Effect of Different Copper Levels on Growth and Morpho-Physiological Parameters in Giant Reed (Arundo donax L.) in Semi-Hydroponic Mesocosm Experiment

 ,
, 
 ,
,  ,
,  and
and
Abstract
:1. Introduction
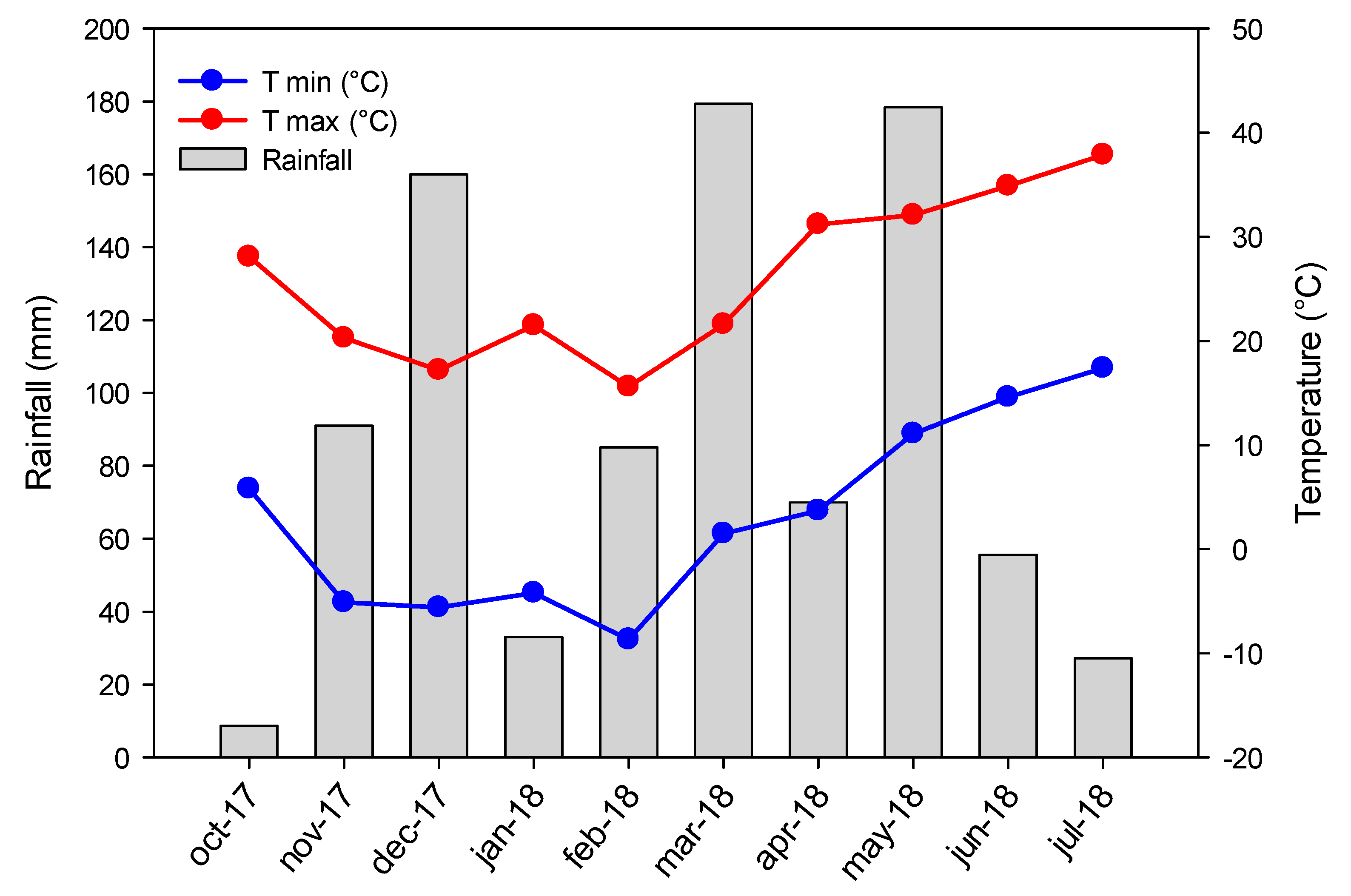
2. Materials and Methods
2.1. Plant Growth and Contamination
2.2. Morphological Characterization of the Plants
2.3. Nutritional Characterization of the Plants
2.4. Determination of Physiological Parameters
2.5. Statistical Analysis
3. Results and Discussion
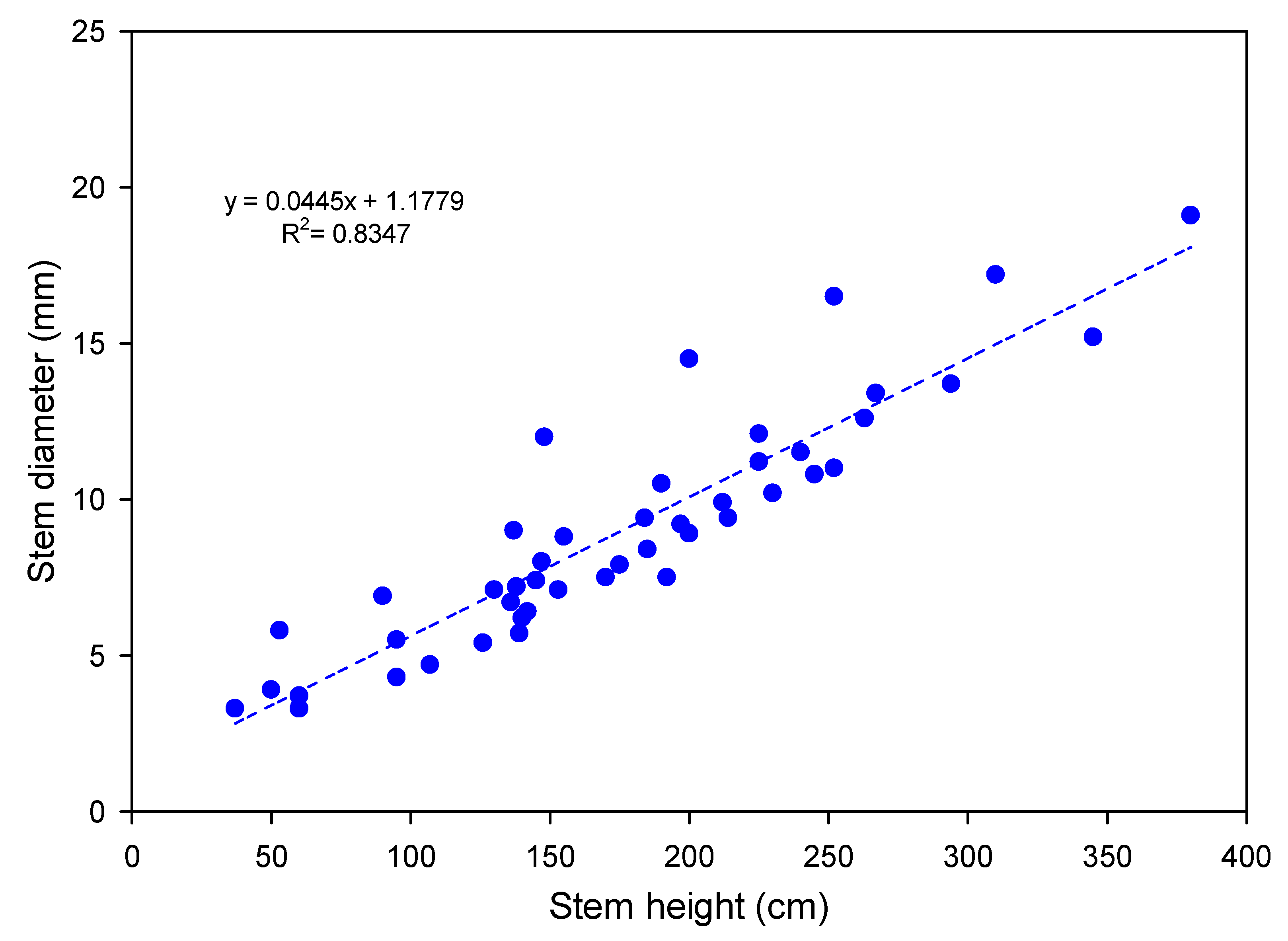
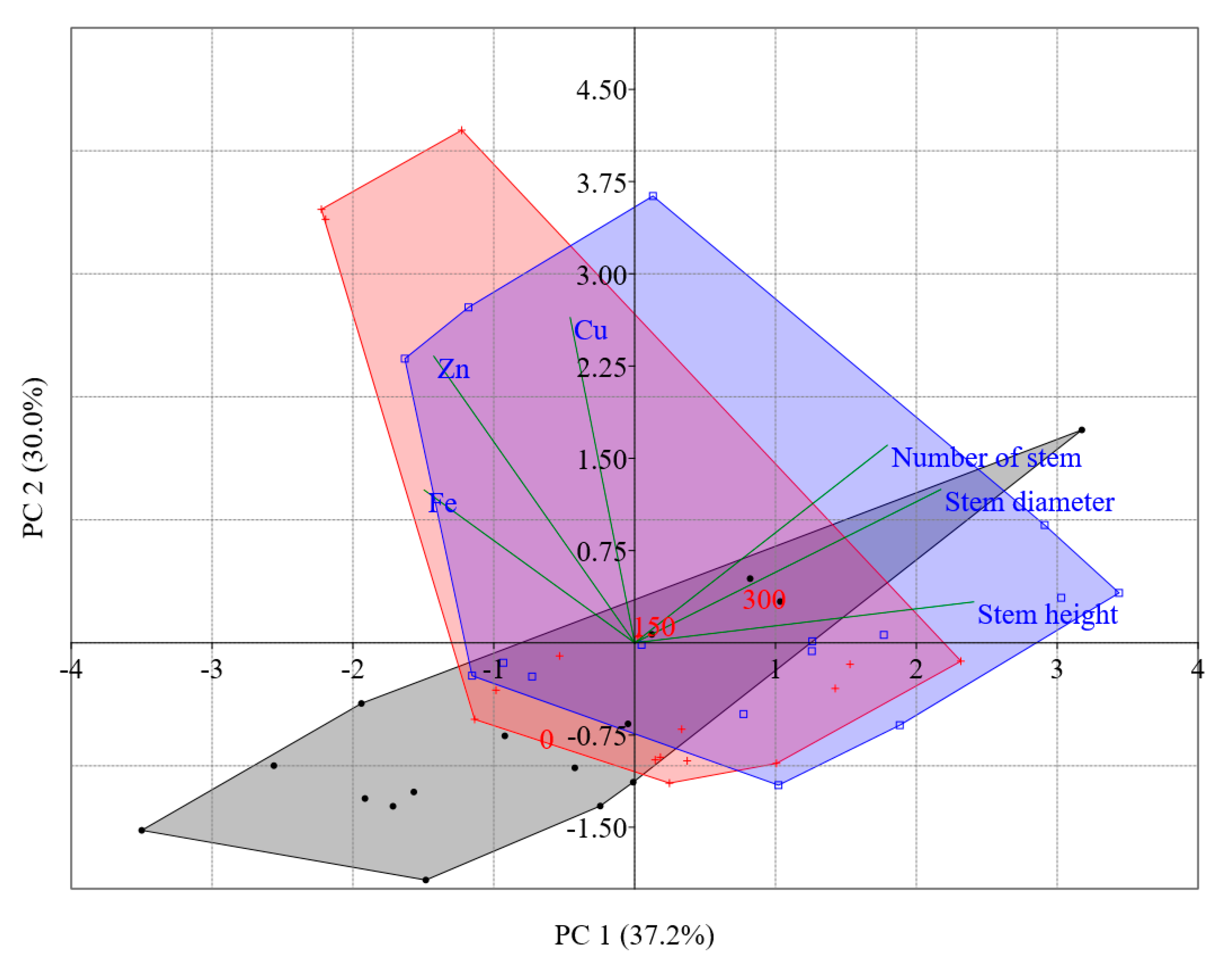
3.1. Morphological Characterization
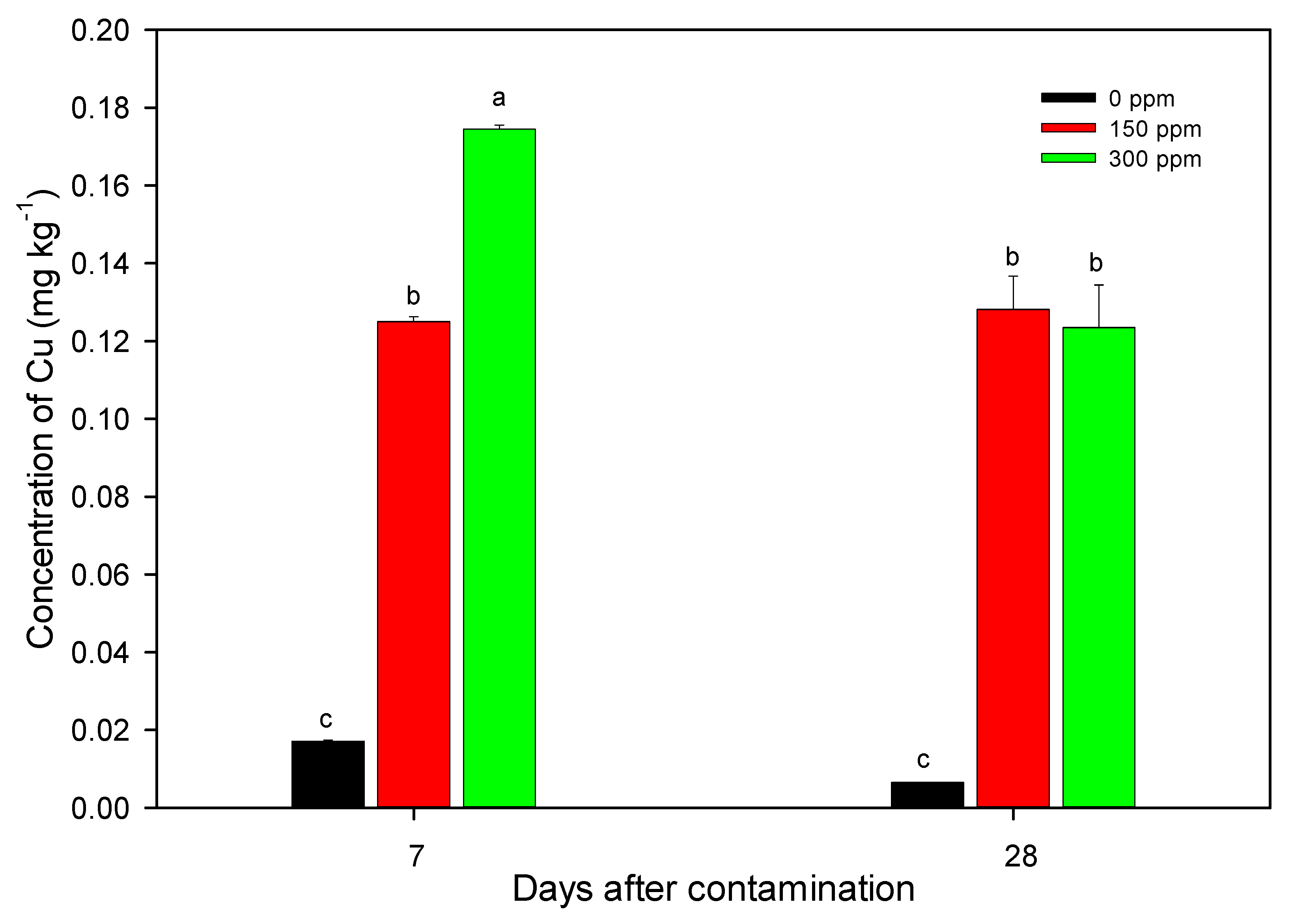
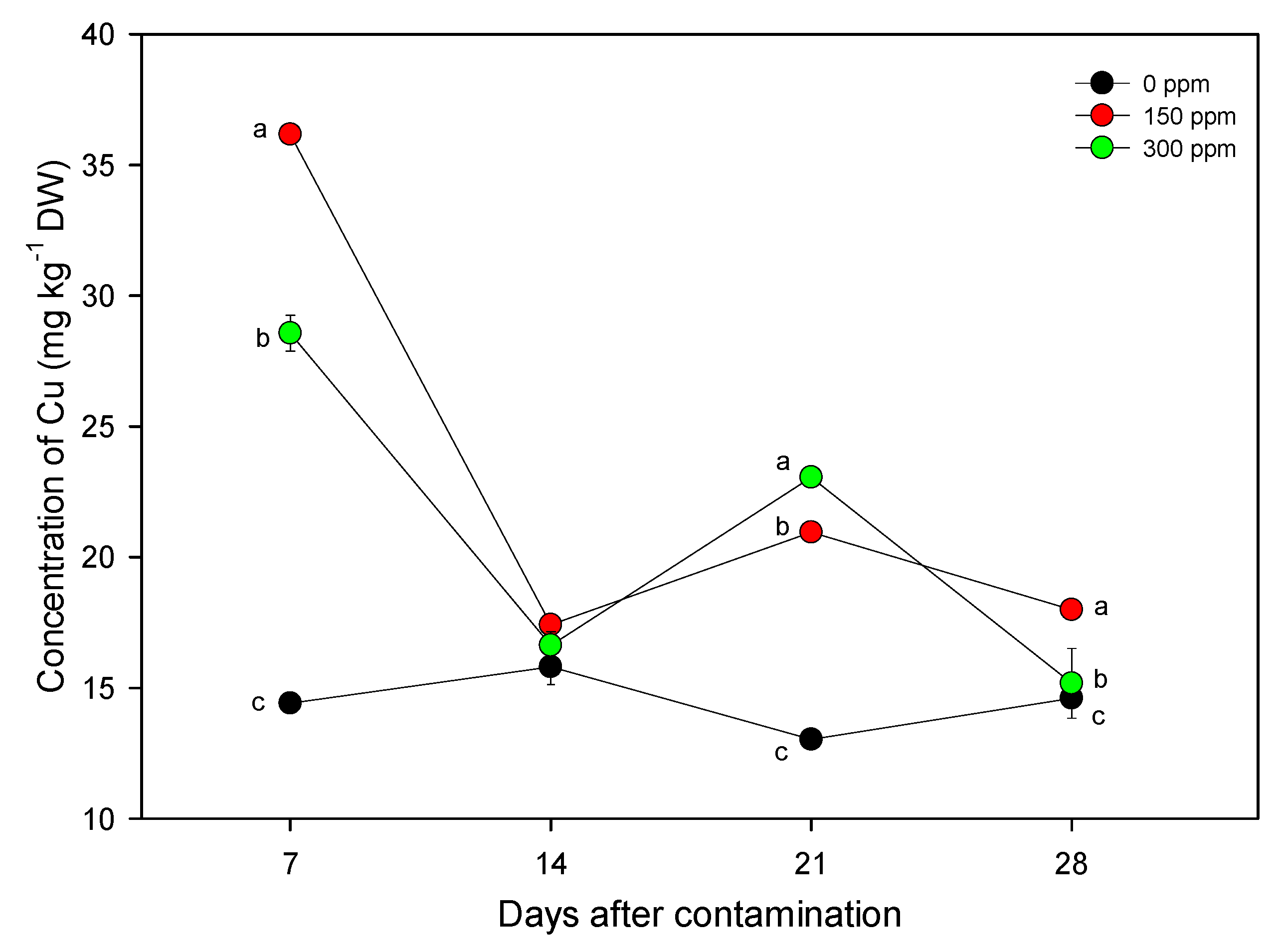
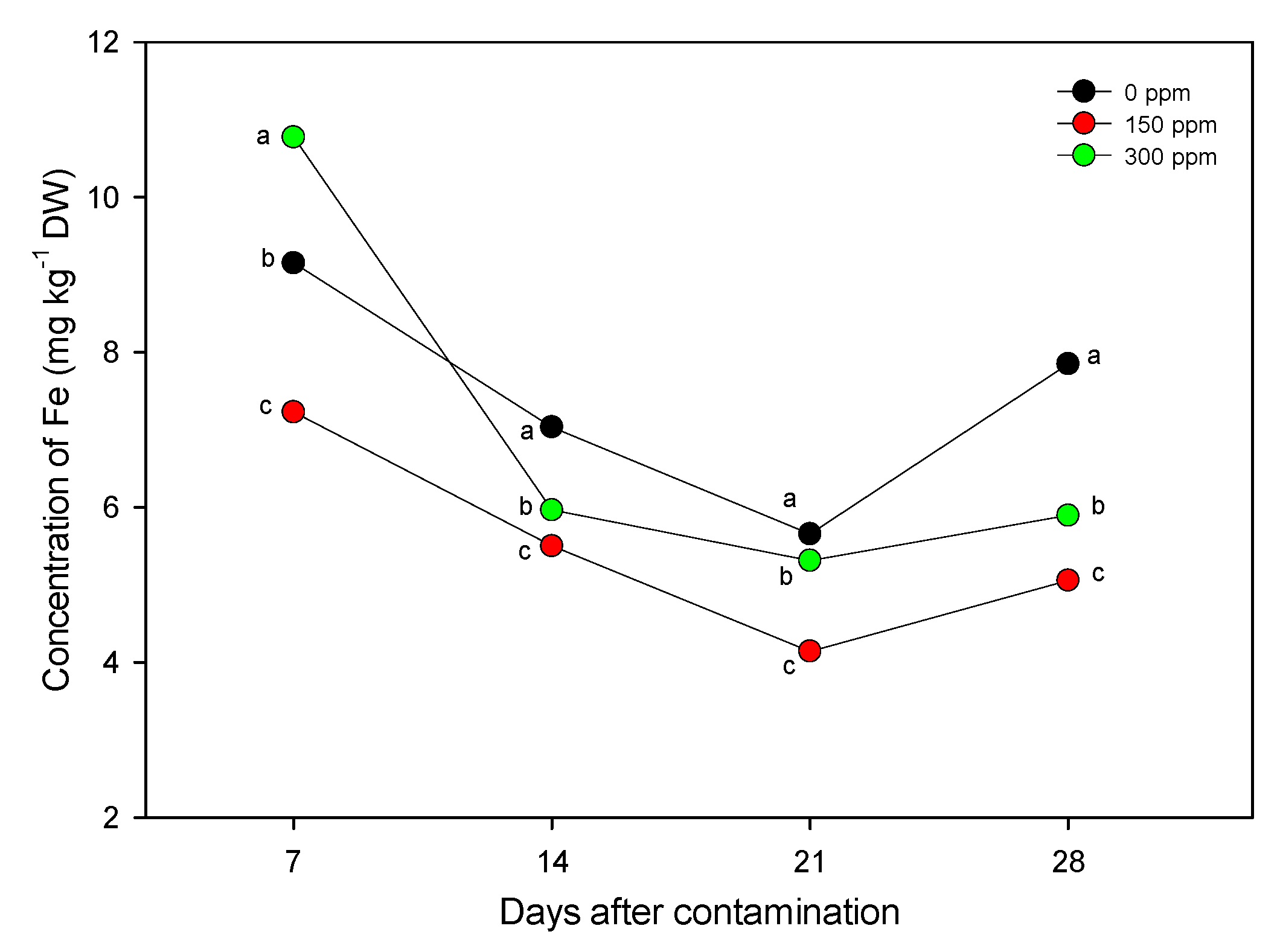
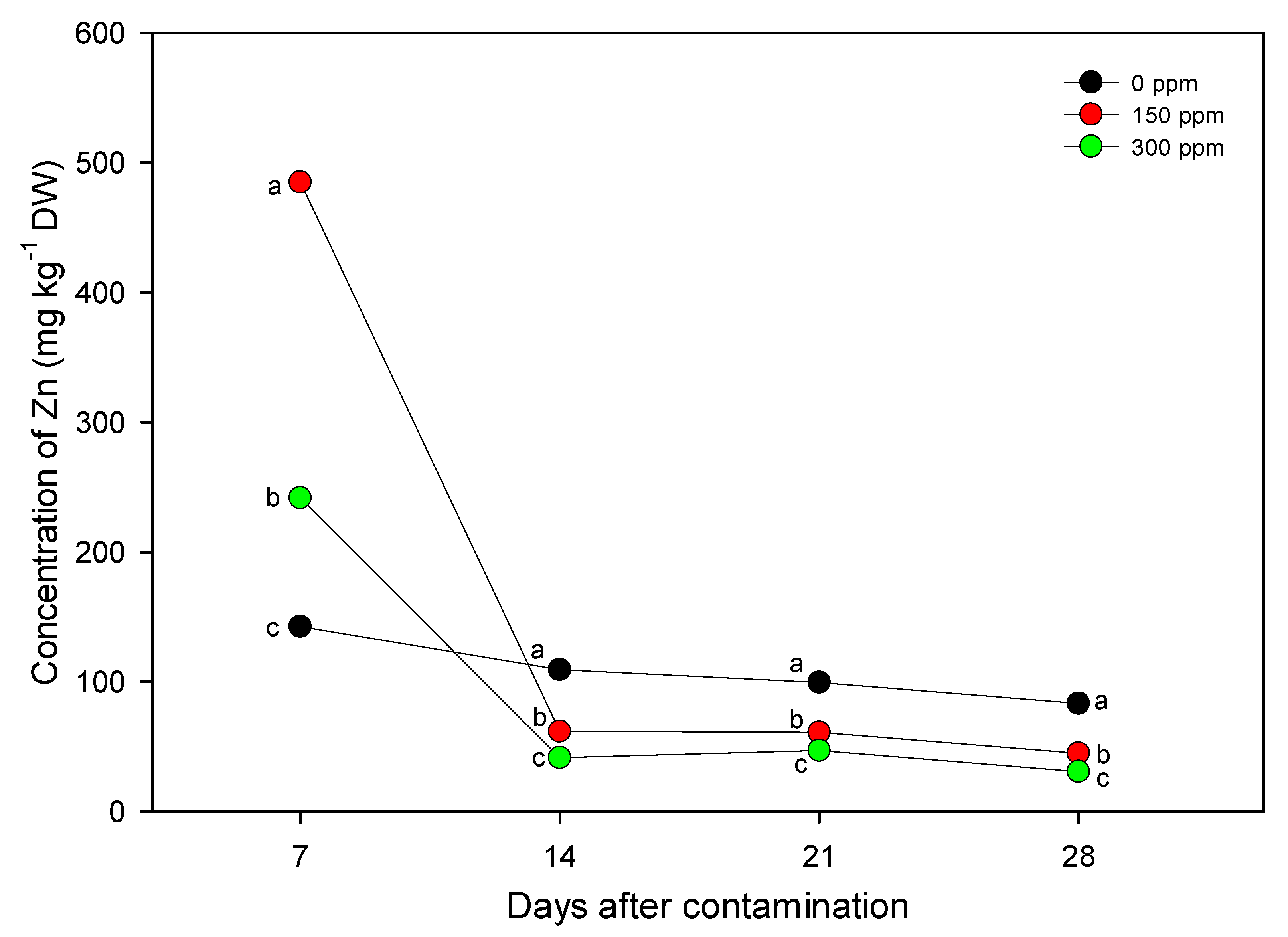
3.2. Nutritional Characterization
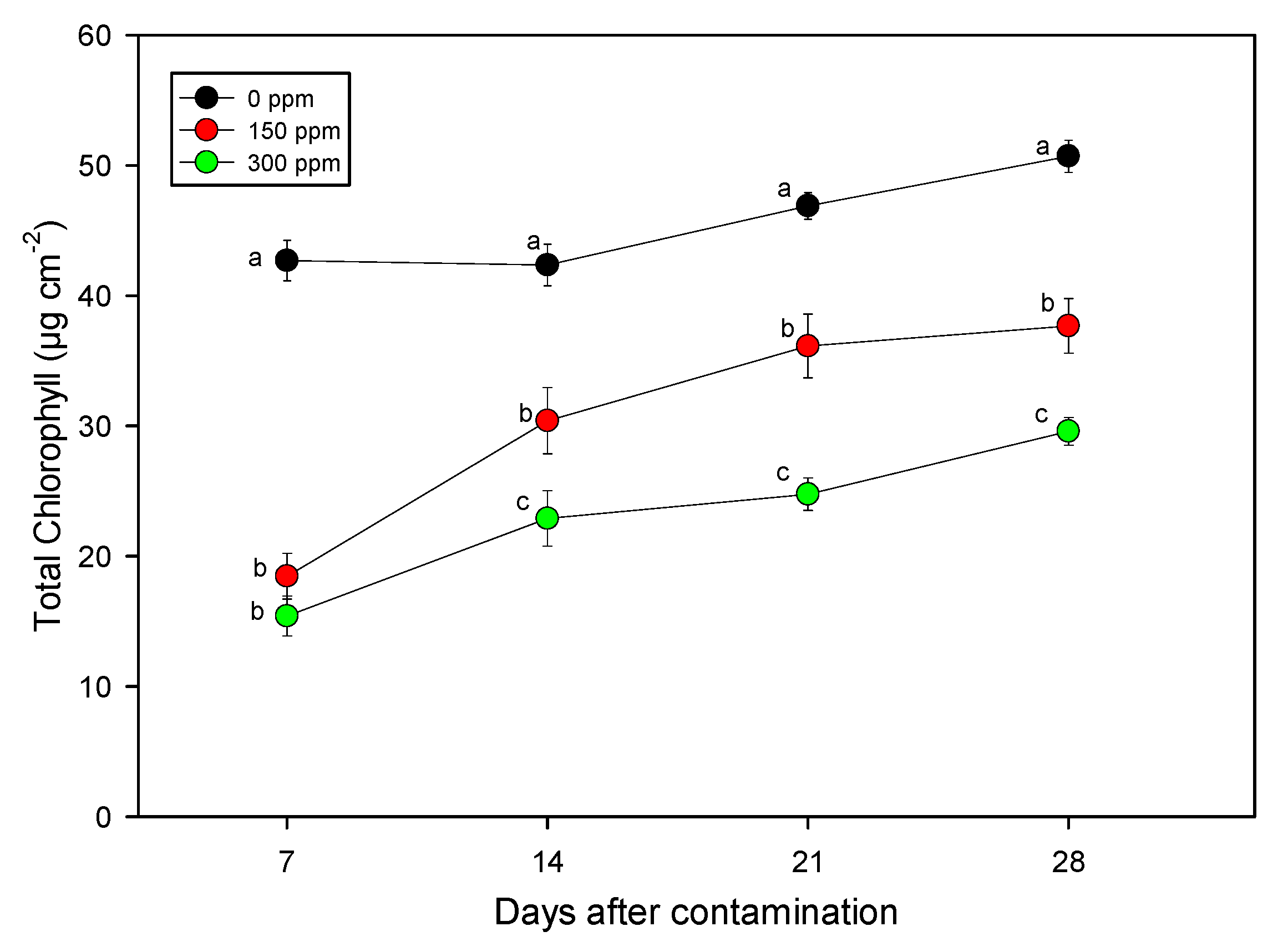
3.3. Determination of Physiological Parameters
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cicchella, D.; Giaccio, L.; Dinelli, E.; Albanese, S.; Lima, A.; Zuzolo, D.; Valera, P.; De Vivo, B. GEMAS: Spatial distribution of chemical elements in agricultural and grazing land soil of Italy. J. Geochem. Explor. 2015, 154, 129–142. [Google Scholar] [CrossRef]
- Toth, G.; Hermann, T.; Da Silva, M.; Montanarella, L. Heavy metals in agricultural soils of the European Union with implications for food safety. Environ. Int. 2016, 88, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Toth, G.; Hermann, T.; Szatmari, G.; Pasztor, L. Maps of heavy metals in the soils of the European Union and proposed priority areas for detailed assessment. Sci. Total Environ. 2016, 565, 1054–1062. [Google Scholar] [CrossRef] [PubMed]
- Marchiol, L.; Fellet, G. Agronomy towards the Green Economy. Optimization of metal phytoextraction. Ital. J. Agron. 2011, 6, 189–197. [Google Scholar] [CrossRef]
- Miotto, A.; Ceretta, C.A.; Brunetto, G.; Nicoloso, F.T.; Girotto, E.; Farias, J.G.; Tiecher, T.L.; De Conti, L.; Trentin, G. Copper uptake, accumulation and physiological changes in adult grapevines in response to excess copper in soil. Plant Soil 2014, 374, 593–610. [Google Scholar] [CrossRef]
- Armentano, G. Rame, si va verso una riduzione delle dosi annue. Inf. Agrar. 2017, 25, 8. [Google Scholar]
- Borghi, M.; Tognetti, R.; Monteforti, G.; Sebastiani, L. Responses of two poplar species (Populus alba and Populus x canadensis) to high copper concentrations. Environ. Exp. Bot. 2008, 62, 290–299. [Google Scholar] [CrossRef]
- Cuypers, A.; Vangronsveld, J.; Clijsters, H. Biphasic effect of copper on the ascorbate-glutathione pathway in primary leaves of Phaseolus vulgaris seedlings during the early stages of metal assimilation. Physiol. Plant. 2000, 110, 512–517. [Google Scholar] [CrossRef]
- La Torre, A.; Iovino, V.; Caradonia, F. Copper in plant protection: Current situation and prospects. Phytopathol. Mediterr. 2018, 57, 201–236. [Google Scholar]
- Ambrosini, V.G.; Rosa, D.J.; Bastos de Melo, G.W.; Zalamena, J.; Cella, C.; Simão, D.G.; Souza da Silva, L.; Pessoa dos Santos, H.; Toselli, M.; Tiecher, T.L.; et al. High copper content in vineyard soils promotes modifications in photosynthetic parameters and morphological changes in the root system of ‘Red Niagara’ plantlets. Plant Physiol. Biochem. 2018, 128, 89–98. [Google Scholar] [CrossRef]
- Pietrini, F.; Di Baccio, D.; Iori, V.; Veliksar, S.; Lemanova, N.; Juškaitė, L.; Maruška, A.; Zacchini, M. Investigation on metal tolerance and phytoremoval activity in the poplar hybrid clone “Monviso” under Cu-spiked water: Potential use for wastewater treatment. Sci. Total Environ. 2017, 592, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Poschenrieder, C.; Bech, J.; Llugany, M.; Pace, A.; Fenés, E.; Barcelò, J. Copper in plant species in a copper gradient in Catalonia (North East Spain) and their potential for phytoremediation. Plant Soil 2001, 230, 247–256. [Google Scholar] [CrossRef]
- Zacchini, M.; Pietrini, F.; Mugnozza, G.S.; Iori, V.; Pietrosanti, L.; Massacci, A. Metal Tolerance, Accumulation and Translocation in Poplar and Willow Clones Treated with Cadmium in Hydroponics. Water Air Soil Pollut. 2009, 197, 23–34. [Google Scholar] [CrossRef]
- Pietrini, F.; Iori, V.; Pietropaoli, S.; Mughini, G.; Beni, C.; Massacci, A.; Zacchini, M. Phytoremediation of cadmium polluted waters by a eucalypt hybrid clone: A mesocosm study. In Proceedings of the Sixth European Bioremediation Conference, Chania, Creta, Greece, 29 June–2 July 2015; pp. 238–241. [Google Scholar]
- Barbosa, B.; Boléo, S.; Sidella, S.; Costa, J.; Duarte, M.P.; Mendes, B.; Cosentino, S.L.; Fernando, A.L. Phytoremediation of heavy metal-contaminated soils using the perennial energy crops Miscanthus spp. and Arundo donax L. Bioenergy Res. 2015, 8, 1500–1511. [Google Scholar] [CrossRef]
- Fiorentino, N.; Ventorino, V.; Rocco, C.; Cenvinzo, V.; Agrelli, D.; Gioia, L.; Di Mola, I.; Adamo, P.; Pepe, O.; Fagnano, M. Giant reed growth and effects on soil biological fertility in assisted phytoremediation of an industrial polluted soil. Sci. Total Environ. 2017, 575, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Nsanganwimana, F.; Marchand, L.; Douay, F.; Mench, M. Arundo donax L., a Candidate for Phytomanaging Water and Soils Contaminated by Trace Elements and Producing Plant-Based Feedstock. A Review. Int. J. Phytoremediat. 2014, 16, 982–1017. [Google Scholar] [CrossRef] [PubMed]
- Oustriere, N.; Marchand, L.; Lottier, N.; Motelica, M.; Mench, M. Long-term Cu stabilization and biomass yields of Giant reed and poplar after adding a biochar, alone or with iron grit, into a contaminated soil from a wood preservation site. Sci. Total Environ. 2017, 579, 620–627. [Google Scholar] [CrossRef]
- Elhawat, N.; Alshaal, T.; Domokos-Szabolcsy, E.; El-Ramady, H.; Antal, G.; Márton, L.; Czakó, M.; Balogh, P.; Fári, M. Copper uptake efficiency and its distribution within bioenergy grass giant reed. Bull. Environ. Contam. Toxicol. 2015, 95, 452–458. [Google Scholar] [CrossRef]
- Marchiol, L. Prospettive e limitazioni del fitorisanamento. Not. ERSA 2008, 4, 65–72. [Google Scholar]
- Ceotto, E.; Di Candilo, M. Shoot cuttings propagation of giant reed (Arundo donax L.) in water and moist soil: The path forward? Biomass Bioenergy 2010, 34, 1614–1623. [Google Scholar] [CrossRef]
- Ruyters, S.; Salaets, P.; Oorts, K.; Smolders, E. Copper toxicity in soils under established vineyards in Europe: A survey. Sci. Total Environ. 2013, 443, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Cerovic, Z.G.; Masdoumier, G.; Ghozlen, N.B.; Latouche, G. A new optical leaf-clip meter for simultaneous non-destructive assessment of leaf chlorophyll and epidermal flavonoids. Physiol. Plant. 2012, 146, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the Chlorophyll a Fluorescence Transient. In Chlorophyll Fluorescence: A Signature of Photosynthesis. Advances in Photosynthesis and Respiration; Papageorgiou, G., Govindjee, Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R. Photosynthetic acclimation and nitrogen partitioning within a lucerne canopy. 2. Stability through time and comparison with a theoretical optimum. Aust. J. Plant Physiol. 1993, 20, 69–82. [Google Scholar] [CrossRef]
- Adrees, M.; Ali, S.; Rizwan, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Zia-ur-Rehman, M.; Irshad, M.K.; Bharwana, S.A. The effect of excess copper on growth and physiology of important food crops: A review. Environ. Sci. Pollut. Res. 2015, 22, 8148–8162. [Google Scholar] [CrossRef]
- Oustriere, N.; Marchand, L.; Roulet, E.; Mench, M. Rhizofiltration of a Bordeaux mixture effluent in pilot-scale constructed wetland using Arundo donax L. coupled with potential Cu-ecocatalyst production. Ecol. Eng. 2017, 105, 296–305. [Google Scholar] [CrossRef]
- Yruela, I. Copper in plants: Acquisition, transport and interactions. Funct. Plant Biol. 2009, 36, 409–430. [Google Scholar] [CrossRef]
- Elhawat, N.; Alshaal, T.; Domokos-Szabolcsy, É.; El-Ramady, H.; Márton, L.; Czakó, M.; Kátai, J.; Balogh, P.; Sztrik, A.; Molnár, M.; et al. Phytoaccumulation potentials of two biotechnologically propagated ecotypes of Arundo donax in copper-contaminated synthetic wastewater. Environ. Sci. Pollut. Res. 2014, 21, 7773–7780. [Google Scholar] [CrossRef]
- Angelini, L.G.; Ceccarini, L.; Nassi o Di Nasso, N.; Bonari, E. Comparison of Arundo donax L. and Miscanthus x giganteus in a long-term field experiment in Central Italy: Analysis of productive characteristics and energy balance. Biomass Bioenergy 2009, 33, 635–643. [Google Scholar] [CrossRef]
- Li, C.; Xiao, B.; Wang, Q.H.; Yao, S.H.; Wu, J.Y. Phytoremediation of Zn- and Cr-contaminated soil using two promising energy grasses. Water Air Soil Pollut. 2014, 225, 2027. [Google Scholar] [CrossRef]
- Mathialagan, T.; Viraraghavan, T. Adsorption of cadmium from aqueous solutions by perlite. J. Hazard. Mater. 2002, 94, 291–303. [Google Scholar] [CrossRef]
- Vijayaraghavan, K.; Raja, F.D. Experimental characterisation and evaluation of perlite as a sorbent for heavy metal ions in single and quaternary solutions. J. Water Process. Eng. 2014, 4, 179–184. [Google Scholar] [CrossRef]
- Ghassabzadeh, H.; Mohadespour, A.; Torab-Mostaedi, M.; Zaheri, P.; Maragheh, M.G.; Taheri, H. Adsorption of Ag, Cu and Hg from aqueous solutions using expanded perlite. J. Hazard. Mater. 2010, 177, 950–955. [Google Scholar] [CrossRef] [PubMed]
- Sari, A.; Tuzen, M.; Citak, D.; Soylak, M. Adsorption characteristics of Cu(II) and Pb(II) onto expanded perlite from aqueous solution. J. Hazard. Mater. 2007, 148, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Pedescoll, A.; Sidrach-Cardona, R.; Hijosa-Valsero, M.; Bécares, E. Design parameters affecting metals removal in horizontal constructed wetlands for domestic wastewater treatment. Ecol. Eng. 2015, 80, 92–99. [Google Scholar] [CrossRef]
- Marchand, L.; Nsanganwimana, F.; Oustrière, N.; Grebenshchykova, Z.; Lizama-Allende, K.; Mench, M. Copper removal from water using a bio-rack system either unplanted or planted with Phragmites australis, Juncus articulatus and Phalaris arundinacea. Ecol. Eng. 2014, 64, 291–300. [Google Scholar] [CrossRef]
- Galletti, A.; Verlicchi, P.; Ranieri, E. Removal and accumulation of Cu, Ni and Zn in horizontal subsurface flow constructed wetlands: Contribution of vegetation and filling medium. Sci. Total Environ. 2010, 408, 5097–5105. [Google Scholar] [CrossRef]
- Rehman, M.; Maqbool, Z.; Peng, D.; Liu, L. Morpho-physiological traits, antioxidant capacity and phytoextraction of copper by ramie (Boehmeria nivea L.) grown as fodder in copper-contaminated soil. Environ. Sci. Pollut. Res. 2019, 26, 5851–5861. [Google Scholar] [CrossRef]
- Brunetto, G.; Bastos de Melo, G.W.; Terzano, R.; Del Buono, D.; Astolfi, S.; Tomasi, N.; Pii, Y.; Mimmo, T.; Cesco, S. Copper accumulation in vineyard soils: Rhizosphere processes and agronomic practices to limit its toxicity. Chemosphere 2016, 162, 293–307. [Google Scholar] [CrossRef] [Green Version]
- Lequeux, H.; Hermans, C.; Lutts, S.; Verbruggen, N. Response to copper excess in Arabidopsis thaliana: Impact on the root system architecture, hormone distribution, lignin accumulation and mineral profile. Plant Physiol. Biochem. 2010, 48, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Azeez, M.O.; Adesanwo, O.O.; Adepetu, J. Effect of Copper (Cu) application on soil available nutrients and uptake. Afr. J. Agric. Res. 2015, 10, 359–364. [Google Scholar] [Green Version]
- Feigl, G.; Kumar, D.; Lehotai, N.; Tugyi, N.; Molnár, Á.; Ördög, A.; Szepesi, Á.; Gémes, K.; Laskay, G.; Erdei, L.; et al. Physiological and morphological responses of the root system of Indian mustard (Brassica juncea L. Czern.) and rapeseed (Brassica napus L.) to copper stress. Ecotoxicol. Environ. Saf. 2013, 94, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Marzilli, M.; Di Santo, P.; Palumbo, G.; Maiuro, L.; Paura, B.; Tognetti, R.; Cocozza, C. Cd and Cu accumulation, translocation and tolerance in Populus alba clone (Villafranca) in autotrophic in vitro screening. Environ. Sci. Pollut. Res. 2018, 25, 10058–10068. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Shen, Z.; Liu, Y.; Wang, L.; Hannaway, D.; Lu, H. Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence, and chlorophyll content of Tetrastigma hemsleyanum Diels et Gilg. Environ. Exp. Bot. 2009, 65, 177–182. [Google Scholar] [CrossRef]
- Cedergreen, N.; Streibig, J.C. Can the choice of endpoint lead to contradictory results of mixture-toxicity experiments? Environ. Toxicol. Chem. 2005, 24, 1676–1683. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Li, C.; Chen, H.; Shao, B. Ecotoxicology and Environmental Safety Cellular response of freshwater green algae to perfluorooctanoic acid toxicity. Ecotoxicol. Environ. Saf. 2013, 88, 103–107. [Google Scholar] [CrossRef]
- Pätsikkä, E.; Kairavuo, M.; Frantisek, S.; Aro, E.; Tyystja, E. Excess Copper Predisposes Photosystem II to Photoinhibition in Vivo by Outcompeting Iron and Causing Decrease in Leaf Chlorophyll. Plant Physiol. 2002, 129, 1359–1367. [Google Scholar] [Green Version]
- Srivastava, S.; Mishra, S.; Tripathi, R.D.; Dwivedi, S.; Gupta, D.K. Copper-induced oxidative stress and responses of antioxidants and phytochelatins in Hydrilla verticillata (L.f.) Royle. Aquat. Toxicol. 2006, 80, 405–415. [Google Scholar] [CrossRef]
- Yruela, I. Copper in plants. Braz. J. Plant Physiol. 2005, 17, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Baker, N.R. Chlorophyll Fluorescence: A Probe of Photosynthesis In Vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bussotti, F.; Desotgiu, R.; Cascio, C.; Pollastrini, M.; Gravano, E.; Gerosa, G.; Marzuoli, R.; Nali, C.; Lorenzini, G.; Salvatori, E.; et al. Ozone stress in woody plants assessed with chlorophyll a fluorescence. A critical reassessment of existing data. Environ. Exp. Bot. 2011, 73, 19–30. [Google Scholar] [CrossRef]
- Gilmore, A.M.; Hazlett, T.L.; Debrunner, P.G. Comparative time-resolved photosystem II chlorophyll a fluorescence analyses reveal distinctive differences between photoinhibitory reaction center damage and xanthophyll cycledependent energy dissipation. Photochem. Photobiol. 1996, 64, 552–563. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.J.; Stirbet, A.D. Heterogeneity of photosystem II probed by the numerically simulated chlorophyll a fluorescence rise (O–J–I–P). Math. Comput. Simul. 1998, 1, 3–9. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The Fluorescence Transient as a Tool to Characterize and Screen Photosynthetic Samples. In Probing Photosynthesis: Mechanisms, Regulation and Adaptation; Mohammad, Y., Pathre, U., Mohanty, P., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 445–483. [Google Scholar]
- Sbihi, K.; Cherifi, O.; El Gharmali, A.; Oudra, B.; Aziz, F. Accumulation and toxicological effects of cadmium, copper and zinc on the growth and photosynthesis of the freshwater diatom Planothidium lanceolatum (Brébisson) Lange-Bertalot: A laboratory study. J. Mater. Environ. Sci. 2012, 3, 497–506. [Google Scholar]
- Cuchiara, C.C.; Silva, I.M.C.; Martinazzo, E.G.; Braga, E.J.B.; Bacarin, M.A.; Peters, J.A. Chlorophyll Fluorescence Transient Analysis in Alternanthera tenella Colla Plants Grown in Nutrient Solution with Different Concentrations of Copper. J. Agric. Sci. 2013, 5, 8–16. [Google Scholar] [CrossRef]
- Cuchiara, C.C.; Silva, I.M.C.; Dalberto, D.S.; Bacarin, M.A.; Peters, J.A. Chlorophyll a fluorescence in sweet potatoes under different copper concentrations. J. Soil Sci. Plant Nutr. 2015, 15, 179–189. [Google Scholar] [CrossRef]
- Perreault, F.; Samadani, M.; Dewez, D. Effect of soluble copper released from copper oxide nanoparticles solubilisation on growth and photosynthetic processes of Lemna gibba L. Nanotoxicology 2016, 8, 374–382. [Google Scholar] [CrossRef]
- Vassilev, A.; Lidon, F.C.; Scotti Campos, P.; Ramalho, J.C.; Barreiro, M.G.; Yordanov, I. Cu-induced changes in chloroplast lipids and photosystem 2 activity in barley plants. Bulg. J. Plant Physiol. 2003, 29, 33–43. [Google Scholar]
- Takahashi, S.; Murata, N. How do environmental stresses accelerate photoinhibition? Trends Plant Sci. 2008, 13, 178–182. [Google Scholar] [CrossRef]
- Wilhelm, C.; Selmar, D. Energy dissipation is an essential mechanism to sustain the viability of plants: The physiological limits of improved photosynthesis. J. Plant Physiol. 2011, 168, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Goh, C.; Ko, S.; Koh, S.; Kim, Y.-Y.; Bae, H.-J. Photosynthesis and Environments: Photoinhibition and Repair Mechanisms in Plants. J. Plant Biol. 2012, 55, 93–101. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
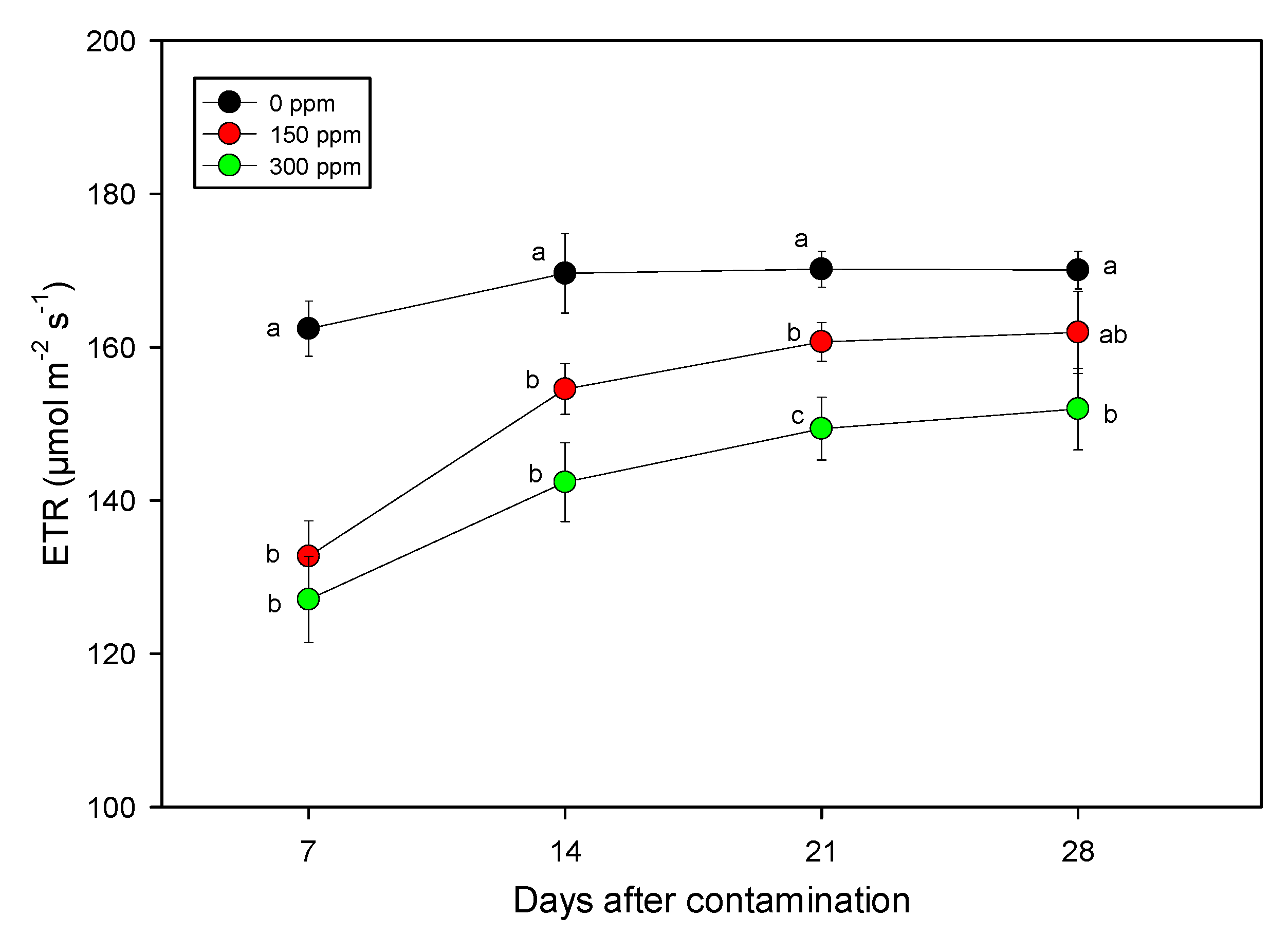{kind=link}
{kind=link}
| Fluorescence Parameters | Description |
|---|---|
| F0 | fluorescence intensity at 50 μs (O step) |
| F300 | fluorescence intensity at 300 μs |
| FJ | fluorescence intensity at 2 ms (J step) |
| FI | fluorescence intensity at 30 ms (I step) |
| Fm | maximal fluorescence intensity (P step) |
| Fv = Fm − F0 | maximal variable fluorescence |
| VJ = (FJ − F0)/(Fm − F0) | variable fluorescence at J step; |
| M0 = 4 (F300 − F0)/(Fm − F0) | approximated initial slope of the fluorescence transient, expressing the rate of RCs’ closure |
| ABS/RC = M0 × (1/VJ) × [1/(Fv/Fm)] | absorption per active reaction center |
| TR0/RC = M0 × (1/VJ) | trapping per active reaction center |
| ET0/RC = M0 × (1/VJ) × (1 − VJ) | electron transport per active reaction center |
| DI0/RC = (ABS/RC) − (TR0/RC) | dissipation per active reaction center |
| TR0/ABS = Fv/Fm = φP0 = (Fm − F0)/Fm | maximum quantum yield of PSII photochemistry |
| ET0/TR0 = ψ0 = (Fm − FJ)/(Fm − F0) | probability that a trapped exciton moves an electron into the electron transport chain beyond QA |
| ET0/ABS = φE0 = φP0 × ψE0 | quantum yield of electron transport |
| Fv/F0 = TR0/DI0 = (Fm − F0)/F0 | maximum ratio of quantum yields of photochemical and concurrent non-photochemical processes in PSII |
| F0/Fm = DI0/ABS = φD0 | maximum quantum yield for energy dissipation at the antenna level |
| PIABS = [φP0 (VJ/M0)] × [φP0/(1 − φP0)] × [ψE0/(1 − ψE0)] | performance index (potential) for energy conservation from photons absorbed by PSII to the reduction of intersystem electron acceptors |
| Variable | Cu Level | Day 0 | Days after Contamination | day28–day0 (%) | |||
|---|---|---|---|---|---|---|---|
| (ppm) | (Contamination Start) | 7 | 14 | 21 | 28 | ||
| Stem height (cm) | 0 | 44.8 ± 9.5 | 56.2 ± 7.2 | 67.1 ± 8.3 | 78.4 ± 12.4 | 110.7 ± 12.9 | 147.0 |
| 150 | 67.5 ± 3.5 | 75.0 ± 2.7 | 95.0 ± 7.3 | 99.2 ± 9.0 | 123.2 ± 9.6 | 82.5 | |
| 300 | 71.5 ± 14.1 | 82.0 ± 11.5 | 100.7 ± 19.0 | 122.0 ± 18.4 | 143.5 ± 15.9 | 100.7 | |
| Basal diameter (mm) | 0 | 5.9 ± 2.4 | 8.9 ± 1.5 | 8.7 ± 0.6 | 11.4 ± 1.3 | 11.5 ± 1.6 | 94.9 |
| 150 | 9.5 ± 1.0 | 10.7 ± 1.1 | 11.0 ± 1.3 | 10.0 ± 0.7 | 11.0 ± 1.7 | 15.8 | |
| 300 | 7.7 ± 1.9 | 10.7 ±0.9 | 13.7 ± 1.7 | 13.0 ± 1.1 | 12.2 ± 1.0 | 58.4 | |
| Number of stems | 0 | 2.9 ± 1.1 | 3.1 ± 0.6 b | 4.3 ± 0.9 | 4.6 ± 1.3 | 5.5 ± 1.4 | 89.7 |
| 150 | 3.5 ± 0.6 | 4.2 ± 0.2 b | 4.0 ± 0.4 | 4.5 ± 0.3 | 4.5 ± 0.3 | 28.6 | |
| 300 | 4.2 ± 2.4 | 6.0 ± 1.1 a | 5.5 ± 0.6 | 4.8 ± 0.7 | 5.3 ± 0.9 | 26.2 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pietrini, F.; Carnevale, M.; Beni, C.; Zacchini, M.; Gallucci, F.; Santangelo, E. Effect of Different Copper Levels on Growth and Morpho-Physiological Parameters in Giant Reed (Arundo donax L.) in Semi-Hydroponic Mesocosm Experiment. Water 2019, 11, 1837. https://doi.org/10.3390/w11091837
Pietrini F, Carnevale M, Beni C, Zacchini M, Gallucci F, Santangelo E. Effect of Different Copper Levels on Growth and Morpho-Physiological Parameters in Giant Reed (Arundo donax L.) in Semi-Hydroponic Mesocosm Experiment. Water. 2019; 11(9):1837. https://doi.org/10.3390/w11091837
Chicago/Turabian StylePietrini, Fabrizio, Monica Carnevale, Claudio Beni, Massimo Zacchini, Francesco Gallucci, and Enrico Santangelo. 2019. "Effect of Different Copper Levels on Growth and Morpho-Physiological Parameters in Giant Reed (Arundo donax L.) in Semi-Hydroponic Mesocosm Experiment" Water 11, no. 9: 1837. https://doi.org/10.3390/w11091837
APA StylePietrini, F., Carnevale, M., Beni, C., Zacchini, M., Gallucci, F., & Santangelo, E. (2019). Effect of Different Copper Levels on Growth and Morpho-Physiological Parameters in Giant Reed (Arundo donax L.) in Semi-Hydroponic Mesocosm Experiment. Water, 11(9), 1837. https://doi.org/10.3390/w11091837