Mini-Hydrocyclone Separation of Cyanobacterial and Green Algae: Impact on Cell Viability and Chlorine Consumption

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
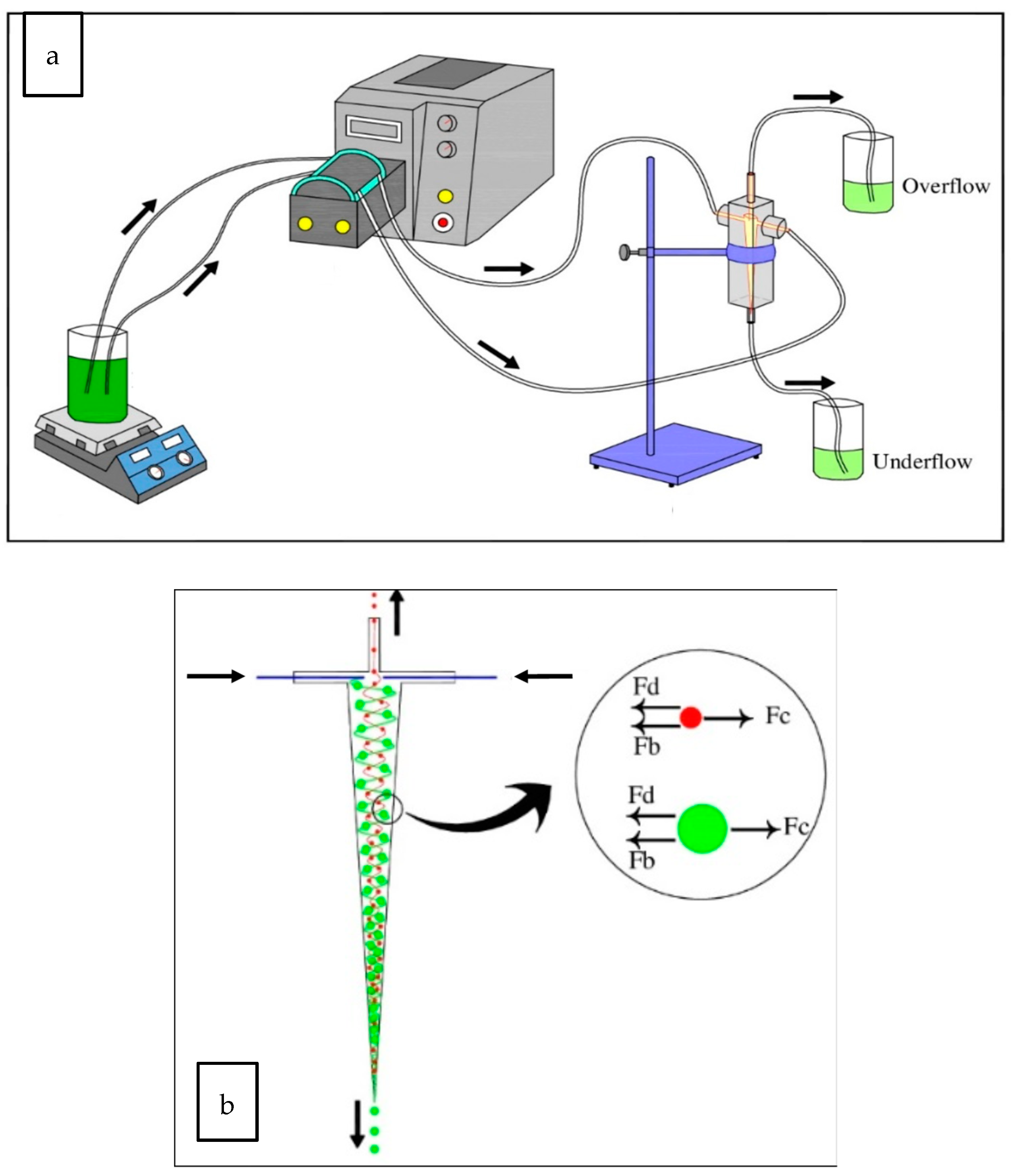
2.1. Mini-Hydrocyclone Fabrication
2.2. Preparation of the Phytoplankton Mixture and Chlorination
2.3. Cell Integrity and Counting Methods
2.4. Theoretical Separation Calculations
3. Results and Discussion
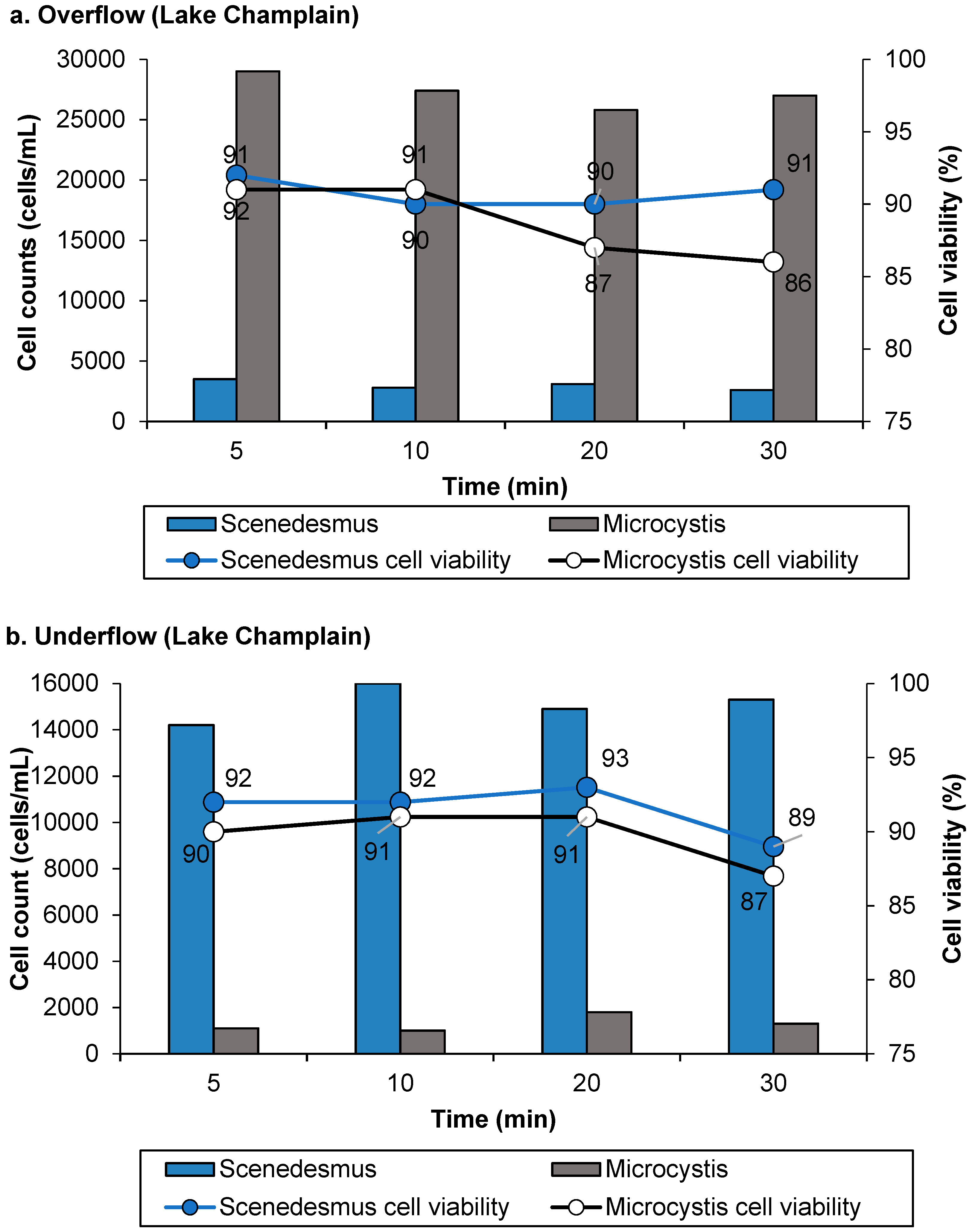
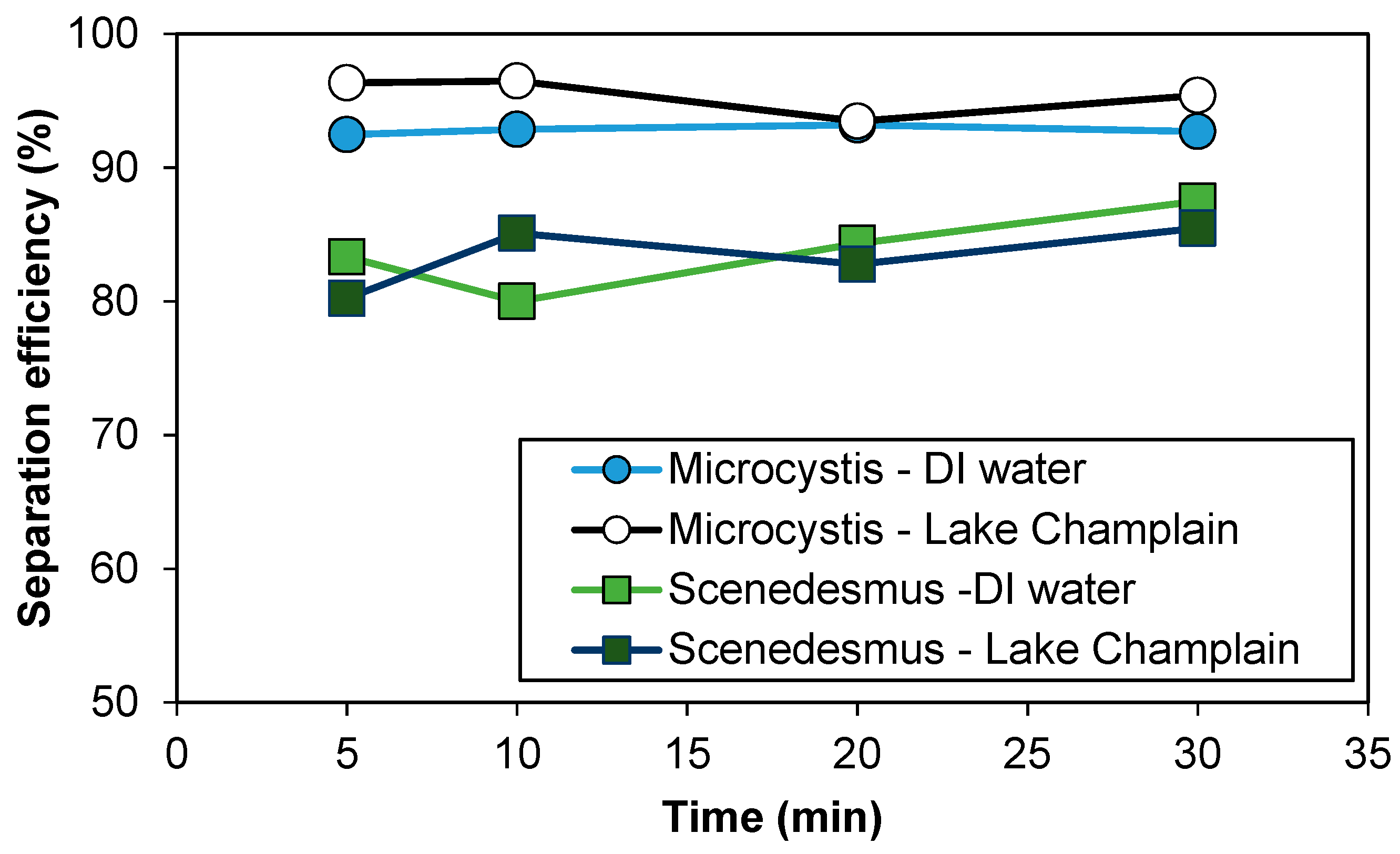
3.1. Separation Efficiency of Mini-Hydrocyclone
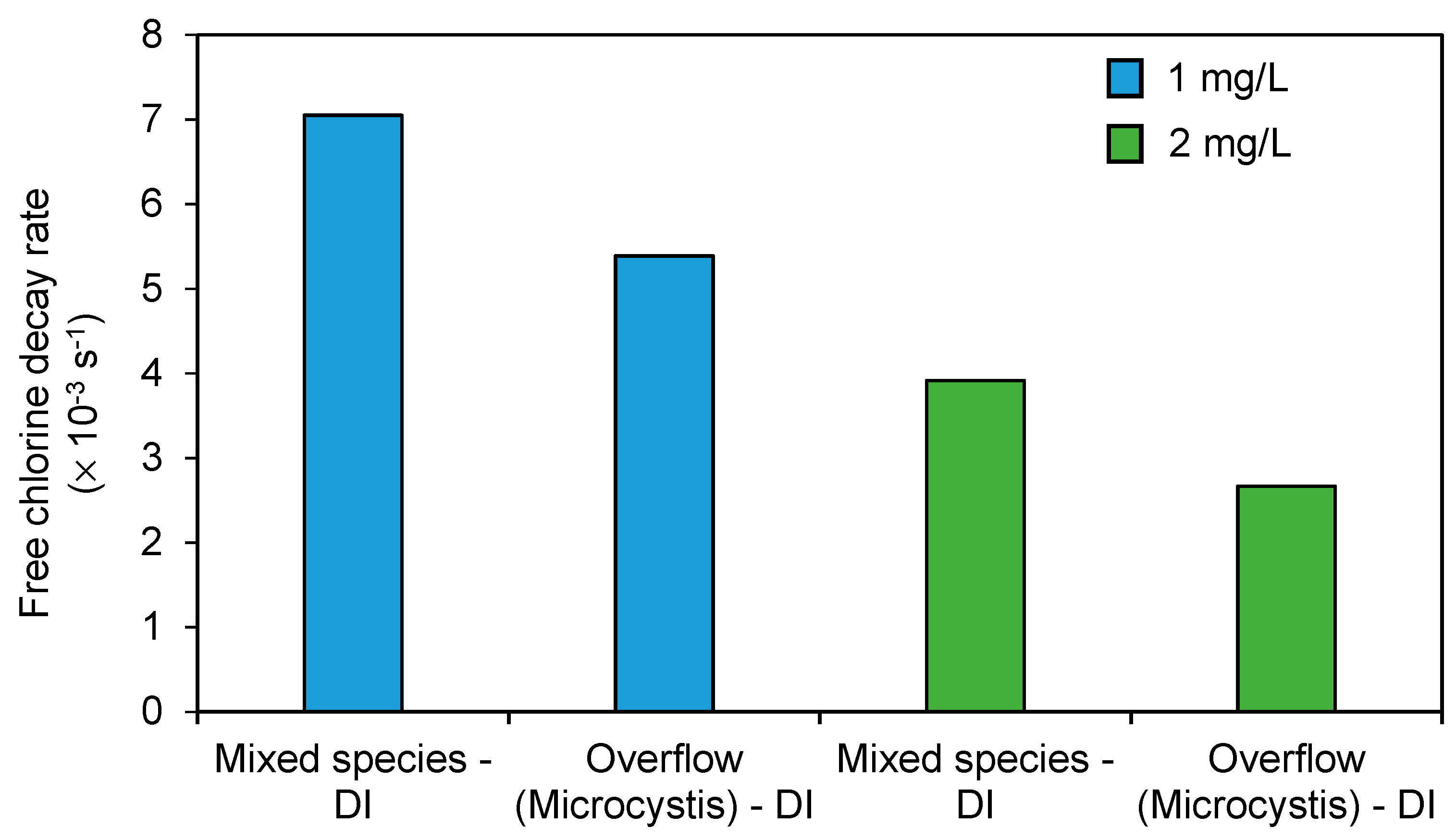
3.2. Impact of Separation of Chlorine Consumption
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tomlinson, A.; Drikas, M.; Brookes, J.D. The role of phytoplankton as pre-cursors for disinfection by-product formation upon chlorination. Water Res. 2016, 102, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Linden, L.G.; Lewis, D.M.; Burch, M.D.; Brookes, J.D. Interannual variability in rainfall and its impact on nutrient load and phytoplankton in Myponga Reservoir, South Australia. Int. J. River Basin Manag. 2004, 2, 169–179. [Google Scholar] [CrossRef]
- Bade, D.L.; Carpenter, S.R.; Cole, J.J.; Pace, M.L.; Kritzberg, E.; Van de Bogert, M.C.; Cory, R.M.; McKnight, D.M. Sources and fates of dissolved organic carbon in lakes as determined by whole-lake carbon isotope additions. Biogeochemistry 2007, 84, 115–129. [Google Scholar] [CrossRef]
- Paerl, H.W. Mitigating Toxic Planktonic Cyanobacterial Blooms in Aquatic Ecosystems Facing Increasing Anthropogenic and Climatic Pressures. Toxins 2018, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Zamyadi, A.; Dorner, S.; Sauvé, S.; Ellis, D.; Bolduc, A.; Bastien, C.; Prévost, M. Species-dependence of cyanobacteria removal efficiency by different drinking water treatment processes. Water Res. 2013, 47, 1080–1090. [Google Scholar] [CrossRef] [PubMed]
- Zamyadi, A.; Romanis, C.; Mills, T.; Neilan, B.; Choo, F.; Coral, L.A.; Gale, D.; Newcombe, G.; Crosbie, N.; Stuetz, R.; et al. Diagnosing water treatment critical control points for cyanobacterial removal: Exploring benefits of combined microscopy, next-generation sequencing, and cell integrity methods. Water Res. 2019, 152, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Pazouki, P.; Prevost, M.; McQuiad, N.; Barbeau, B.; de Boutray, M.-L.; Zamyadi, A.; Dorner, S. Breakthrough of cyanobacteria in bank filtration. Water Res. 2016, 102, 170–179. [Google Scholar] [CrossRef]
- Zamyadi, A.; Coral, L.A.; Barbeau, B.; Dorner, S.; Lapolli, F.R.; Prévost, M. Fate of toxic cyanobacterial genera from natural bloom events during ozonation. Water Res. 2015, 73, 204–215. [Google Scholar] [CrossRef]
- Zamyadi, A.; Henderson, R.K.; Stuetz, R.; Newcombe, G.; Newton, K.; Gladman, B. Cyanobacterial management in full-scale water treatment and recycling processes: Reactive dosing following intensive monitoring. Environ. Sci. Water Res. Technol. 2016, 2, 362–375. [Google Scholar] [CrossRef]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; World Health Organization (WHO): London, UK, 1999. [Google Scholar]
- Holtcamp, W. The emerging science of BMAA: Do cyanobacteria contribute to neurodegenerative disease? Environ. Health Perspect. 2012, 120, a110–a116. [Google Scholar] [CrossRef]
- Bradley, W.G.; Borenstein, A.R.; Nelson, M.; Codd, G.A.; Rosen, B.H.; Stommel, E.W.; Cox, P.A. Is exposure to cyanobacteria an environmental risk factor for amyotrophic lateral sclerosis and other neurodegenerative disease? Amyotroph. Lateral Scler. Frontotemporal Degener. 2013, 14, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Chernoff, N.; Hill, D.J.; Diggs, D.L.; Faison, B.D.; Francis, B.M.; Lang, J.R.; Larue, M.M.; Le, T.-T.; Loftin, K.A.; Lugo, J.N.; et al. A critical review of the postulated role of the nonessential amino acid, b-N-methylamino-L-alanine, in neurodegenerative disease in humans. J. Toxicol. Environ. Health 2017, 20, 183–229. [Google Scholar] [CrossRef] [PubMed]
- Suffet, I.H.; Corado, A.; Chou, D.; McGuire, M.J.; Butterworth, S. AWWA taste and odor survey. JAWWA 1996, 88, 168–180. [Google Scholar] [CrossRef]
- Watson, S.B.; Ridal, J.; Boyer, G.L. Taste and odour and cyanobacterial toxins: Impairment, prediction, and management in the Great Lakes. Can. J. Fish. Aquat. Sci. 2008, 65, 1779–1796. [Google Scholar] [CrossRef]
- Satchwill, T.; Watson, S.B.; Dixon, E. Odourous algal-derived alkenes: Differences in stability and treatment responses in drinking water. Water Sci. Technol. 2007, 55, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Jüttner, F.; Watson, S.B. Biochemical and ecological control of geosmin and 2-methylisoborneol in source waters. Appl. Environ. Microbiol. 2007, 73, 4395–4406. [Google Scholar] [CrossRef] [PubMed]
- Zamyadi, A.; Fan, Y.; Daly, R.I.; Prevost, M. Chlorination of microcystis aeruginosa: Toxin release and oxidation, cellular chlorine demand and disinfection by-products formation. Water Res. 2013, 47, 1080–1090. [Google Scholar] [CrossRef]
- Syed, M.S.; Rafeie, M.; Henderson, R.; Vandamme, D.; Asadnia, M.; Ebrahimi Warkiani, M. A 3D-printed mini-hydrocyclone for high throughput particle separation: Application to primary harvesting of microalgae. Lab Chip 2017, 17, 2459–2469. [Google Scholar] [CrossRef]
- Maier, R.; Pepper, I.L.; Gerba, C.P. Environmental Microbiology, 2nd ed.; Academic Press, Elsevier: Cambridge, MA, USA, 2009. [Google Scholar]
- Kommineni, S.; Amante, K.; Karnik, B.; Sommerfeld, M.; Dempster, T. Strategies for Controlling and Mitigating Algal Growth within Water Treatment Plants; Water Research Foundation: Denver, CO, USA, 2009. [Google Scholar]
- American Public Health Association (APHA). Standard Methods for the Examination of Water and Wastewater; American Water Works Association, Water Environment Federation: Washington, DC, USA, 2012. [Google Scholar]
- Lund, J.W.G.; Kipling, C.; Le Cren, E.D. The inverted microscope method of estimating algal number and the statistical basis of estimations by counting. Hydrobiologia 1958, 11, 143–170. [Google Scholar] [CrossRef]
- Henderson, R.; Parson, S.A.; Jefferson, B. The impact of algal properties and pre-oxidation on solid-liquid separation of algae. Water Res. 2008, 42, 1827–1845. [Google Scholar] [CrossRef]
- Walsby, A.E. Gas vesicles. Microbiol. Rev. 1994, 58, 94–144. [Google Scholar] [CrossRef] [PubMed]
- Oliver, R. Floating and sinking in gas-vacuolate cyanobacteria. J. Phycol. 1994, 30, 161–173. [Google Scholar] [CrossRef]
- Li, M.; Zhu, W.; Guo, L.; Hu, J.; Chen, H.; Xiao, M. To increase size or decrease density? Different Microcystis species has different choice to form blooms. Sci. Rep. 2016, 6, 37056. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.S.; Oliver, R.L.; Walsby, A.E. Cyanobacterial dominance: The role of buoyancy regulation in dynamic lake environments. N. Z. J. Mar. Freshw. Res. 1987, 21, 379–390. [Google Scholar] [CrossRef]
- Simioni, T.; Quadri, M.B.; Derner, R.B. Drying of Scenedesmus obliquus: Experimental and modeling study. Algal Res. 2019, 39, 101428. [Google Scholar] [CrossRef]
- Lavoie, A.; Mouget, J.-L.; de la Noüe, J. Measurement of freshwater micro-algae cell density with Percoll density gradients. J. Microbiol. Methods 1986, 4, 251–259. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, L.-L.; Li, J.; Chen, M.; Zhang, Y.-W. Mechanism of the influence of hydrodynamics on Microcystis aeruginosa, a dominant bloom species in reservoirs. Sci. Total Environ. 2018, 636, 230–239. [Google Scholar] [CrossRef]
- Wang, C.; Lan, C.Q. Effects of shear stress on microalgae—A review. Biotechnol. Adv. 2018, 36, 986–1002. [Google Scholar] [CrossRef]
- Matthijs, H.C.P.; Visser, P.M.; Reeze, B.; Meeuse, J.; Slot, P.C.; Wijn, G.; Talens, R.; Huisman, J. Selective suppression of harmful cyanobacteria in an entire lake with hydrogen peroxide. Water Res. 2012, 46, 1460–1472. [Google Scholar] [CrossRef]
- Fan, J.J.; Ho, L.; Hobson, P.; Brookes, J. Evaluating the effectiveness of copper sulfate, chlorine, potassium permanganate, hydrogen peroxide, and ozone on cyanobacterial cell integrity. Water Res. 2013, 47, 5153–5164. [Google Scholar] [CrossRef]
- Xiao, M.; Li, M.; Reynolds, C.S. Colony formation in the cyanobacterium Microcystis. Biol. Rev. 2018, 93, 1399–1420. [Google Scholar] [CrossRef] [PubMed]
- Daly, R.I.; Ho, L.; Brookes, J.D. Effect of chlorination on Microcystis aeruginosa cell integrity and subsequent microcystin release and degradation. Environ. Sci. Technol. 2007, 41, 4447–4453. [Google Scholar] [CrossRef] [PubMed]
- Zamyadi, A.; MacLeod, S.L.; Fan, Y.; McQuaid, N.; Dorner, S.; Sauvé, S.; Prévost, M. Toxic cyanobacterial breakthrough and accumulation in a drinking water treatment plant: A monitoring and treatment challenge. Water Res. 2012, 46, 1511–1523. [Google Scholar] [CrossRef] [PubMed]
- Zamyadi, A.; Ho, L.; Newcombe, G.; Daly, R.I.; Burch, M.; Baker, P.; Prévost, M. Release and oxidation of cell-bound saxitoxins during chlorination of Anabaena circinalis cells. Environ. Sci. Technol. 2010, 44, 9055–9061. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type | Cell Diameter (μm) | Density (kg m−3) | Estimated Total Force | Overflow or Underflow |
|---|---|---|---|---|
| Microcystis aeruginosa | 5 | 985 | −1250 | Overflow |
| 5 | 1005 | 1250 | ||
| Scenedesmus obliquus | 10 | 1070 | 75,000 | Underflow |
| 10 | 1310 | 315,000 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moradinejad, S.; Vandamme, D.; Glover, C.M.; Seighalani, T.Z.; Zamyadi, A. Mini-Hydrocyclone Separation of Cyanobacterial and Green Algae: Impact on Cell Viability and Chlorine Consumption. Water 2019, 11, 1473. https://doi.org/10.3390/w11071473
Moradinejad S, Vandamme D, Glover CM, Seighalani TZ, Zamyadi A. Mini-Hydrocyclone Separation of Cyanobacterial and Green Algae: Impact on Cell Viability and Chlorine Consumption. Water. 2019; 11(7):1473. https://doi.org/10.3390/w11071473
Chicago/Turabian StyleMoradinejad, Saber, Dries Vandamme, Caitlin M. Glover, Tahere Zadfathollah Seighalani, and Arash Zamyadi. 2019. "Mini-Hydrocyclone Separation of Cyanobacterial and Green Algae: Impact on Cell Viability and Chlorine Consumption" Water 11, no. 7: 1473. https://doi.org/10.3390/w11071473
APA StyleMoradinejad, S., Vandamme, D., Glover, C. M., Seighalani, T. Z., & Zamyadi, A. (2019). Mini-Hydrocyclone Separation of Cyanobacterial and Green Algae: Impact on Cell Viability and Chlorine Consumption. Water, 11(7), 1473. https://doi.org/10.3390/w11071473