fang, G.F. Nutrient removal and microbial community structure in hybrid biofilm reactor. Chin. J. Ophthalmol. 2012, 48, 24–27. [Google Scholar]
- Sukačová, K.; Kočí, R.; Žídková, M.; Vítěz, T.; Trtílek, M. Novel insight into the process of nutrients removal using an algal biofilm: The evaluation of mechanism and efficiency. Int. J. Phytoremediat. 2017, 19, 909–914. [Google Scholar] [CrossRef] [PubMed]
- Romaní, A.M.; Giorgi, A.; Acuña, V.; Sabater, S. The influence of substratum type and nutrient supply on biofilm organic matter utilization in streams. Limnol. Oceanogr. 2004, 49, 1713–1721. [Google Scholar] [CrossRef] [Green Version]
- McCormick, P.V.; Shuford, R.B.E.; Chimney, M.J. Periphyton as a potential phosphorus sink in the Everglades Nutrient Removal Project. Ecol. Eng. 2006, 27, 279–289. [Google Scholar] [CrossRef]
- Chiou, R.J.; Yang, Y.R. An evaluation of the phosphorus storage capacity of an anaerobic/aerobic sequential batch biofilm reactor. Bioresour. Technol. 2008, 99, 4408–4413. [Google Scholar] [CrossRef] [PubMed]
- Hickman, J.W.; Tifrea, D.F.; Harwood, C.S. A chemosensory system that regulates biofilm formation through modulation of cyclic diguanylate levels. Proc. Nat. Acad. Sci. USA 2005, 102, 14422–14427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Wang, C.; Qin, H.; Li, Y.; Zheng, J.; Peng, C.; Li, D. Influence of phosphorus availability on the community structure and physiology of cultured biofilms. J. Environ. Sci. 2016, 42, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Spencer, D.F.; Linquist, B.A. Reducing rice field algae and cyanobacteria abundance by altering phosphorus fertilizer applications. Paddy Water Environ. 2013, 12, 147–154. [Google Scholar] [CrossRef]
- Tlili, A.; Hollender, J.; Kienle, C.; Behra, R. Micropollutant-induced tolerance of in situ periphyton: Establishing causality in wastewater-impacted streams. Water Res. 2017, 111, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Artigas, J.; Fund, K.; Kirchen, S.; Morin, S.; Obst, U.; Romaní, A.M.; Schwartz, T. Patterns of biofilm formation in two streams from different bioclimatic regions: Analysis of microbial community structure and metabolism. Hydrobiologia 2012, 695, 83–96. [Google Scholar] [CrossRef]
- Joint, I.; Tait, K.; Wheeler, G. Cross-kingdom signalling: Exploitation of bacterial quorum sensing molecules by the green seaweed Ulva. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2007, 362, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Carr, G.M.; Morin, A.; Chambers, P.A. Bacteria and algae in stream periphyton along a nutrient gradient. Freshwater Biol. 2005, 50, 1337–1350. [Google Scholar] [CrossRef]
- Suberkropp, K.; Gulis, V.; Rosemond, A.D.; Benstead, J.P. Ecosystem and physiological scales of microbial responses to nutrients in a detritus-based stream: Results of a 5-year continuous enrichment. Limnol. Oceanogr. 2009, 55, 149–160. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.Y.; Xu, Y.; Liu, Y.; Liu, Y.; Liu, Z.P. Bacterial diversity, community structure and function associated with biofilm development in a biological aerated filter in a recirculating marine aquaculture system. Mar. Biol. 2011, 42, 1–11. [Google Scholar] [CrossRef]
- Atabek, A.; Camesano, T.A. Atomic force microscopy study of the effect of lipopolysaccharides and extracellular polymers on adhesion of Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 8503–8509. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.W.; Wang, X.L.; Xing, M. Dynamic variation of microbial metabolites and community involved in membrane fouling in A/O-MBR. J. Memb. Sci. 2014, 458, 157–163. [Google Scholar] [CrossRef]
- Martiny, A.C.; Albrechtsen, H.J.; Arvin, E.; Molin, S. Identification of bacteria in biofilm and bulk water samples from a nonchlorinated model drinking water distribution system: Detection of a large nitrite-oxidizing population associated with Nitrospira spp. Appl. Environ. Microbiol. 2005, 71, 8611–8617. [Google Scholar] [CrossRef]
- Ren, J.C.; Zhou, H.; Sun, Y.T. Vertical distribution of light intensity and light compensation depth of submerged macrophyte in Lake Dianchi. Acta Scicentiarum Nat. Univ. Pekin. 1997, 2, 211–214. (In Chinese) [Google Scholar]
- Feldmann, T.; Nõges, P. Factors controlling macrophyte distribution in large shallow Lake Võrtsjärv. Aquat. Bot. 2007, 87, 15–21. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Sequence Number | OTU Number | Shannon Index | Simpson Index | Chao Index | ACE Index | Goods-Coverage |
|---|---|---|---|---|---|---|---|
| CW0 | 50,000 | 1764 | 7.774 | 0.984 | 1701.511 | 1745.288 | 0.994 |
| CW1 | 50,874 | 1445 | 6.370 | 0.925 | 1398.249 | 1420.827 | 0.995 |
| CW2 | 44,922 | 1469 | 6.354 | 0.92 | 1512.567 | 2332.759 | 0.987 |
| TW0 | 46,587 | 1672 | 6.764 | 0.969 | 1599.339 | 1649.734 | 0.992 |
| TW1 | 61,178 | 2214 | 8.336 | 0.991 | 2174.538 | 2204.640 | 0.993 |
| TW2 | 54,552 | 2014 | 8.273 | 0.992 | 1913.526 | 1992.917 | 0.993 |
| TB1 | 52,842 | 2056 | 7.856 | 0.968 | 2009.426 | 2033.116 | 0.993 |
| TB2 | 49,478 | 2099 | 8.377 | 0.984 | 2087.237 | 2097.015 | 0.993 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Huang, L.; Wang, Y.; Li, L.; Li, G.; Xiao, B.; Song, L. Reducing the Phytoplankton Biomass to Promote the Growth of Submerged Macrophytes by Introducing Artificial Aquatic Plants in Shallow Eutrophic Waters. Water 2019, 11, 1370. https://doi.org/10.3390/w11071370
Wu Y, Huang L, Wang Y, Li L, Li G, Xiao B, Song L. Reducing the Phytoplankton Biomass to Promote the Growth of Submerged Macrophytes by Introducing Artificial Aquatic Plants in Shallow Eutrophic Waters. Water. 2019; 11(7):1370. https://doi.org/10.3390/w11071370
Chicago/Turabian StyleWu, Yue, Licheng Huang, Yalin Wang, Lin Li, Genbao Li, Bangding Xiao, and Lirong Song. 2019. "Reducing the Phytoplankton Biomass to Promote the Growth of Submerged Macrophytes by Introducing Artificial Aquatic Plants in Shallow Eutrophic Waters" Water 11, no. 7: 1370. https://doi.org/10.3390/w11071370
APA StyleWu, Y., Huang, L., Wang, Y., Li, L., Li, G., Xiao, B., & Song, L. (2019). Reducing the Phytoplankton Biomass to Promote the Growth of Submerged Macrophytes by Introducing Artificial Aquatic Plants in Shallow Eutrophic Waters. Water, 11(7), 1370. https://doi.org/10.3390/w11071370