Role of Groundwater-Borne Geogenic Phosphorus for the Internal P Release in Shallow Lakes



Abstract
:1. Introduction
2. Materials and Methods
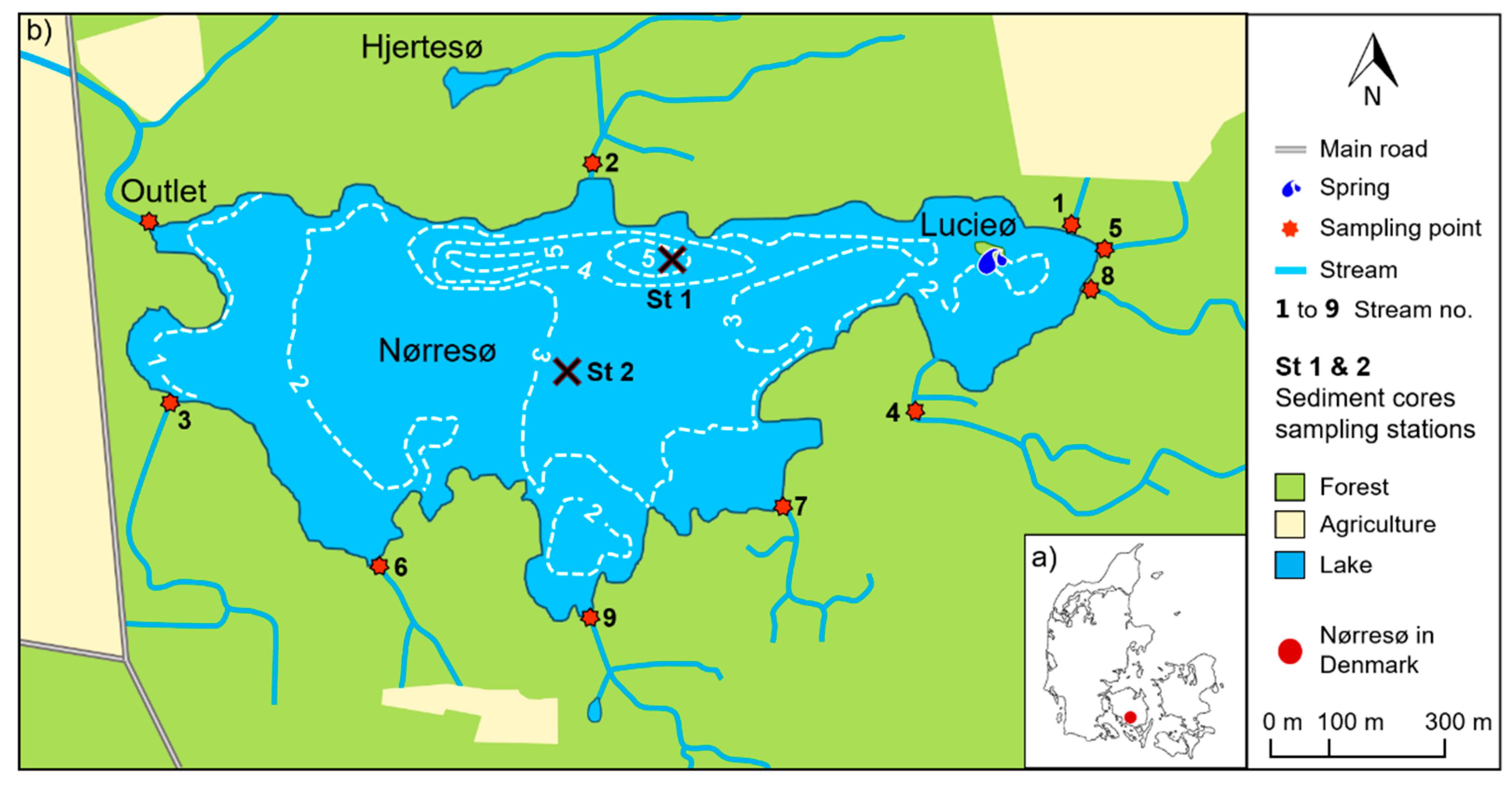
2.1. Study Site
2.2. Data Collection
2.2.1. External Phosphorus (P) Loadings
2.2.2. Water Sampling and Dissolved P Analysis
2.2.3. Mass-Loading Model of lake Water Column P
2.2.4. Estimation of the Internal P Release Load
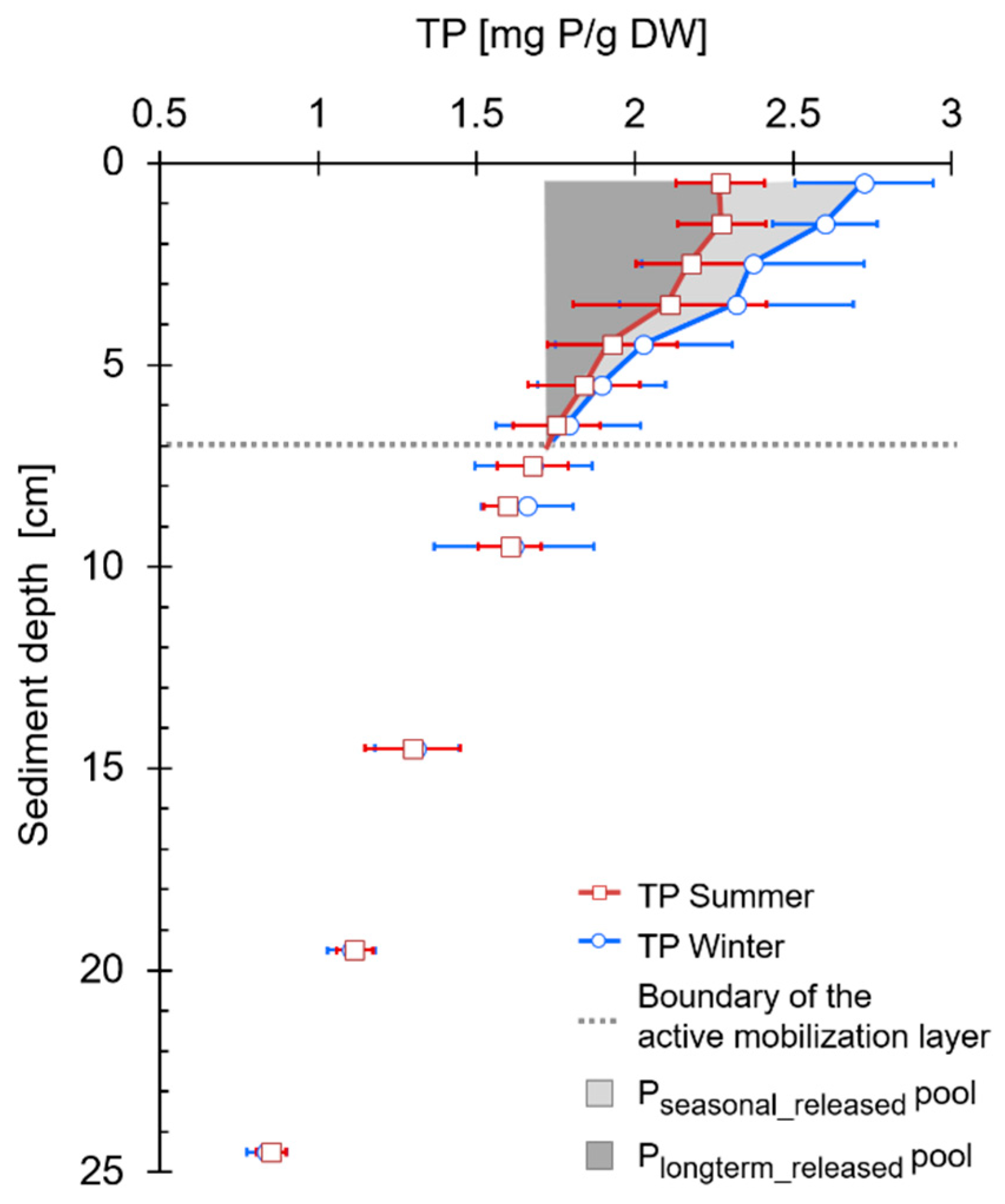
2.2.5. Total P Sediment Depth Profiles
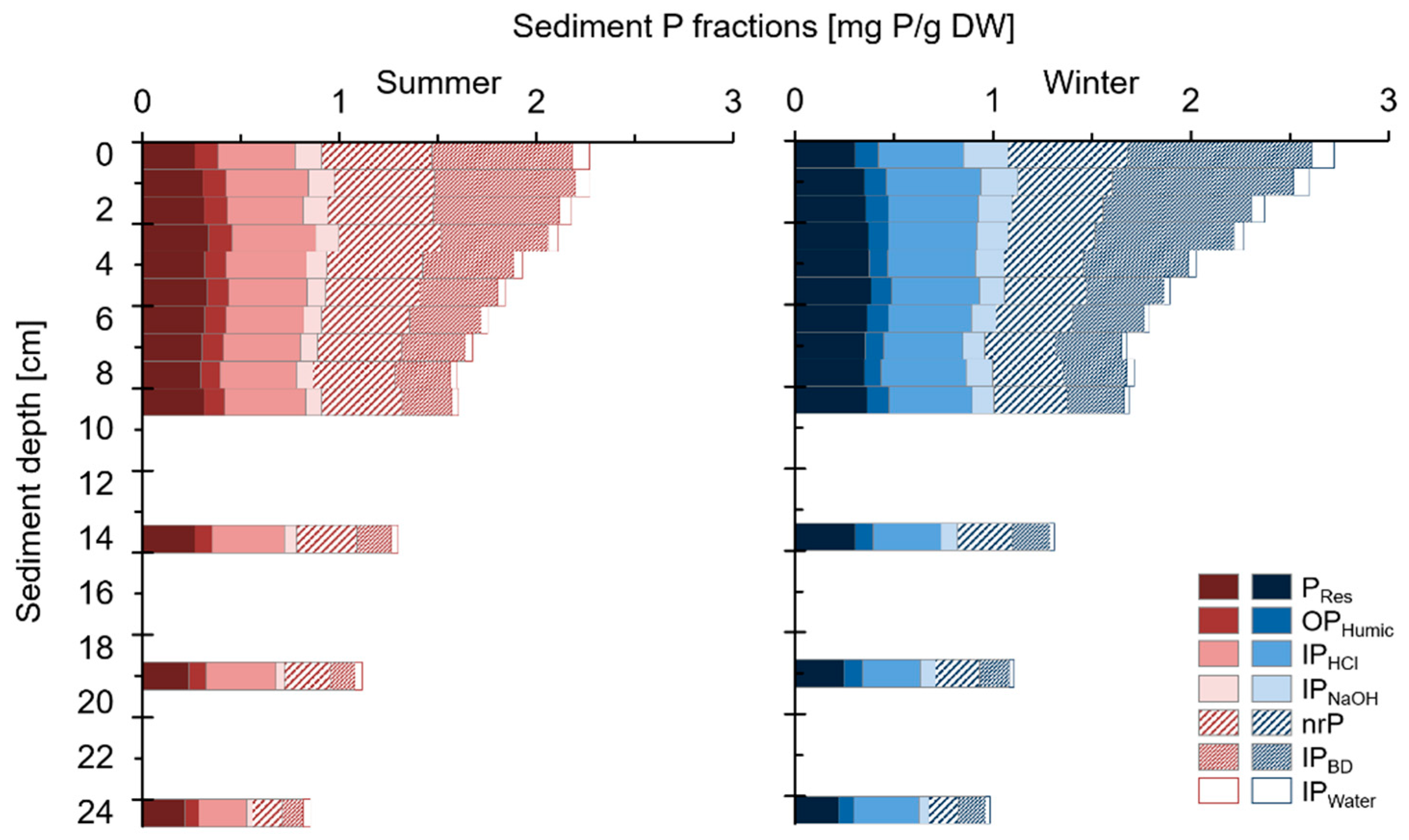
2.2.6. Sediment-P Composition
3. Results
3.1. External Waterborne Dissolved P Loadings
3.2. Internal P Release Load
3.3. Sediment-P Composition
3.4. Dissolved P Concentrations in the Lake, Spring, and Streams
4. Discussion
4.1. Current Lake P Budget
4.1.1. External P Loadings
4.1.2. Internal P Loading
4.2. Long-Term Influence of External TDP
5. Conclusions
- Geogenic groundwater-borne P can be the dominant cause of P accumulation: the amount of geogenic groundwater-borne P brought to the lake during Holocene is in the same order of magnitude as the total accumulated P in the sediment.
- Even though the seasonal release of internal P controls seasonal fluctuations in water column P, groundwater-borne geogenic P may be the actual eutrophicating factor.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blake, R.E.; O’Neil, J.R.; Surkov, A.V. Biogeochemical cycling of phosphorus: Insights from oxygen isotope effects of phosphoenzymes. Am. J. Sci. 2005, 305, 596–620. [Google Scholar] [CrossRef]
- Hecky, R.E.; Kilham, P. Nutrient limitation of phytoplankton in freshwater and marine environments: A review of recent evidence on the effects of enrichment1. Limnol. Oceanogr. 1988, 33, 796–822. [Google Scholar] [CrossRef] [Green Version]
- Schindler, D. Eutrophication and recovery in experimental lakes—Implications for lake management. Science. 1974, 184, 897–899. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, R.G. The phosphorus cycle. In Limnology: Lake and River Ecosystems; Academic Press: San Diego, CA, USA, 2001; pp. 242–250. ISBN 9780127447605. [Google Scholar]
- Søndergaard, M.; Jensen, J.P.; Jeppesen, E. Role of sediment and internal loading of phosphorus in shallow lakes. Hydrobiologia 2003, 506–509, 135–145. [Google Scholar] [CrossRef]
- Smolders, A.J.P.; Lamers, L.P.M.; Lucassen, E.C.H.E.T.; Van Der Velde, G.; Roelofs, J.G.M. Internal eutrophication: How it works and what to do about it—A review. Chem. Ecol. 2006, 22, 93–111. [Google Scholar] [CrossRef]
- Boström, B.; Andersen, J.M.; Fleischer, S.; Jansson, M. Exchange of phosphorus across the sediment-water interface. Hydrobiologia 1988, 170, 229–244. [Google Scholar] [CrossRef]
- Paytan, A.; Roberts, K.; Watson, S.; Peek, S.; Chuang, P.C.; Defforey, D.; Kendall, C. Internal loading of phosphate in lake erie central basin. Sci. Total Environ. 2017, 579, 1356–1365. [Google Scholar] [CrossRef] [PubMed]
- Van der Molen, D.T.; Boers, P.C.M. Influence of internal loading on phosphorus concentration in shallow lakes before and after reduction of the external loading. Hydrobiologia 1994, 275, 379–389. [Google Scholar] [CrossRef]
- Murray, G.C.; Hesterberg, D. Iron and phosphate dissolution during abiotic reduction of ferrihydrite-boehmite mixtures. Soil Sci. Soc. Am. J. 2006, 70, 1318. [Google Scholar] [CrossRef]
- Hupfer, M.; Gloess, S.; Grossart, H.P. Polyphosphate-accumulating microorganisms in aquatic sediments. Aquat. Microb. Ecol. 2007, 47, 299–311. [Google Scholar] [CrossRef]
- Hupfer, M.; Lewandowski, J. Oxygen controls the phosphorus release from lake sediments—A long-lasting paradigm in limnology. Int. Rev. Hydrobiol. 2008, 93, 415–432. [Google Scholar] [CrossRef]
- Hansen, K.; Mouridsen, S.; Kristensen, E. The impact of Chironomus plumosus larvae on organic matter decay and nutrient (N, P) exchange in a shallow eutrophic lake sediment following a phytoplankton sedimentation. Hydrobiologia 1998, 364, 65–74. [Google Scholar] [CrossRef]
- Golterman, H.L. The calcium-and iron bound phosphate phase diagram. Hydrobiologia 1988, 159, 149–151. [Google Scholar] [CrossRef]
- Penn, M.R.; Auer, M.T.; Doerr, S.M.; Driscoll, C.T.; Brooks, C.M.; Effler, S.W. Seasonality in phosphorus release rates from the sediments of a hypereutrophic lake under a matrix of pH and redox conditions. Can. J. Fish. Aquat. Sci. 2000, 57, 1033–1041. [Google Scholar] [CrossRef]
- House, W.A. Geochemical cycling of phosphorous in rivers. Appl. Geochem. 2003, 18, 739–748. [Google Scholar] [CrossRef]
- Heiberg, L.; Pedersen, T.V.; Jensen, H.S.; Kjaergaard, C.; Hansen, H.C.B. A comparative study of phosphate sorption in lowland soils under oxic and anoxic conditions. J. Environ. Qual. 2010, 39, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Kjaergaard, C.; Heiberg, L.; Jensen, H.S.; Hansen, H.C.B. Phosphorus mobilization in rewetted peat and sand at variable flow rate and redox regimes. Geoderma 2012, 173–174, 311–321. [Google Scholar] [CrossRef]
- Prem, M.; Hansen, H.C.B.; Wenzel, W.; Heiberg, L.; Sørensen, H.; Borggaard, O.K. High spatial and fast changes of iron redox state and phosphorus solubility in a seasonally flooded temperate wetland soil. Wetlands 2015, 35, 237–246. [Google Scholar] [CrossRef]
- Jensen, H.S.; Andersen, F.O. Importance of temperature, nitrate, and pH for phosphate release from aerobic sediments of four shallow, eutrophic lakes. Limnol. Oceanogr. 1992, 37, 577–589. [Google Scholar] [CrossRef]
- Bennion, H.; Smith, M.A. Variability in the water chemistry of shallow ponds in southeast England, with special reference to the seasonality of nutrients and implications for modelling trophic status. Hydrobiologia 2000, 436, 145–158. [Google Scholar] [CrossRef]
- Ekholm, P.; Malve, O.; Kirkkala, T. Internal and external loading as regulators of nutrient concentrations in the agriculturally loaded lake Pyhajarvi (southwest Finland). Hydrobiologia 1997, 345, 3–14. [Google Scholar] [CrossRef]
- Søndergaard, M.; Jensen, P.J.; Jeppesen, E. Retention and internal loading of phosphorus in shallow, eutrophic lakes. Sci. World 2001, 1, 427–442. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, E.; Kristensen, P.; Jensen, J.P.; Søndergaard, M.; Mortensen, E.; Lauridsen, T. Recovery resilience following a reduction in external phosphorus loading of shallow, eutrophic Danish lakes: Duration, regulating factors and methods for over-coming resilience. Mem. Ist. Ital. Idrobiol. 1991, 48, 127–148. [Google Scholar]
- Van der Grift, B.; Osté, L.; Schot, P.; Kratz, A.; Van Popta, E.; Wassen, M.; Griffioen, J. Forms of phosphorus in suspended particulate matter in agriculture-dominated lowland catchments: Iron as phosphorus carrier. Sci. Total Environ. 2018, 631, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Heathwaite, A.L.; Burke, S.P.; Bolton, L. Field drains as a route of rapid nutrient export from agricultural land receiving biosolids. Sci. Total Environ. 2006, 365, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Heathwaite, A.L.; Dils, R.M. Characterising phosphorus loss in surface and subsurface hydrological pathways. Sci. Total Environ. 2000, 251, 523–538. [Google Scholar] [CrossRef]
- Withers, P.J.A.; Haygarth, P.M. Agriculture, phosphorus and eutrophication: A European perspective. Soil Use Manag. 2007, 23, 1–4. [Google Scholar] [CrossRef]
- Hodgkinson, R.A.; Chambers, B.J.; Withers, P.J.A.; Cross, R. Phosphorus losses to surface waters following organic manure applications to a drained clay soil. Agric. Water Manag. 2002, 57, 155–173. [Google Scholar] [CrossRef]
- Gilliom, R.J.; Patmont, C.R. Lake phosphorus loading from septic systems by seasonally perched groundwater. Water Pollut. Control. Fed. 1983, 55, 1297–1305. [Google Scholar]
- Zanini, L.; Robertson, W.D.; Ptacek, C.J.; Schiff, S.L.; Mayer, T. Phosphorus characterization in sediments impacted by septic effluent at four sites in central Canada. J. Contam. Hydrol. 1998, 33, 405–429. [Google Scholar] [CrossRef]
- Burkart, M.R.; Simpkins, W.W.; Morrow, A.J.; Gannon, J.M. Occurrence of total dissolved phosphorus in unconsolidated aquifers and aquitards in Iowa. J. Am. Water Resour. Assoc. 2004, 40, 827–834. [Google Scholar] [CrossRef]
- Holman, I.P.; Whelan, M.J.; Howden, N.J.K.; Bellamy, P.H.; Willby, N.J.; Rivas-Casado, M.; McConvey, P. Phosphorus in groundwater—An overlooked contributor to eutrophication? Hydrol. Process. 2008, 22, 5121–5127. [Google Scholar] [CrossRef]
- Holman, I.P.; Howden, N.J.K.; Bellamy, P.; Willby, N.; Whelan, M.J.; Rivas-Casado, M. An assessment of the risk to surface water ecosystems of groundwater P in the UK and Ireland. Sci. Total Environ. 2009, 408, 1847–1857. [Google Scholar] [CrossRef] [PubMed]
- Kilroy, G.; Coxon, C. Temporal variability of phosphorus fractions in Irish karst springs. Environ. Geol. 2005, 47, 421–430. [Google Scholar] [CrossRef]
- Kidmose, J.; Nilsson, B.; Engesgaard, P.; Frandsen, M.; Karan, S.; Landkildehus, F.; Søndergaard, M.; Jeppesen, E. Focused groundwater discharge of phosphorus to a eutrophic seepage lake (Lake Væng, Denmark): Implications for lake ecological state and restoration. Hydrogeol. J. 2013, 21, 1787–1802. [Google Scholar] [CrossRef]
- Lewandowski, J.; Meinikmann, K.; Nützmann, G.; Rosenberry, D.O. Groundwater—The disregarded component in lake water and nutrient budgets. Part 2: Effects of groundwater on nutrients. Hydrol. Process. 2015, 29, 2922–2955. [Google Scholar] [CrossRef]
- Walter, D.A.; Rea, B.A.; Sollenwerk, K.G.; Savoie, J. Geochemical and Hydrologic Controls on Phosphorus Transport. in a Sewage Contaminated Sand and Gravel Aquifer Near Ashumet Pond, Cape Cod, Massachusetts; Technical report No. 95-381; United States Geological Survey Water-Supply Paper 2463: Washington, DC, USA, 1996.
- Kenoyer, G.; Anderson, M.P. Groundwater’s dynamiv role in regulating acidity and chemistry in a precipitatuon-dominated lake. J. Hydrol. 1989, 109, 287–306. [Google Scholar] [CrossRef]
- Vanek, V. The interactions between lake and groundwater and their ecological significance. Stygologia 1987, 3, 1–23. [Google Scholar]
- Shaw, R.D.; Shaw, J.F.H.; Fricker, H.; Prepas, E.E. An integrated approach to quantify groundwater transport of phosphorus to Narrow Lake, Alberta. Limnol. Oceanogr. 1990, 35, 870–886. [Google Scholar] [CrossRef]
- Meinikmann, K.; Lewandowski, J.; Hupfer, M. Phosphorus in groundwater discharge - A potential source for lake eutrophication. J. Hydrol. 2015, 524, 214–226. [Google Scholar] [CrossRef]
- Heathwaite, A.L.; Dils, R.M.; Liu, S.; Carvalho, L.; Brazier, R.E.; Pope, L.; Hughes, M.; Phillips, G.; May, L. A tiered risk-based approach for predicting diffuse and point source phosphorus losses in agricultural areas. Sci. Total Environ. 2005, 344, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Orderud, G.I.; Vogt, R.D. Trans-disciplinarity required in understanding, predicting and dealing with water eutrophication. Int. J. Sustain. Dev. World Ecol. 2013, 20, 404–415. [Google Scholar] [CrossRef] [Green Version]
- Sharpley, A.N.; Daniel, T.; Sims, T.; Lemunyon, J.; Stevens, R.; Parry, R. Agricultural Phosphorus and Eutrophication, 2nd ed.; U.S. Department of Agriculture, Agricultural Research Service: Lincoln, NE, USA, 2003.
- Smith, V.H.; Schindler, D.W. Eutrophication science: Where do we go from here? Trends Ecol. Evol. 2009, 24, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Høy, T. Danmarks Søer—Søerne i Fyns Amt; Strandbergs Forlag: Charlottenlund, Denmark, 2000. [Google Scholar]
- Nisbeth, C.S.; Kidmose, J.; Weckström, K.; Reitzel, K.; Odgaard, B.V.; Thorling, L.; McGowan, S.; Schomacker, A.; Kristensen, D.L.J.; Jessen, S. Dissolved inorganic geogenic phosphorus load to a groundwater-fed lake: Implications of terrestrial phosphorus cycling by groundwater. Spec. Issues Water 2019, in press. [Google Scholar]
- GEUS. (Geological Survey of Denmark and Greenland) 2018 Danish National Borehole Achieve (Jupiter). Available online: www.geus.dk (accessed on 21 September 2018).
- NOVANA. (Det Nationale Overvågningsprogram for Vandmiljø og Natur). Available online: http://novana.dmi.dk/ (accessed on 3 April 2018).
- Bennike, O.; Odgaard, B.V.; Eriksen, A.A.; McGowan, S.; Siggaard-Andersen, M.L.; Schomacker, A.; Jessen, S.; Kazmierczak, J.; Olsen, J.; Rasmussen, P.; et al. Multi-proxy records of late Quaternary environmental changes based on lake sediment records from Vængsø and Nørresø, Denmark. Unpublished work.
- Eriksen, A.A. Holocene Catchment Changes at and Brahetrolleborg Nørresø and Lake Eutrophication Response. Master’s Thesis, Aarhus University, Aarhus, Denmark, 2017. [Google Scholar]
- Fyn’s County. Nørresø 1989–1993, Lake Monitoring in Fyn’s County; Technical report No.1; Fyn’s County: Odense, Denmark, 1994. [Google Scholar]
- Vollenweider, R.A. Advances in defining critical loading levels for phosphorus in lake eutrophication. Mem. dell’Istituto Ital. Idrobiol. Dott. Marco Marchi Verbania Pallanza 1976, 33, 53–83. [Google Scholar]
- ODA. Surface Water Monitoring, Aarhus University. Available online: https://odaforalle.au.dk/main.aspx (accessed on 25 February 2019).
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1986, 27, 31–36. [Google Scholar] [CrossRef]
- Vollenweider, R.A. Input-output models with special reference to the phosphorus loading concept in limnology. Schweiz. Z. Hydrol. 1975, 37, 53–84. [Google Scholar]
- Brett, M.T.; Benjamin, M.M. A review and reassessment of lake phosphorus retention and the nutrient loading concept. Freshw. Biol. 2008, 53, 194–211. [Google Scholar] [CrossRef]
- DMI. The Danish Meteorological Institute. Available online: https://www.dmi.dk/vejrarkiv/ (accessed on 25 February 2019).
- Renberg, I.; Hansson, H. The HTH sediment corer. J. Paleolimnol. 2008, 40, 655–659. [Google Scholar] [CrossRef]
- Paludan, C.; Jensen, H.S. Sequential extractiom of phosphorus in freshwater wetland and lake sediment: Significance of humic acids. Wetlands 1995, 15, 365–373. [Google Scholar] [CrossRef]
- Rydin, E. Potentially mobile phosphorus in Lake Erken sediment. Water Res. 2000, 34, 2037–2042. [Google Scholar] [CrossRef]
- Reitzel, K. Separation of aluminum bound phosphate from iron bound phosphate in freshwater sediment by sequential extraction procedure. Phosphates Sediments 2005, 109–117. [Google Scholar]
- Ellermann, T.; Bossi, R.; Christensen, J.; Løfstrøm, P.; Monies, C.; Grundahl, L.; Geels, C. Atmosfærisk Deposition 2014; Technical report No. 163; Department of Environmental Sciences (DCE): Aarhus, Denmark, 2015. [Google Scholar]
- Søndergaard, M.; Jensen, J.P.; Jeppesen, E. Internal phosphorus loading in shallow Danish lakes. Hydrobiologia 1999, 408/409, 145–152. [Google Scholar] [CrossRef]
- Reitzel, K.; Hansen, J.; Andersen, F.; Hansen, K.S.; Jensen, H.S. Lake restoration by dosing aluminum relative to mobile phosphorus in the sediment. Environ. Sci. Technol. 2005, 39, 4134–4140. [Google Scholar] [CrossRef] [PubMed]
- Griffioen, J. Extent of immobilisation of phosphate during aeration of nutrient-rich, anoxic groundwater. J. Hydrol. 2006, 320, 359–369. [Google Scholar] [CrossRef]
- Marion, L.; Clergeau, P.; Brient, L.; Bertru, G. The importance of avian-contributed nitrogen (N) and phosphorus (P) to Lake Grand-Lieu, France. Hydrobiologia 1994, 279–280, 133–147. [Google Scholar] [CrossRef]
- Gwiazda, R.; Jarocha, K.; Szarek-Gwiazda, E. Impact of a small cormorant (Phalacrocorax carbo sinensis) roost on nutrients and phytoplankton assemblages in the littoral regions of a submontane reservoir. Biologia 2010, 65, 742–748. [Google Scholar] [CrossRef]
- Søndergaard, M.; Lauridsen, T.L. Fugle og Karpers Påvirkning af Søer; Technical report No. 84; Department of Environmental Sciences (DCE): Aarhus, Denmark, 2014. [Google Scholar]
- Phillips, G.; Jackson, R.; Bennett, C.; Chilvers, A. The importance of sediment phosphorus release in the restoration of very shallow lakes (The Norfolk Broads, England) and implications for biomanipulation. Hydrobiologia 1994, 275–276, 445–456. [Google Scholar] [CrossRef]
- Welch, E.B.; Cooke, G.D. Internal phosphorus loading in shallow lakes: Importance and control. Lake Reserv. Manag. 2005, 21, 209–217. [Google Scholar] [CrossRef]
- Odgaard, B.; Møller, P.F.; Wolin, J.A.; Rasmussen, P.; Anderson, N.J. Brahetrolleborg Nørresø. Palæolimnologisk Undersøgelse og Oplandsanalyse; Company Report (48); Denmark’s Geological Surveys: Copenhagen, Denmark, 1995; pp. 1–56. [Google Scholar]
- Ding, S.; Wang, Y.; Wang, D.; Li, Y.Y.; Gong, M.; Zhang, C. In situ, high-resolution evidence for iron-coupled mobilization of phosphorus in sediments. Nat. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef]
- Malcolm, R.L. The uniqueness of humic substances in each of soil, stream and marine environments. Anal. Chim. Acta 1990, 232, 19–30. [Google Scholar] [CrossRef]
- Reitzel, K.; Ahlgren, J.; Gogoll, A.; Jensen, H.S.; Rydin, E. Characterization of phosphorus in sequential extracts from lake sediments using 31 P nuclear magnetic resonance spectroscopy. Can. J. Fish. Aquat. Sci. 2006, 63, 1686–1699. [Google Scholar] [CrossRef]
- Chambers, R.; Odum, W. Porewater oxidation, dissolved phosphate and the iron curtain. Biogeochemistry 1990, 10, 37–52. [Google Scholar] [CrossRef]
- Pacini, N.; Gächter, R. Speciation of riverine particulate phosphorus during rain events. Biogeochemistry 1999, 47, 87–109. [Google Scholar] [CrossRef]
- McGowan, S.; Britton, G.; Haworth, E.; Moss, B. Ancient blue-green blooms. Limnol. Oceanogr. 1999, 44, 436–439. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| In (m3/yr) | |
| Groundwater input | 450,000 |
| Precipitation | 410,000 |
| Surface water input | 110,000 |
| Out (m3/yr) | |
| Lake evaporation | 550,000 |
| Outlet | 480,000 |
| Storage change (m3/yr) | −60,000 |
| Lake hydraulic retention time (yr) | 1.6 |
| kg P/yr | |
|---|---|
| External TDP loads | |
| Groundwater a | 84 |
| Surface water a | 9 |
| Atmospheric deposition b | 4 |
| Total external TDP load | 97 |
| Internal P release load | |
| Pseasonal_released c | 680 |
| Plongterm_released d | 40 |
| Total internal P load | 720 |
| TDP Output | 126 |
| Net P increase in the water column during summer | 650 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nisbeth, C.S.; Jessen, S.; Bennike, O.; Kidmose, J.; Reitzel, K. Role of Groundwater-Borne Geogenic Phosphorus for the Internal P Release in Shallow Lakes. Water 2019, 11, 1783. https://doi.org/10.3390/w11091783
Nisbeth CS, Jessen S, Bennike O, Kidmose J, Reitzel K. Role of Groundwater-Borne Geogenic Phosphorus for the Internal P Release in Shallow Lakes. Water. 2019; 11(9):1783. https://doi.org/10.3390/w11091783
Chicago/Turabian StyleNisbeth, Catharina S., Søren Jessen, Ole Bennike, Jacob Kidmose, and Kasper Reitzel. 2019. "Role of Groundwater-Borne Geogenic Phosphorus for the Internal P Release in Shallow Lakes" Water 11, no. 9: 1783. https://doi.org/10.3390/w11091783
APA StyleNisbeth, C. S., Jessen, S., Bennike, O., Kidmose, J., & Reitzel, K. (2019). Role of Groundwater-Borne Geogenic Phosphorus for the Internal P Release in Shallow Lakes. Water, 11(9), 1783. https://doi.org/10.3390/w11091783