1. Introduction
Coastal lagoons are typically human-managed, highly productive ecosystems which provide multiple uses and services [
1,
2,
3]. However, in the last few decades, engineering interventions have increasingly affected lagoons’ morphology, hydrodynamics, and sedimentology, resulting in excessive inputs of nutrients and organic matter (OM) that have led to eutrophication and organic over-enrichment of the sediments [
4,
5,
6,
7]. These factors have profoundly impaired the biological structure, trophic status, and functioning of coastal lagoon systems worldwide [
8,
9,
10,
11]. As an example, dystrophic events are the cause of anoxia, sulfide development, mass mortalities, and shifts to alternative ecohydrological states characterized by reducing conditions and lower pH [
12,
13,
14,
15]. Within this context, the concept of “saprobity”, originally developed for rivers and lakes more than a century ago [
16,
17], applies well to coastal lagoons as a “state descriptor” of the ecosystem’s condition resulting from the input and decomposition of OM and the removal of its catabolites [
18]. Thus, both the trophic features (i.e., the amount of nutrients and OM in water and sediments) and the degree of saprobity (i.e., the balance between input of OM and other processes such as mineralization, sinking, dilution and export of OM) are instrumental for assessing the natural conditions and the environmental quality of a lagoon. Contrary to estuaries where the salinity gradient is dominant [
19], the seawater renewal and hydrodynamics strongly govern the land-sea gradient in coastal lagoons [
18,
20]. Several field studies have described how species or groups of benthic organisms reduce their abundance in a given order as environmental stress increases, and how the number of taxonomic groups is reduced as stress increases [
21,
22,
23,
24,
25]. Similarly, the main effect of saprobity on the macrozoobenthic assemblages is that the number of species that can cope with it decrease progressively as saprobity increases, leading to a reduction of species richness and diversity [
18,
26]. Yet, OM input associated with low saprobity due to high oxygen availability or low by-product (e.g., ammonia and sulfide) concentrations can promote the vitality of the biocoenosis, increasing biomass and abundance.
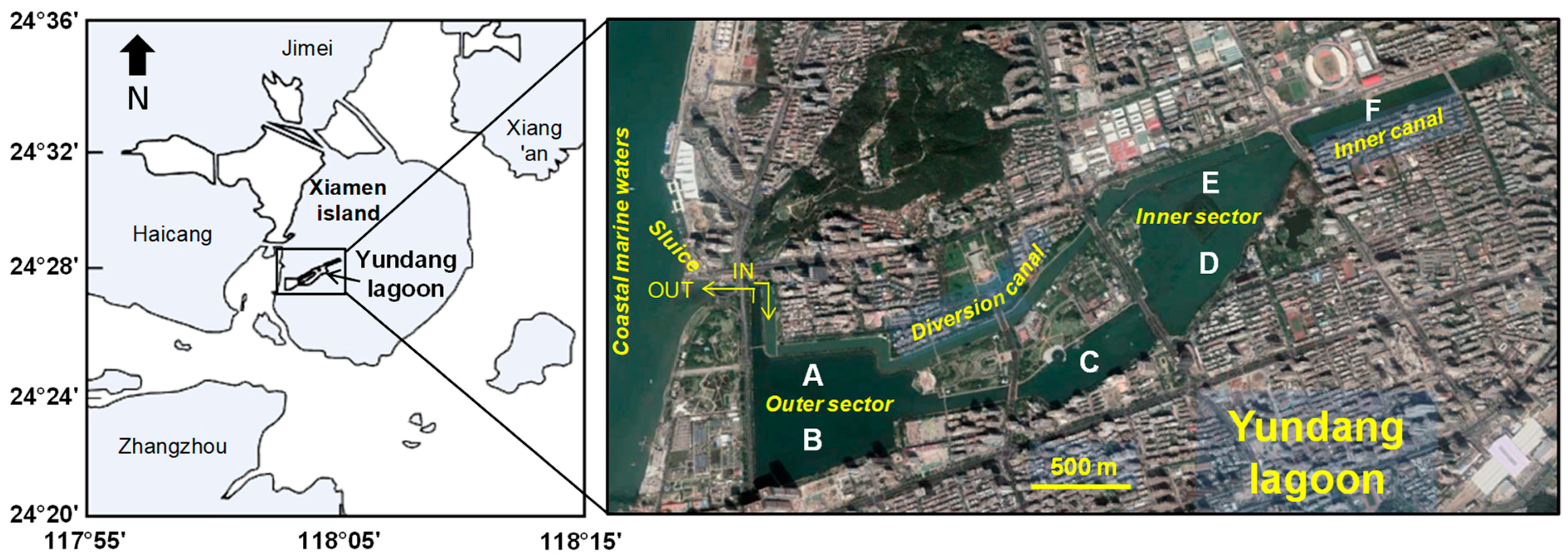
In China, many coastal areas are experiencing major urban and economic development, which is often coupled with growing anthropogenic pressures such as land reclamation, which often causes loss of coastal wetlands and serious environmental problems [
27]. Monitoring and assessment of man-made pollutants and their sources are, thus, crucial for evaluating the environmental quality of these areas, including coastal lagoons, located at the interface between continental and marine ecosystems. Accordingly, a substantial body of legislation exists to address environmental protection in China, such as laws on Water (21 January 1988) and Environmental Protection (26 December 1989), Sea Water Quality GB 3097-1997, Environmental Quality for Surface Water GB 3838-2002, and Provisions for Monitoring of Marine Culture and Propagation Areas (1 April 2002) [
28]. The Yundang Lagoon is an urban water body located in the highly populated and industrialized city of Xiamen (Southeast China). This lagoon, a natural bay open to the sea and historically rich in marine species, was severely damaged in the 1970s due to domestic and industrial pollution and land reclamation for agriculture. In particular, the Yundang Lagoon was reduced in size by more than 90% (from 10 km
2 before 1970 to 0.8 km
2 by 1988) and suffered decreased water flow and enhanced siltation, anoxia, loss of habitat and species, and the eventual collapse of the whole ecosystem [
29,
30,
31]. Thus, an integrated management program aimed at restoring the ecosystem was put in place in the late 1980s, which included clean-up activities, wastewater treatment, flood prevention, and sludge dredging. The lagoon was also opened to the outer bay of the Xiamen Sea through a diversion canal inlet in the early 1990s, coupled with new restoration actions including a mangrove plantation and phytoremediation techniques [
29,
32]. One of the main current aspirations of residents and the government is to restore the ecosystem services that a healthy Yundang Lagoon could provide [
29,
33].
Environmental and ecological studies in Chinese coastal lagoons are sparse, but growing steadily, owing to the scientific and socio-economic interest in these highly productive but fragile ecosystems [
27,
31]. Within this framework, in the present study the environmental and benthic features of the Yundang Lagoon were investigated as a unique case-study of a formerly “dead” lagoon which underwent a massive restoration effort over the past 30 years. We hypothesized that the long-term restoration interventions may have led to improved environmental and biological conditions of the lagoon. To address these issues, we analyzed a comprehensive set of physicochemical variables in water and sediments, as well as the macrozoobenthic assemblages, at six different sites across the Yundang Lagoon. Our objectives were to: (1) evaluate the trophic features of the lagoon and possible areas of major environmental concern; (2) investigate the pattern of spatial variation in the macrozoobenthic assemblages; and (3) assess the benthic recovery in relation to both the main environmental (e.g., trophic and saprobity) gradients and the presence of invasive alien species.
4. Discussion
For the first time, this study assesses the trophic features and benthic diversity of the Yundang Lagoon, an urban coastal lagoon in Southeast China, in a comprehensive and integrated manner. The Yundang Lagoon was referred to as a “dead” lagoon in the 1970s, and has undergone 30-year restoration effort. Microbiological and eco-toxicological studies conducted in the last decade suggest that the recovery of the Yundang Lagoon has not yet been accomplished. In particular, high levels of heavy metals and estrogenic compounds originating mainly from municipal wastewaters have been found in sediments and pore-water of the Yundang Lagoon posing serious toxic risk to the biota including humans [
44,
45,
46]. Furthermore, evidence has shown that the discharge of surrounding sewage is still a major source of OM pollution in the Yundang Lagoon [
34,
46].
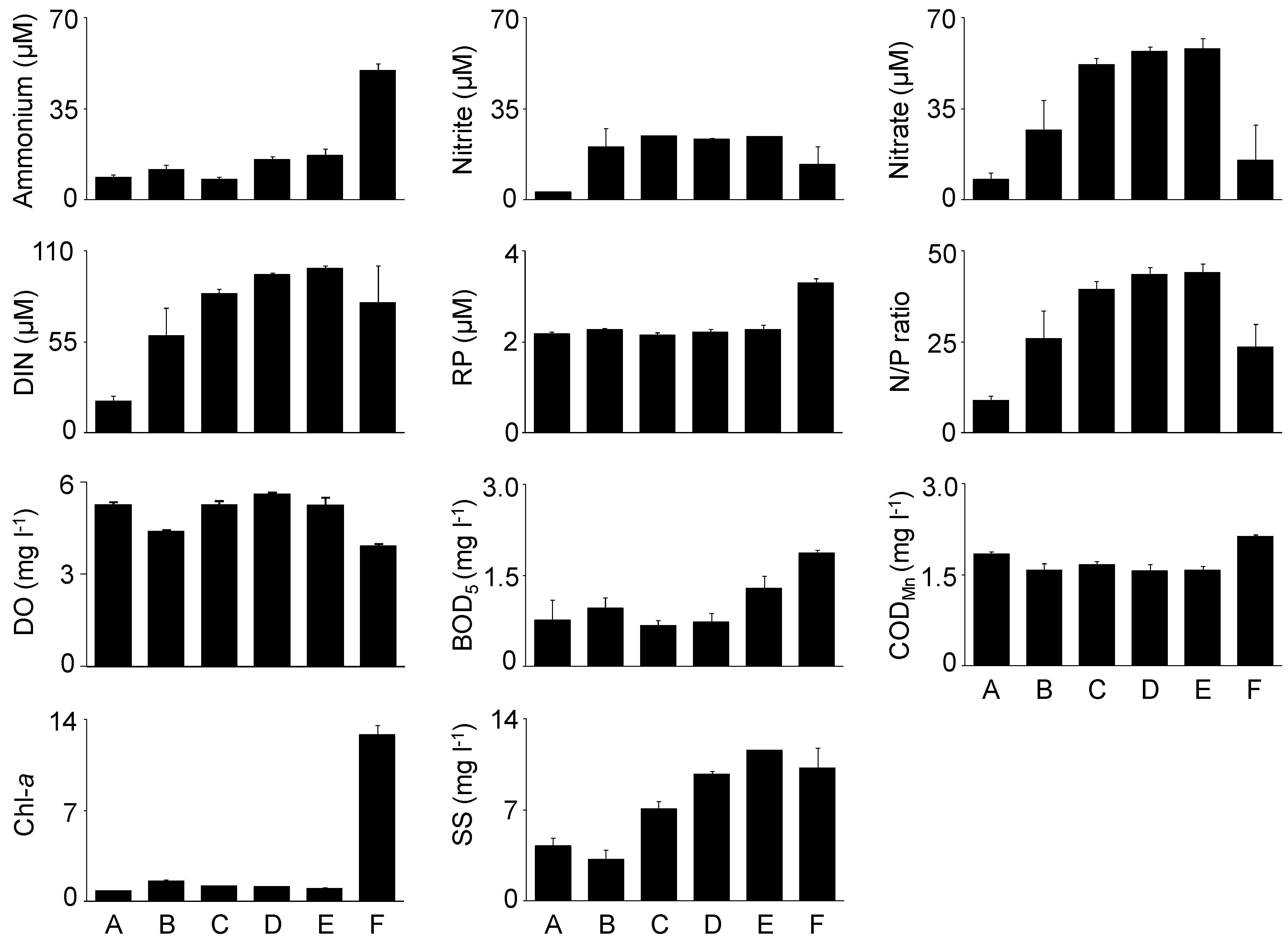
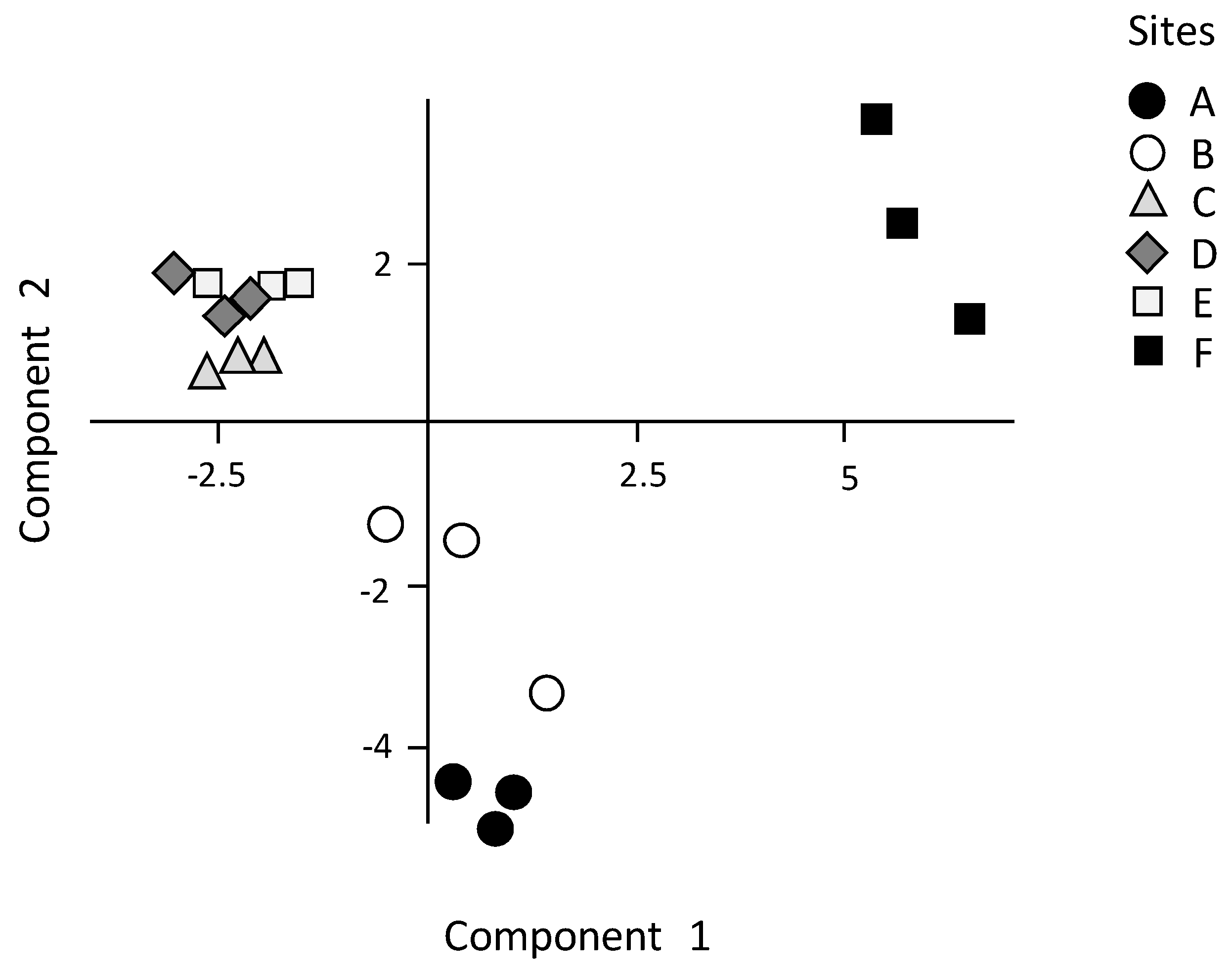
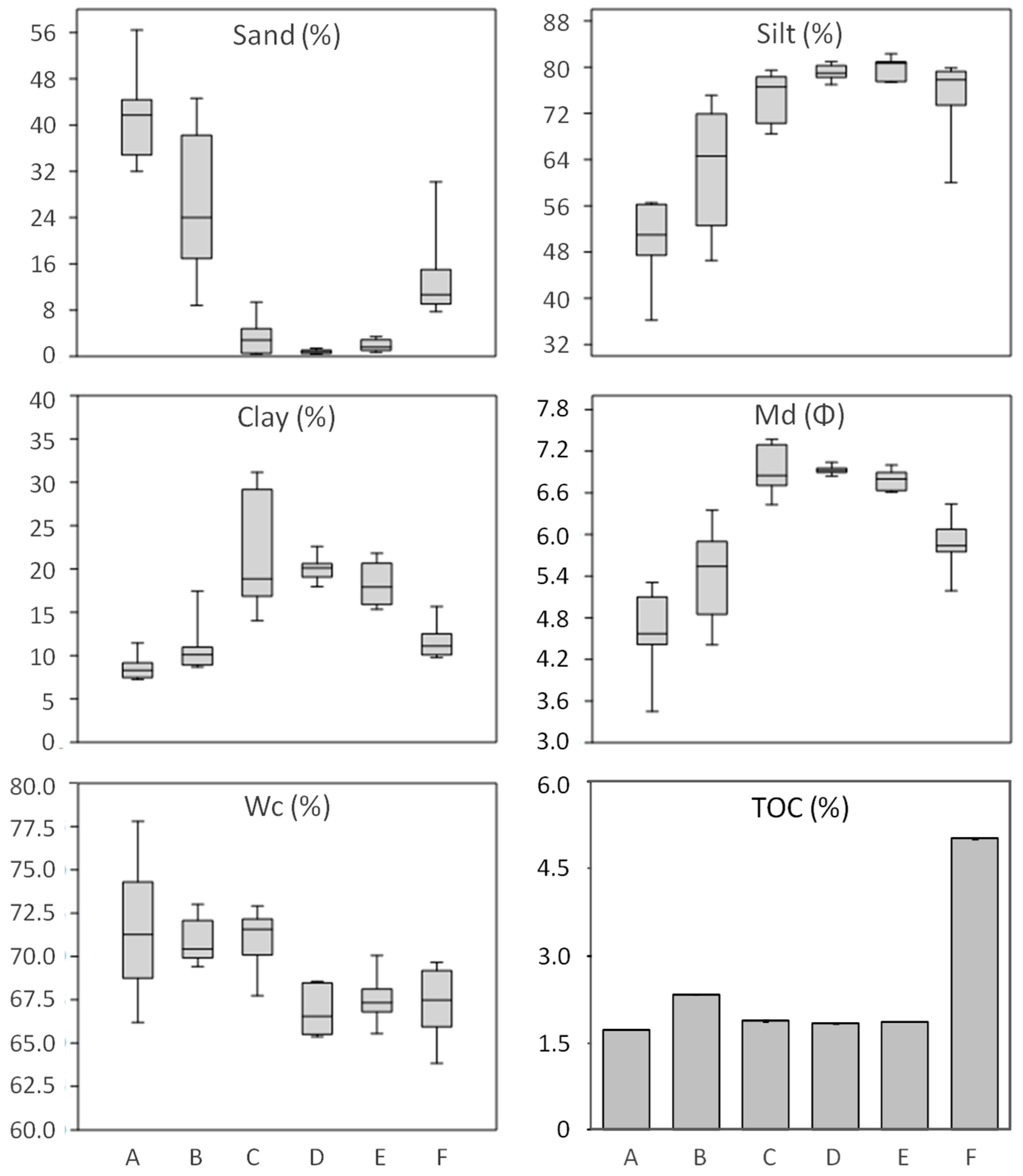
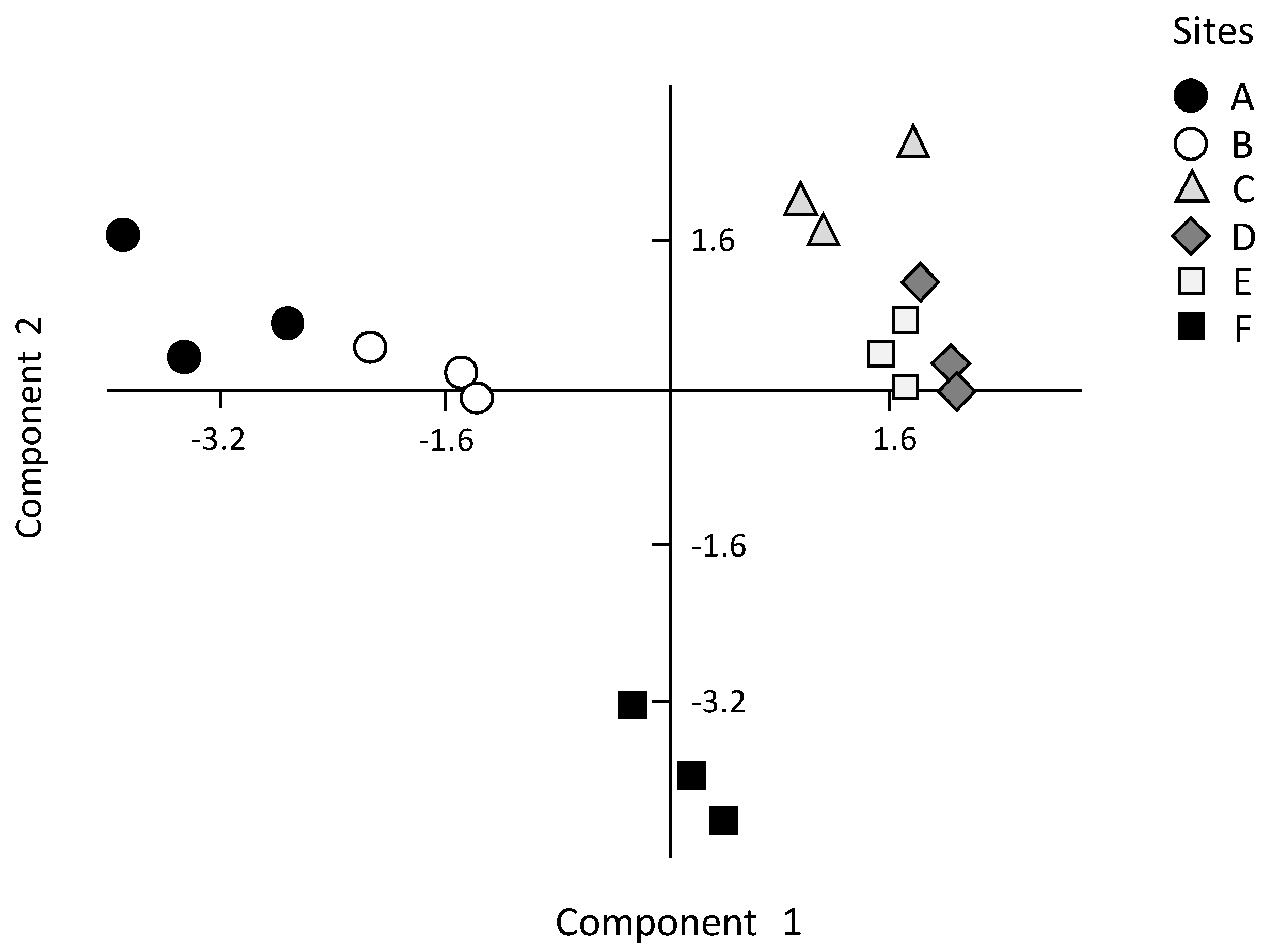
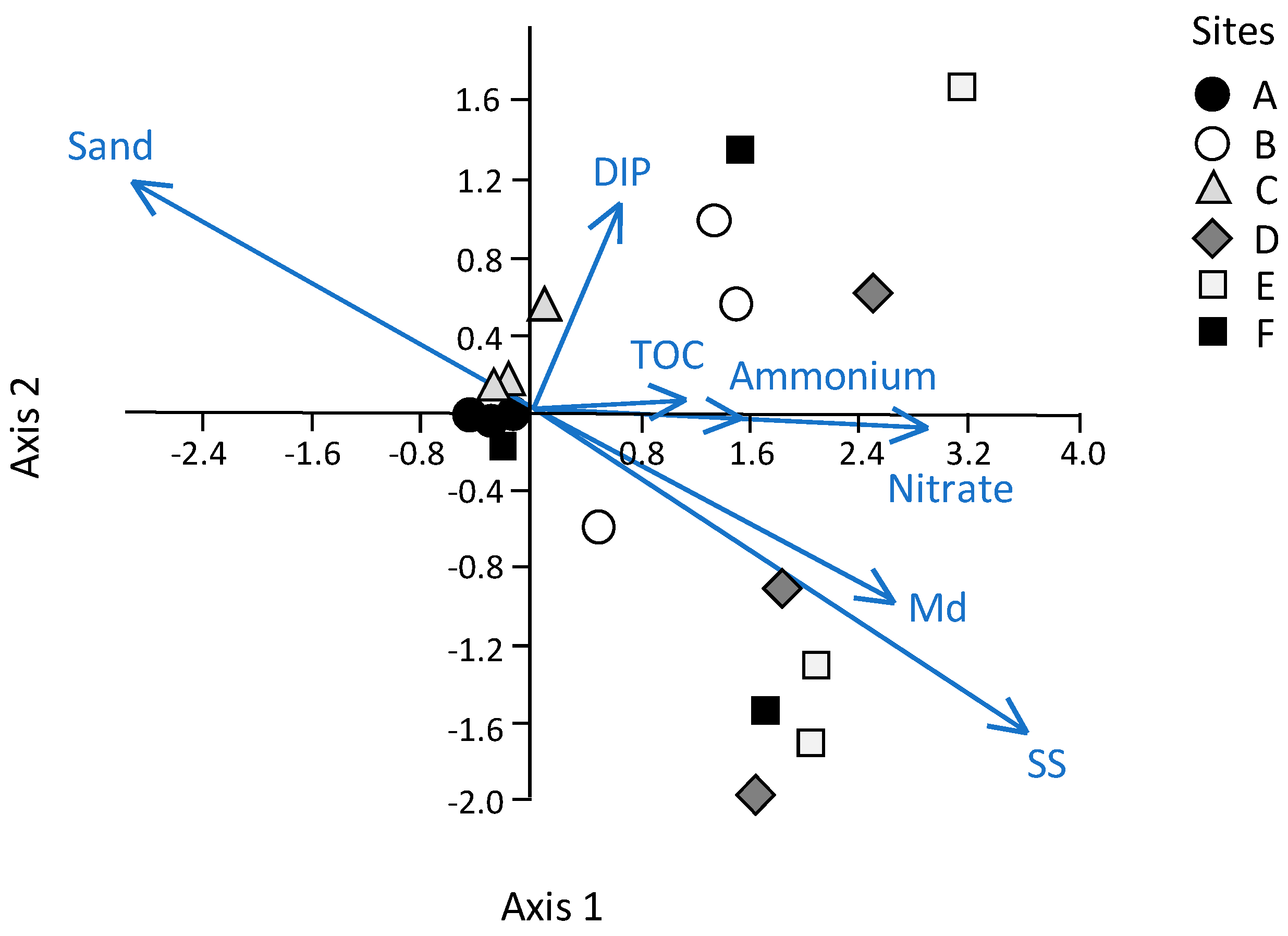
These results provide a detailed analysis of the trophic features of the Yundang Lagoon and highlight the high spatial heterogeneity of both abiotic (water and sediments) and biotic (macrozoobenthos) components, in spite of its small size and the absence of a salinity gradient. From an abiotic perspective, both water and sediment variables independently point in the same direction. In particular, there was a major separation between the low-nutrient and sandier sites A and B in the outer sector of the lagoon, and the hypertrophic and organically over-enriched site F in the inner canal. Site F departed the most from all the other sites, which was consistent with the fact that this sector of the lagoon is still subjected to numerous sewage discharges [
32,
47]. The confined location and reduced hydrodynamics of site F are indicated by high levels of suspended solids in the near-bottom water and a high silt content in surface sediments, making the site likely to favor the accumulation of toxic by-products derived from the microbial decomposition of high amounts of OM which were present above established critical thresholds [
24,
25]. Indeed, higher BOD
5 and COD
Mn concentrations measured at this site (where we also observed methane bubbling from the sediment) may be a concurrent evidence of high saprobity levels in this sector of the lagoon which, in turn, will severely affect the biota [
18]. On the other hand, all other sites of the lagoon showed significantly lower Chl-
a (in water) and TOC (in sediment) concentrations than those found at site F, notwithstanding seasonal variation, which was not considered here, during phytoplankton blooms [
34]. Interestingly, our results are consistent with those reported by Sun et al. [
33] who found spatial variation in Chl-
a and TOC to be greater in highly urbanized watersheds such as Yundang Lagoon, than in less urbanized watersheds. Overall, our study demonstrates that at the time samples were taken for this study (2012), the Yundang Lagoon had much lower nutrient, BOD
5, and COD
Mn concentrations than those found in previous decades [
29], supporting the inference that a partial, but significant environmental recovery has occurred.
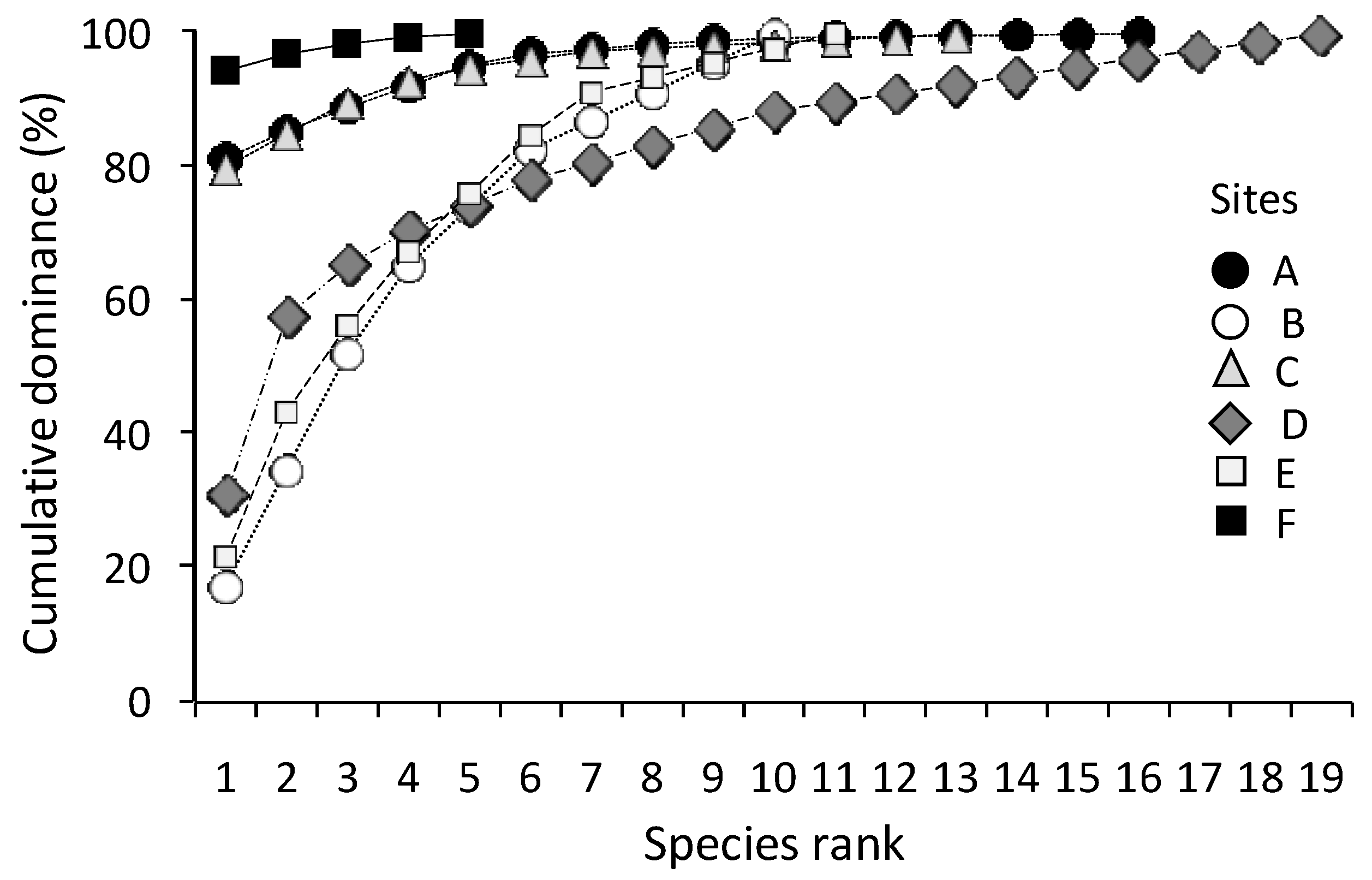
The overall improvement of the environmental condition of the Yundang Lagoon was reflected in a major recovery and revitalization of the lagoon’s soft-bottom benthic assemblage during the last decade. Following anoxic and azoic sediments found when the lagoon was highly polluted and closed to the adjacent coastal waters until 1987 [
29], a single polychaete species,
Neanthes succinea, typical of organically enriched sediments with high H
2S content [
14,
48], was reported during the first phase of the restoration program (1989−1992). Between 2001 and 2008, nine species started to colonize the lagoon’s sediments [
43]. Among them, the opportunistic polychaetes
Capitella capitata and
Neanthes glandicincta, typical of muddy, organically enriched sediments were dominant, while they occurred with few individuals in our study. These results show a significant recovery of the macrozoobenthic assemblages for the first time, with a marked increase in species richness and diversity, yet a heterogeneous distribution across the lagoon. Of the 43 species found, eight were dominant (accounting for 93% of the total abundance), mainly represented by mollusks and amphipods, and including the polychaete
Cossurella dimorpha which was not reported previously. Overall, the most dominant species was the bivalve Dreissenidae
Mytilopsis sallei, an invasive alien species native to the Caribbean and a massive component of fouling events [
49,
50].
M. sallei was transported to China via ballast waters, and was first recorded in Xiamen waters (Maluan Bay) in 1990 and in the Yundang Lagoon in 2000 [
51,
52,
53]. In a basin as small as the Yundang Lagoon, one may expect little spatial variation in the soft-bottom benthic assemblages. However, our extensive sampling effort allowed us to identify significant variation in the macrozoobenthic assemblages within relatively short distances of a few hundred meters. A number of environmental and biological factors may help explaining the high spatial variability and the recovery of the macrozoobenthic assemblages in the Yundang Lagoon. These are: (1) the heterogeneous morphology and the peculiar hydrology of the lagoon, including the daily water exchange with the adjacent coastal sea, (2) the high environmental variability in water and sediments across the lagoon, (3) the presence of nonpoint sources of pollution and other anthropogenic pressures, and (4) the dominance of the invasive
M. sallei and the biotic interactions within the benthic community. Thus, the present work represents a unique case-study to evaluate the complex dynamics of macrozoobenthic assemblages in a formerly dead urban coastal lagoon following long-term restoration.
As an example, sites A and B in the outer sector of the lagoon, although similar in several environmental features and, thus, significantly different from the other sites, were the least similar to one another in terms of benthic species composition, richness and diversity. In particular, site A was the most abundant, biomass-rich, and species-rich site in the lagoon, while the opposite was true for site B, which had a species richness and diversity as low as that found at the most degraded site, F. We infer that such biotic differences between sites A and B may be related to their position within the outer sector of the lagoon. In particular, site B was located near the heavily modified shoreline made by a concrete wall, following earlier dredging and reclamation, which may have left little of the historic ecohydrological system on this site of the lagoon. This may have contributed to determine a poor macrozoobenthic assemblage which included few species typical of muddy sediments, such as
Cossurella dimorpha,
Corophium sp. and
Gammaropsis sp. On the other hand, site A was located closer to the shoreline where mangrove species, such as
Kandelia candel,
Avicennia marina,
Rhizophora stylosa, and
Heritiera littoralis, had been successfully planted from 2001 to 2006 [
54], contributing to increases of birds and fish species in the lagoon [
29]. It is likely that the presence of mangroves along this shore has favored the colonization of the invasive fouling species
M. sallei. This bivalve was associated with the richest benthic assemblage of the lagoon, correlating with the highly diverse assemblages found at site A which also included species typical of mangrove habitats such as
R. plicatula and
S. glabra. The diversity of the site was also seen in
Corophium uenoi, which lives within muddy tubes on soft bottom sediments, but is also present in the fouling community, and
P. tsurumaru, which is typical of muddy sediments and possibly shares the burrow of holothurian or crabs (commensal). On the contrary, the innermost site F had the most impoverished assemblage, consistent with its degraded condition in both water and sediments. Yet,
M. sallei was also found at one station at this site, providing evidence of its high resistance and resilience, and its ability to settle when a substrate is available, owing to the characteristic traits of opportunistic
r-strategists [
49,
50]. The apparently random distribution of
M. sallei, heavily colonizing some sectors of the lagoon, was also indicated by its dominance at site C, where environmental conditions were intermediate between the outer (sites A and B) and the innermost (site F) sectors. This indicates that, in addition to the environmental factors, species-specific biological interactions should be considered to evaluate the complex dynamics that occur within a benthic assemblage.
Our results suggest that
M. sallei, known as an invasive ecosystem engineer that creates new habitats and favors the settlement of other species [
55], may support a rich and diversified community. In particular, in the present study
M. sallei was associated with the highest number of species having different biological traits (e.g., fouling, parasite and commensal) and typical of different habitats (e.g., muddy sediments, mangroves). Among them,
S. glabra, an IUCN threatened species [
56] in need of protection measures, and
P. tsurumaru which is known to establish symbiotic relations with other species [
57]. Nevertheless,
M. sallei is also well known as a pest species characterized by wide temperature, salinity, and oxygen tolerances, as well as a fast growth-rate and a high fecundity which may favor competition with native species and inhibit the growth of other species [
49]. As an example, an experimental field study on the fouling macrofauna associated with
M. sallei conducted in the Yundang Lagoon showed a reduction in species diversity in summer when environmental conditions worsened, but the density and biomass of both
M. sallei and fouling macrofauna were highest [
52]. Furthermore,
M. sallei is known to erode fishing facilities and artificial structures leading to ecosystem damage, economic loss, and inconvenience to locals [
53], thus, its spatial and temporal variation should be carefully monitored.
With regards to the spatiotemporal evolution of the macrozoobenthic assemblages in the Yundang Lagoon, one possibility is an even more massive colonization of the lagoon by
M. sallei as the most dominant species. The successful establishment of
M. sallei in the Yundang Lagoon may prefigure a wider, yet probably overlooked colonization of this species as a structural component of the benthic assemblages in other Chinese coastal waters and lagoons for which information is very scant [
53]. A similar colonization process occurred in the Mediterranean Sea during the first half of the last century by Serpulidae
Ficopomatus enigmaticus which is now a typical bioengineer species in Mediterranean lagoons [
58,
59,
60]. However, after a period of dominance,
M. sallei may decrease in dominance or even be replaced by other species typical of sessile fouling, such as
S. plicata. A similar alternation of species has been found between the mussel
Mytilus edulis and the polychaete
Sabellaria spinulosa along the eastern Atlantic coast, and in the Mediterranean Sea between
M. galloprovincialis and the extensive bioconstructions of
S. spinulosa along the Italian Puglia coast [
61,
62].

 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}