Nutrient Retention in Ecologically Functional Floodplains: A Review


Abstract
:1. Introduction
1.1. Floodplain Processes Driving Nutrient Cycling
1.2. Limitations to the Nutrient Cycle in Floodplains
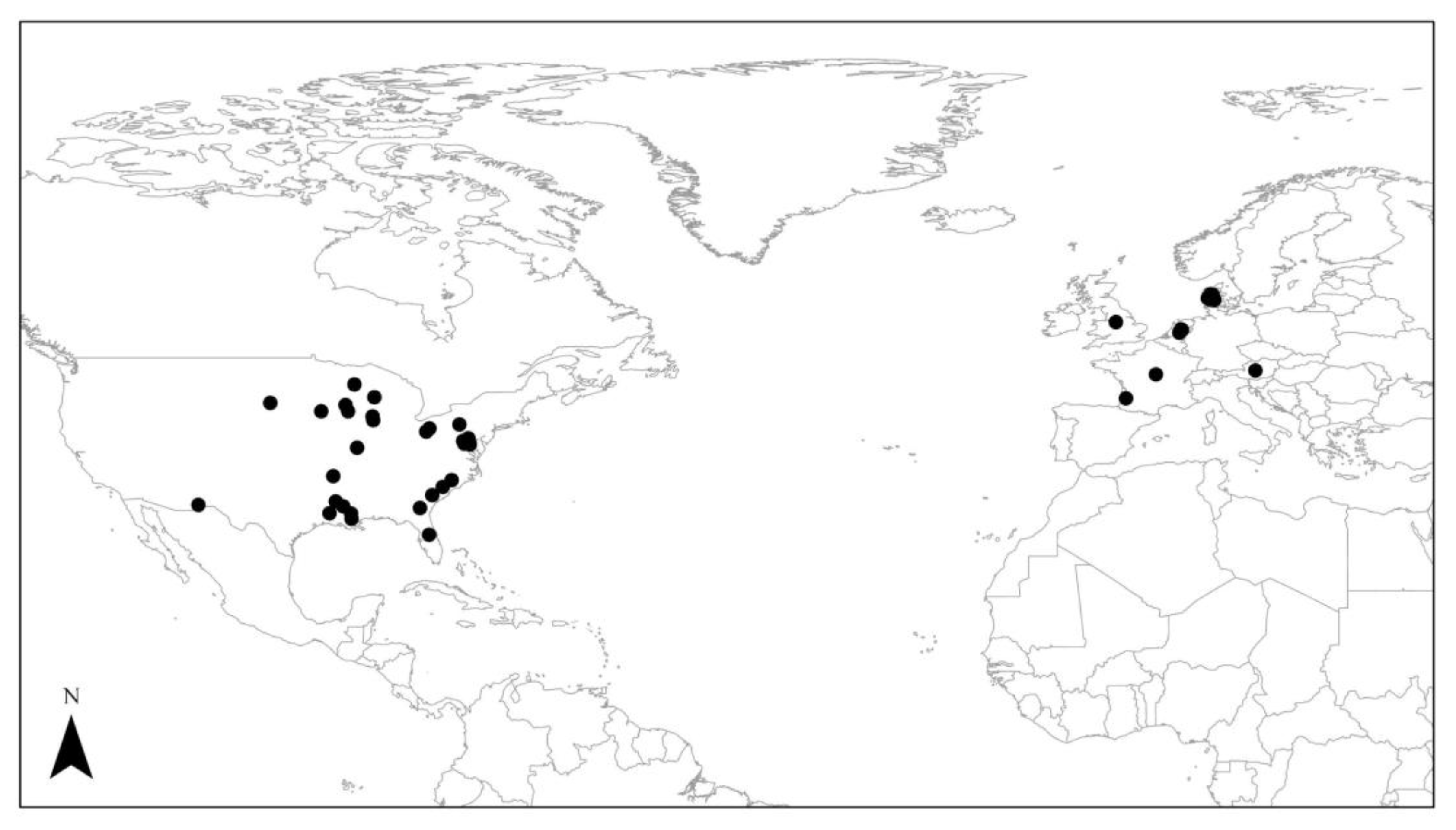
1.3. Selection Criteria for Literature Search
1.4. Evaluating Nutrient Removal from Previous Studies
1.5. Evaluation Considerations
1.5.1. Seasonal Variations and Climate
1.5.2. Nutrient Forms
1.5.3. Size
2. Nutrient Removal Results
3. Discussion
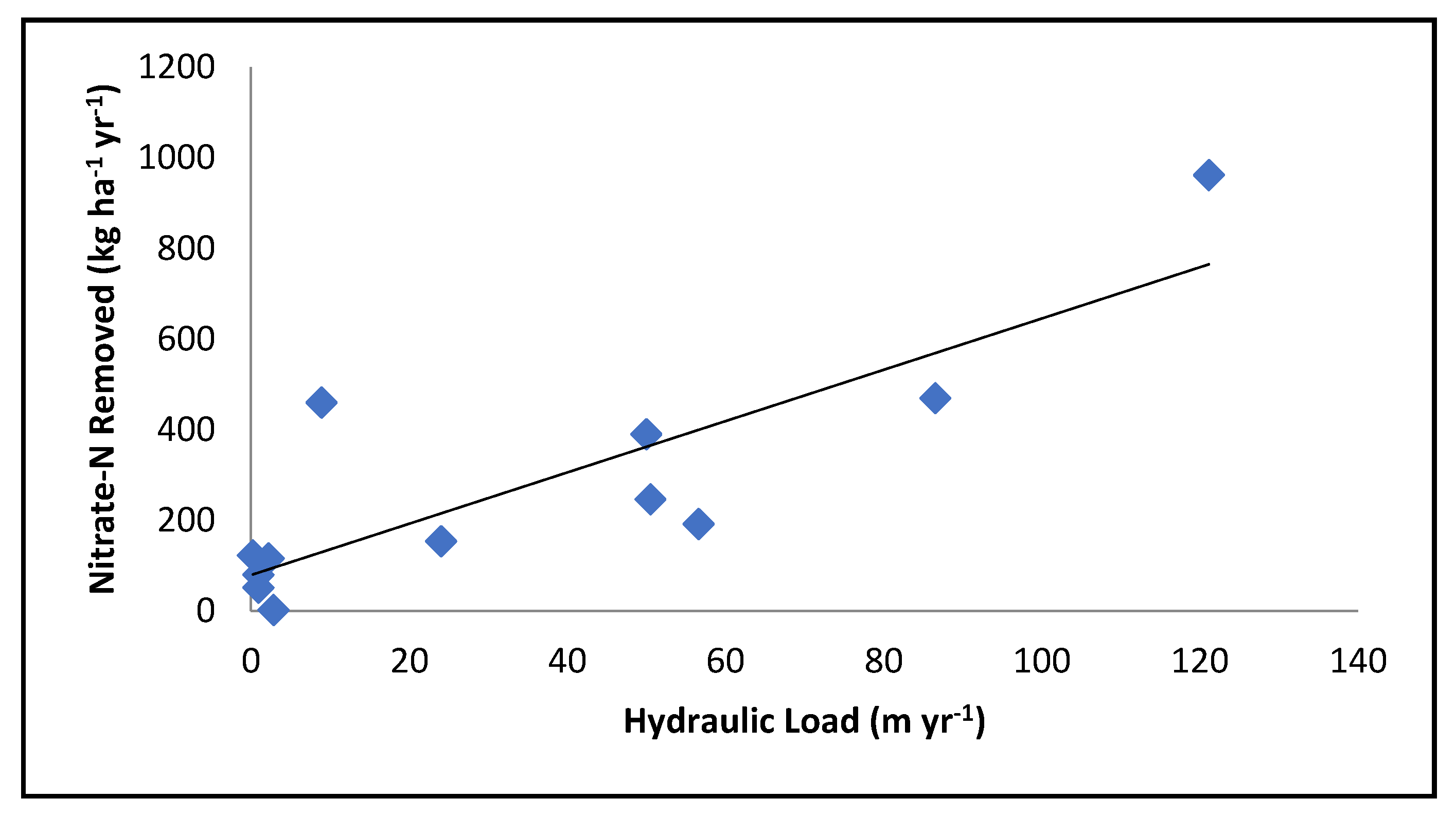
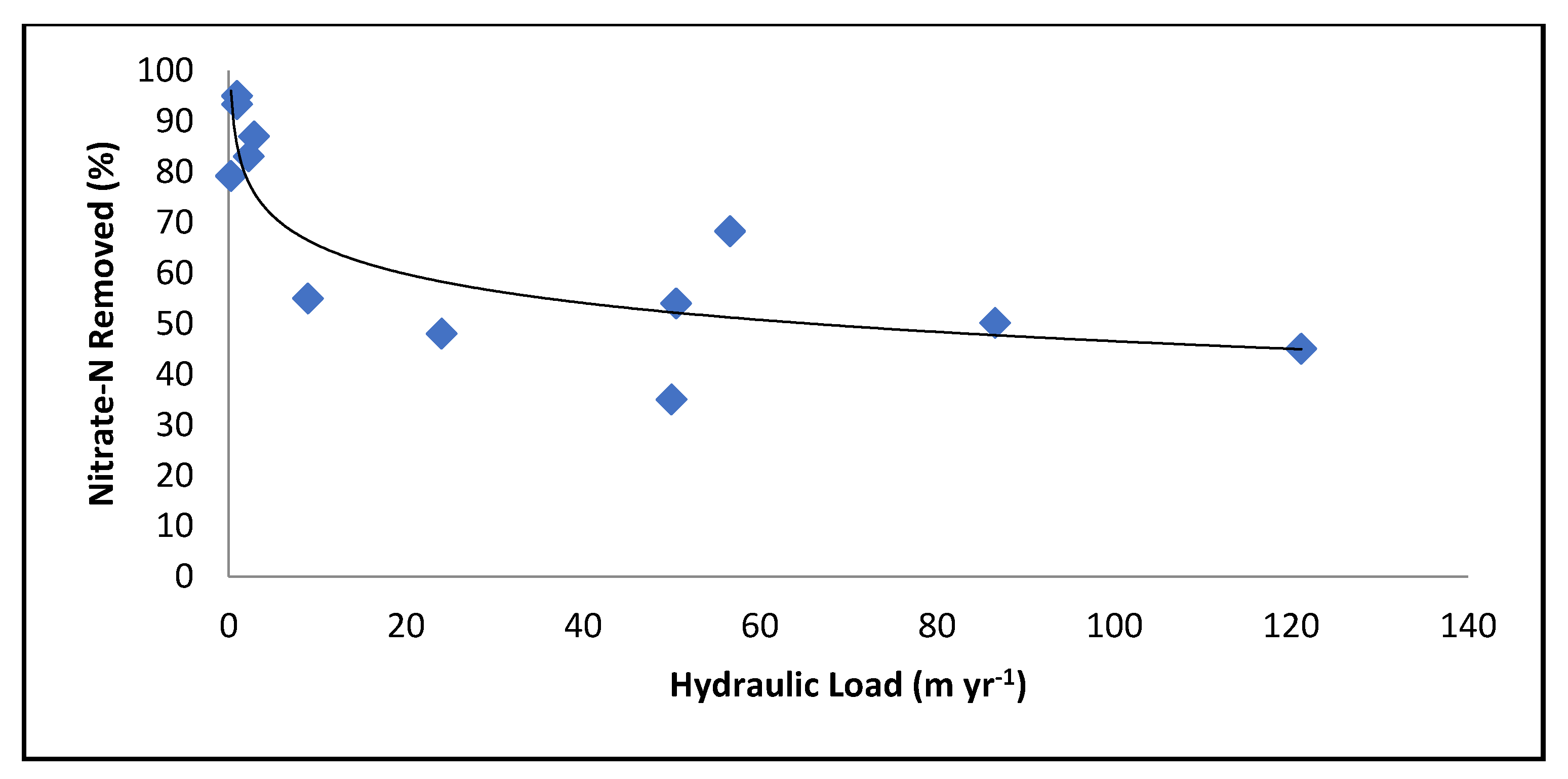
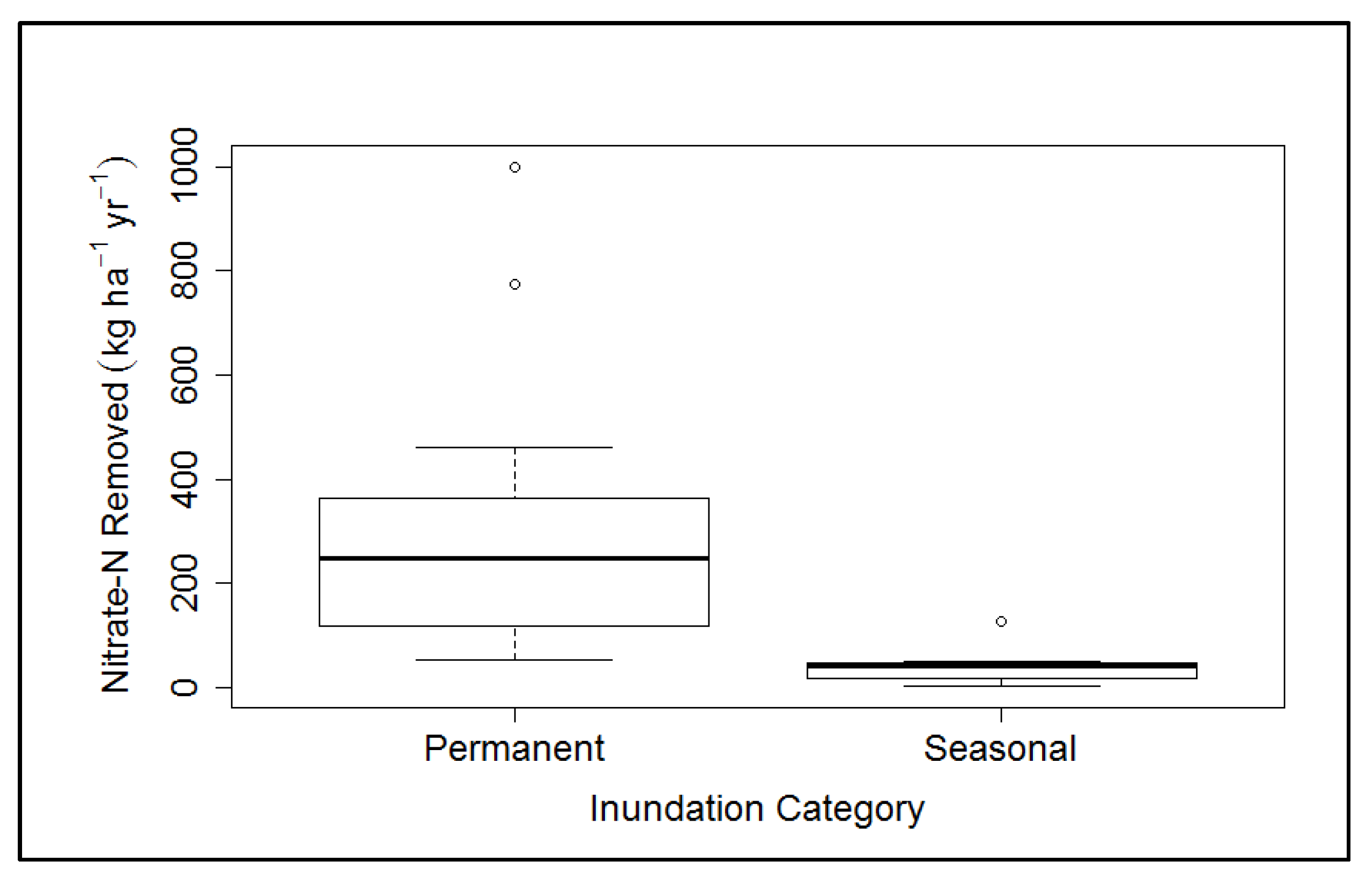
3.1. Hydrology
3.2. Microbes
3.3. Vegetation
3.4. Nutrient Loading
3.5. Nutrient Release from Floodplains
3.6. Future Research
4. Conclusions
- Engineer the floodplain to optimize hydraulic load. Although more flow across the floodplain could lead to a greater total mass of nutrients removed, the floodplain will lose effectiveness, (the percent removal) as flow rates increase.
- Incorporate a permanently inundated wetland in the floodplain area to improve NO3−-N removal.
- Ensure geomorphic diversity across the floodplain to increase both N and P removal due to improved microbial habitat for denitrification and more areas for sedimentation and accretion of P.
- Restore dense vegetation to improve nutrient removal by providing organic matter for denitrifying microbes and slow water flow for better sedimentation and accretion.
- Harvest vegetation from floodplains where feasible to aid in P removal, but caution should be taken due to the unknown impact on native plant communities.
- Restore floodplains along waterways with higher concentrations of nutrients to increase the load of nutrients into the floodplain. Limit flow to maximize nutrient removal.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Glibert, P.M.; Burford, M.A. Globally changing nutrient loads and harmful algal blooms: Recent advances, new paradigms, and continuuing challenges. Oceanography 2017, 30, 58–69. [Google Scholar] [CrossRef] [Green Version]
- Galloway, J.N.; Cowling, E.B. Reactive nitrogen and the world: 200 years of change. AMBIO J. Hum. Environ. 2002, 31, 64–71. [Google Scholar] [CrossRef]
- Howarth, R.W. Coastal nitrogen pollution: A review of sources and trends globally and regionally. Harmful Algae 2008, 8, 14–20. [Google Scholar] [CrossRef]
- Goolsby, D.A.; Battaglin, W.A.; Lawrence, G.B.; Artz, R.S.; Aulenbach, B.T.; Hooper, R.P.; Keeney, D.R.; Stensland, G.J. Flux and Sources of Nutrients in the Mississippi-Atchafalaya River Basin Topic 3 Report for the Integrated Assessment on Hypoxia in the Gulf of Mexico; National Centers for Coastal Ocean Science: Silver Spring, MD, USA, 1999.
- Alexander, R.B.; Smith, R.A.; Schwarz, G.E.; Boyer, E.W.; Nolan, J.V.; Brakebill, J.W. Differences in Phosphorus and Nitrogen Delivery to The Gulf of Mexico from the Mississippi River Basin. Environ. Sci. Technol. 2008, 42, 822–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christianson, L.E.; Harmel, R.D.; Smith, D.; Williams, M.R.; King, K. Assessment and synthesis of 50 years of published drainage phosphorus losses. J. Environ. Qual. 2016, 45. [Google Scholar] [CrossRef] [PubMed]
- King, K.W.; Williams, M.R.; Macrae, M.L.; Fausey, N.R.; Frankenberger, J.; Smith, D.R.; Kleinman, P.J.A.; Brown, L.C. Phosphorus transport in agricultural subsurface drainage: A review. J. Environ. Qual. 2015, 44, 467–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, D.M.; Saad, D.A. SPARROW Models Used to Understand Nutrient Sources in the Mississippi/Atchafalaya River Basin. J. Environ. Qual. 2013, 42, 1422. [Google Scholar] [CrossRef]
- Smith, D.R.; King, K.W.; Johnson, L.; Francesconi, W.; Richards, P.; Baker, D.; Sharpley, A.N. Surface runoff and tile drainage transport of phosphorus in the midwestern United States. J. Environ. Qual. 2015, 44, 495–502. [Google Scholar] [CrossRef]
- Hardison, D.R.; Sunda, W.G.; Litaker, R.W.; Shea, D.; Tester, P.A. Nitrogen limitation increases brevetoxins in Karenia brevis (Dinophyceae): Implications for bloom toxicity. J. Phycol. 2012, 48, 844–858. [Google Scholar] [CrossRef]
- Hardison, D.R.; Sunda, W.G.; Shea, D.; Litaker, R.W. Increased toxicity of Karenia brevis during phosphate limited growth: Ecological and evolutionary implications. PLoS ONE 2013, 8, e58545. [Google Scholar] [CrossRef] [Green Version]
- Granéli, E.; Flynn, K.J. Chemical and physical factors influencing toxin content. In Ecology of Harmful Algae; Granéli, E., Turner, J.T., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 229–241. [Google Scholar]
- Wilhelm, C.; Büchel, C.; Fisahn, J.; Goss, R.; Jakob, T.; LaRoche, J.; Lavaud, J.; Lohr, M.; Riebesell, U.; Stehfest, K.; et al. The regulation of carbon and nutrient assimilation in diatoms is significantly different from green algae. Protist 2006, 157, 91–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glibert, P.M. Margalef revisited: A new phytoplankton mandala incorporating twelve dimensions including nutritional physiology. Harmful Algae 2016, 55, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Ward, M.H.; deKok, T.M.; Levallois, P.; Brender, J.; Gulis, G.; Nolan, B.T.; VanDerslice, J. Workgroup report: Drinking-water nitrate and health-Recent findings and research needs. Environ. Health Perspect. 2005, 113, 1607–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Committee on Environment and Natural Resources. Scientific Assessment of Hypoxia in U.S. Coastal Waters; Committee on Environment and Natural Resources: Washington, DC, USA, 2010.
- Deacon, J.R.; Lee, C.J.; Norman, J.; Reutter, D. Nutrient and Pesticide Data Collected from the USGS National Water Quality Network and Previous Networks, 1963–2016. Available online: https://dx.doi.org/doi:10.5066/F7BK19GZ (accessed on 1 November 2019).
- Hoffmann, C.C.; Kjaergaard, C.; Uusi-Kämppä, J.; Hansen, H.C.B.; Kronvang, B. Phosphorus Retention in Riparian Buffers: Review of Their Efficiency. J. Environ. Qual. 2009, 38, 1942–1955. [Google Scholar] [CrossRef]
- Mayer, P.M.; Reynolds, S.K.; McCutchen, M.D.; Canfield, T.J. Meta-analysis of nitrogen removal in riparian buffers. J. Environ. Qual. 2007, 36, 1172–1180. [Google Scholar] [CrossRef]
- Feyereisen, G.W.; Wilson, B.N.; Sands, G.R.; Strock, J.S.; Porter, P.M. Potential for a rye cover crop to reduce nitrate loss in southwestern Minnesota. Agron. J. 2006, 98, 1416. [Google Scholar] [CrossRef]
- Kaspar, T.C.; Jaynes, D.B.; Parkin, T.B.; Moorman, T.B.; Singer, J.W. Effectiveness of oat and rye cover crops in reducing nitrate losses in drainage water. Agric. Water Manag. 2012, 110, 25–33. [Google Scholar] [CrossRef]
- Gassman, P.W.; Osei, E.; Saleh, A.; Rodecap, J.; Norvell, S.; Williams, J. Alternative practices for sediment and nutrient loss control on livestock farms in northeast Iowa. Agric. Ecosyst. Environ. 2006, 117, 135–144. [Google Scholar] [CrossRef]
- Christianson, L.E.; Harmel, R.D. 4R water quality impacts: An assessment and synthesis of forty years of drainage nitrogen losses. J. Environ. Qual. 2015, 44, 1852–1860. [Google Scholar] [CrossRef]
- Crumpton, W.; Stenback, G.; Miller, B.; Helmers, M. Potential benefits of wetland filters for tile drainage systems: Impact on nitrate loads to Mississippi River subbasins. In Agricultural and Biosystems Engineering Technical Reports White Papers; Iowa State University: Ames, IA, USA, 2006; Volume 8. [Google Scholar]
- Addy, K.; Gold, A.J.; Christianson, L.E.; David, M.B.; Schipper, L.A.; Ratigan, N.A. Denitrifying bioreactors for nitrate removal: A meta-analysis. J. Environ. Qual. 2016, 45, 873–881. [Google Scholar] [CrossRef]
- Sparks, R.E. Need for ecosystem management of large rivers and their floodplains. Bioscience 1995, 45, 168–182. [Google Scholar] [CrossRef]
- Kadlec, R.H.; Wallace, S.D. Treatment Wetlands, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2008; ISBN 9781566705264. [Google Scholar]
- Xue, Y.; David, M.B.; Gentry, L.E.; Mulvaney, R.L.; Kovacic, D.A.; Lindau, C.W. In situ measurements of denitrification in constructed wetlands. J. Environ. Qual. 1999, 28, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Philippot, L.; Hallin, S.; Schloter, M. Ecology of denitrifying prokaryotes in agricultural soil. Adv. Agron. 2007, 96, 249–305. [Google Scholar] [CrossRef]
- Knighton, D. Fluvial Forms and Processes: A New Perspective; Routledge: England, UK, 2014. [Google Scholar]
- Walling, D.E.; He, Q.; Blake, W.H. River flood plains as phosphorus sinks. IAHS Publ. Int. Assoc. Hydrol. Sci. 2000, 263, 211–218. [Google Scholar]
- Jarvie, H.P.; Johnson, L.T.; Sharpley, A.N.; Smith, D.R.; Baker, D.B.; Bruulsema, T.W.; Confesor, R. Increased soluble phosphorus loads to Lake Erie: Unintended consequences of conservation practices? J. Environ. Qual. 2017, 46, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Batjes, N.H. Global Distribution of Soil Phosphorus Retention Potential. In ISRIC-World Soil Inf., No. 2011/0; ISRIC: Wageningen, The Netherlands, 2011. [Google Scholar]
- Nelson, J.C.; Redmond, A.; Sparks, R.E. Impacts of settlement on floodplain vegetation at the confluence of the Illinois and Mississippi Rivers. Trans. Ill. State Acad. Sci. 1994, 87, 117–133. [Google Scholar]
- Llewellyn, D.W.; Shaffer, G.P.; Craig, N.J.; Creasman, L.; Pashley, D.; Mark, S.; Brown, C. A decision-support system for prioritizing restoration sites on the Mississippi River alluvial plain. Conserv. Biol. 1996, 10, 1446–1455. [Google Scholar] [CrossRef]
- Delaney, R.L.; Craig, M.R. Longitudinal Changes in Mississippi River Floodplain Structure; U.S. Geological Survey: Onalaska, WI, USA, 1997.
- Tockner, K.; Stanford, J.A. Riverine flood plains: Present state and future trends. Environ. Conserv. 2002, 29, 308–330. [Google Scholar] [CrossRef] [Green Version]
- Galloway, G.E. Learning from the Mississippi flood of 1993: Impacts, management issues, and areas for research. In Proceedings of the U.S.—Italy Research Workshop on the Hydrometeorology, Impacts, and Management of Extreme Floods, Perugia, Italy, 13–17 November 1995; pp. 1–29. [Google Scholar]
- Magilligan, F.J. Sedimentology of a fine-grained aggrading floodplain. Geomorphology 1992, 4, 393–408. [Google Scholar] [CrossRef]
- Lenhart, C.F.; Titov, M.L.; Ulrich, J.S.; Nieber, J.L.; Suppes, B.J. The role of hydrologic alteration and riparian vegetation dynamics in channel evolution along the lower Minnesota River. Trans. ASABE 2013, 56, 549–561. [Google Scholar] [CrossRef]
- Brady, N.C.; Weil, R.R. The Nature and Properties of Soils; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2007. [Google Scholar]
- Vymazal, J. The use of hybrid constructed wetlands for wastewater treatment with special attention to nitrogen removal: A review of a recent development. Water Res. 2013, 47, 4795–4811. [Google Scholar] [CrossRef] [PubMed]
- Kadlec, R. Large constructed wetlands for phosphorus control: A review. Water 2016, 8, 243. [Google Scholar] [CrossRef] [Green Version]
- Vymazal, J. Removal of nutrients in various types of constructed wetlands. Sci. Total Environ. 2007, 380, 48–65. [Google Scholar] [CrossRef] [PubMed]
- Crumpton, W.; Van Der Valk, A.; Hoyer, W.; Osterberg, D.; Van Der Valk, A.G. Wetland Restoration in Iowa: Challenges and Opportunities; The Iowa Policy Project: Iowa City, IA, USA, 2012. [Google Scholar]
- Rozema, E.R.; VanderZaag, A.C.; Wood, J.D.; Drizo, A.; Zheng, Y.; Madani, A.; Gordon, R.J. Constructed wetlands for agricultural wastewater treatment in northeastern North America: A review. Water 2016, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Newcomer Johnson, T.A.; Kaushal, S.S.; Mayer, P.M.; Smith, R.M.; Sivirichi, G.M. Nutrient retention in restored streams and rivers: A global review and synthesis. Water 2016, 8, 28. [Google Scholar] [CrossRef] [Green Version]
- Kelly, J.M.; Kovar, J.L.; Sokolowsky, R.; Moorman, T.B. Phosphorus uptake during four years by different vegetative cover types in a riparian buffer. Nutr. Cycl. Agroecosyst. 2007, 78, 239–251. [Google Scholar] [CrossRef]
- Koerselman, W.; Bakker, S.A.; Blom, M. Nitrogen, phosphorus and potassium budgets for two small fens surrounded by heavily fertilized pastures. J. Ecol. 1990, 78, 428. [Google Scholar] [CrossRef]
- Hoffmann, C.C.; Dahl, M.; Kamp-Nielsen, L.; Stryhn, H. Vandog Stofbalance i en Natureng; Miljøstyrelsen: Copenhagen, Denmark, 1993. [Google Scholar]
- Venterink, H.O.; Pieterse, N.M.; Belgers, J.D.M.; Wassen, M.J.; de Ruiter, P.C. N, P, and K budgets along nutrient availability and productivity gradients in wetlands. Ecol. Appl. 2002, 12, 1010–1026. [Google Scholar] [CrossRef]
- Fail, J.L.; Haines, B.L.; Todd, R.L. Riparian forest communities and their role in nutrient conservation in an agricultural watershed. Am. J. Altern. Agric. 1987, 2, 114. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing; R Founding for Statistical Computing: Vienna, Austria, 2013.
- Lauer, J.W.; Echterling, C.; Lenhart, C.; Belmont, P.; Rausch, R. Air-photo based change in channel width in the Minnesota River basin: Modes of adjustment and implications for sediment budget. Geomorphology 2017, 297, 170–184. [Google Scholar] [CrossRef]
- Dubbe, D.R.; Garver, E.G.; Pratt, D.C. Production of cattail (Typha spp.) biomass in Minnesota, USA. Biomass 1988, 17, 79–104. [Google Scholar] [CrossRef]
- Grosshans, R.; Grieger, L.; Ackerman, J.; Gauthier, S.; Swystun, K.; Gass, P.; Roy, D. Cattail Biomass in a Watershed-Based Bioeconomy: Commercial-Scale Harvesting and Processing for Nutrient Capture, Biocarbon and High-Value Bioproducts; International Institute for Sustainable Development: Winnipeg, MB, Canada, 2014. [Google Scholar]
- Correll, D.L.; Weller, D.E. Factors limiting processes in freshwater wetlands: An agricultural primary stream riparian forest. In Freshwater Wetlands and Wildlife; Sharitz, R.R., Gibbons, J.W., Eds.; United States Department of Energy: Oak Ridge, TN, USA, 1989. [Google Scholar]
- Peterjohn, W.T.; Correll, D.L. Nutrient dynamics in an agricultural watershed: Observations on the role of a riparian forest. Ecology 1984, 65, 1466–1475. [Google Scholar] [CrossRef]
- Dee, M.M.; Tank, J.L.; Mahl, U.H.; Powers, S.M. Estimating the Impact of Floodplain Restoration on Nutrient and Sediment Export from the Wabash River Watershed: A Historical Perspective; Indiana Water Resources Research Center: Marsteller St., IN, USA, 2014. [Google Scholar]
- USDA NRCS Part 651 agricultural waste management system component design. In National Engineering Handbook; USDA: Washington, DC, USA, 2009; p. 216.
- Argiroff, W.A.; Zak, D.R.; Lanser, C.M.; Wiley, M.J. Microbial community functional potential and composition are shaped by hydrologic connectivity in riverine floodplain soils. Microb. Ecol. 2017, 73, 630–644. [Google Scholar] [CrossRef] [PubMed]
- Tomasek, A.; Staley, C.; Wang, P.; Kaiser, T.; Lurndahl, N.; Kozarek, J.L.; Hondzo, M.; Sadowsky, M.J. Increased denitrification rates associated with shifts in prokaryotic community composition caused by varying hydrologic connectivity. Front. Microbiol. 2017, 8, 2304. [Google Scholar] [CrossRef] [PubMed]
- Ligi, T.; Truu, M.; Truu, J.; Nõlvak, H.; Kaasik, A.; Mitsch, W.J.; Mander, Ü. Effects of soil chemical characteristics and water regime on denitrification genes (nirS, nirK, and nosZ) abundances in a created riverine wetland complex. Ecol. Eng. 2013, 72, 47–55. [Google Scholar] [CrossRef]
- Wang, C.; Zhu, G.; Wang, Y.; Wang, S.; Yin, C. Nitrous oxide reductase gene (nosZ) and N2O reduction along the littoral gradient of a eutrophic freshwater lake. J. Environ. Sci. 2013, 25, 44–52. [Google Scholar] [CrossRef]
- Tomasek, A.; Kozarek, J.L.; Hondzo, M.; Lurndahl, N.; Sadowsky, M.J.; Wang, P.; Staley, C. Environmental drivers of denitrification rates and denitrifying gene abundances in channels and riparian areas. Water Resour. Res. 2017. [Google Scholar] [CrossRef]
- Tomasek, A.A.; Hondzo, M.; Kozarek, J.L.; Staley, C.; Wang, P.; Lurndahl, N.; Sadowsky, M.J. Intermittent flooding of organic-rich soil promotes the formation of denitrification hot moments and hot spots. Ecosphere 2019, 10, e02549. [Google Scholar] [CrossRef]
- Scott, D.T.; Gomez-Velez, J.D.; Jones, C.N.; Harvey, J.W. Floodplain inundation spectrum across the United States. Nat. Commun. 2019, 10, 1–8. [Google Scholar] [CrossRef]
- Hernandez, M.E.; Mitsch, W.J. Denitrification in created riverine wetlands: Influence of hydrology and season. Ecol. Eng. 2007, 30, 78–88. [Google Scholar] [CrossRef]
- Tomasek, A.A.; Barman, T.D.; Wang, P.; Kozarek, J.L.; Staley, C.; Sadowsky, M.J.; Hondzo, M. The effects of turbulence and carbon amendments on nitrate uptake and microbial gene abundances in stream sediment. J. Geophys. Res. Biogeosci. 2018, 123, 1289–1301. [Google Scholar] [CrossRef]
- Brunet, R.C.; Pinay, G.; Gazelle, F.; Roques, L. Role of the floodplain and riparian zone in suspended matter and nitrogen retention in the adour river, south-west France. Regul. Rivers Res. Manag. 1994, 9, 55–63. [Google Scholar] [CrossRef]
- Kronvang, B.; Andersen, I.K.; Hoffmann, C.C.; Pedersen, M.L.; Ovesen, N.B.; Andersen, H.E. Water exchange and deposition of sediment and phosphorus during inundation of natural and restored lowland floodplains. Water. Air. Soil Pollut. 2007, 181, 115–121. [Google Scholar] [CrossRef]
- Noe, G.B.; Hupp, C.R. Carbon, nitrogen, and phosphorus accumulation in floodplains of Atlantic Coastal Plain rivers, USA. Ecol. Appl. 2005, 15, 1178–1190. [Google Scholar] [CrossRef]
- Osborne, L.L.; Kovacic, D.A. Riparian vegetated buffer strips in water-quality restoration and stream management. Freshw. Biol. 1993, 29, 243–258. [Google Scholar] [CrossRef]
- Lowrance, R.; Todd, R.; Fail, J.; Hendrickson, O.; Leonard, R.; Asmussen, L. Riparian forests as nutrient filters in agricultural watersheds. Bioscience 1984, 34, 374–377. [Google Scholar] [CrossRef]
- Jordan, T.; Thomas, E.; Correl, D.; Weller, D. Nutrient interception by a riparian forest receiving inputs from adjacent cropland. J. Environ. Qual. 1993, 22, 467–473. [Google Scholar] [CrossRef] [Green Version]
- Zedler, J.B. Feedbacks that might sustain natural, invaded and restored states in herbaceous wetlands. In New Models for Ecosystem Dynamics and Restoration; Hobbs, R.J., Suding, K.N., Eds.; Society for Ecological Restoration International: Washington, DC, USA, 2009; pp. 236–258. [Google Scholar]
- Lishawa, S.C.; Lawrence, B.A.; Albert, D.A.; Tuchman, N.C. Biomass harvest of invasive Typha promotes plant diversity in a Great Lakes coastal wetland. Restor. Ecol. 2015, 23, 228–237. [Google Scholar] [CrossRef] [Green Version]
- Crumpton, W.; Stenback, G. Iowa Conservation Reserve Enhancement Program. In Annual Performance Report; CREP: London, UK, 2016; Volume 16. [Google Scholar]
- Iovanna, R.; Hyberg, S.; Crumpton, W. Treatment wetlands: Cost-effective practice for intercepting nitrate before it reaches and adversely impacts surface waters. J. Soil Water Conserv. 2008, 63, 14A–15A. [Google Scholar] [CrossRef]
- Hyberg, S.; Iovanna, R.; Crumpton, W.; Richmond, S. The cost effectiveness of wetlands designed and sited for nitrate removal: The effect of increased efficiency, rising easement costs, and lower interest rates. J. Soil Water Conserv. 2015, 70, 30A–32A. [Google Scholar] [CrossRef]
- Kovacic, D.A.; David, M.B.; Gentry, L.E.; Starks, K.M.; Cooke, R.A. Effectiveness of constructed wetlands in reducing nitrogen and phosphorus export from agricultural tile drainage. J. Environ. Qual. 2000, 29, 1262. [Google Scholar] [CrossRef] [Green Version]
- Lenhart, C.; Gordon, B.; Gamble, J.; Current, D.; Ross, N.; Herring, L.; Nieber, J.; Peterson, H. Design and hydrologic performance of a tile drainage treatment wetland in Minnesota, USA. Water 2016, 8, 549. [Google Scholar] [CrossRef] [Green Version]
- Kovacic, D.A.; David, M.B.; Gentry, L.E. Grassed detention buffer strips for reducing agricultural nonpoint-source pollution from tile drainage systems. In Proceedings of the Research on Agricultural Chemicals in Illinois Groundwater: State and Future Directions; Davis, M., Ed.; Southern Illinois University: Carbondale, IL, USA, 1996; pp. 88–97. [Google Scholar]
- Carlson, B.; Vetsch, J.; Randall, G. Nitrates in Drainage Water in Minnesota; University of Minnesota Extension Publication: Minneapolis, MN, USA, 2013. [Google Scholar]
- Hoffmann, C.C.; Kronvang, B.; Audet, J. Evaluation of nutrient retention in four restored Danish riparian wetlands. Hydrobiologia 2011, 674, 5–24. [Google Scholar] [CrossRef]
- Brunet, R.C.; Astin, K.B. A 12-month sediment and nutrient budget in a floodplain reach of the River Adour, southwest France. Regul. Rivers Res. Manag. 2000, 16, 267–277. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Load Reduction | |||||
|---|---|---|---|---|---|
| 25th (kg ha−1 year−1) | 75th (kg ha−1 year−1) | Mean (kg ha−1 year−1) | Median (kg ha−1 year−1) | N | |
| NO3−-N | 77.1 | 260 | 200 | 137 | 28 |
| TP or Particulate P | 2.58 | 22.9 | 21.0 | 8.99 | 41 |
| Load Reduction | |||||
|---|---|---|---|---|---|
| 25th (%) | 75th (%) | Mean (%) | Median (%) | N | |
| NO3−-N | 50.0 | 79.1 | 64.2 | 62.7 | 21 |
| TP or Particulate P | 6.0 | 43.5 | 26.5 | 13.0 | 21 |
| Source | Phosphorus Harvested (kg-P ha−1 year−1) | Plant Community Type |
|---|---|---|
| [18] | 4.5–15 | Reed marshes |
| [48] | 25 | Alfalfa, switchgrass, and cottonwood |
| [49] | 3.9 | Reeds, grasses, and sedges |
| [50] | 11 | Sedge meadow |
| [51] | 3.3–19 | Sedge meadow |
| [52] | 3.8 | Riparian forest |
| [57] | 3–5 | Riparian forest |
| [58] | 1.7 | Riparian forest |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gordon, B.A.; Dorothy, O.; Lenhart, C.F. Nutrient Retention in Ecologically Functional Floodplains: A Review. Water 2020, 12, 2762. https://doi.org/10.3390/w12102762
Gordon BA, Dorothy O, Lenhart CF. Nutrient Retention in Ecologically Functional Floodplains: A Review. Water. 2020; 12(10):2762. https://doi.org/10.3390/w12102762
Chicago/Turabian StyleGordon, Brad A., Olivia Dorothy, and Christian F. Lenhart. 2020. "Nutrient Retention in Ecologically Functional Floodplains: A Review" Water 12, no. 10: 2762. https://doi.org/10.3390/w12102762
APA StyleGordon, B. A., Dorothy, O., & Lenhart, C. F. (2020). Nutrient Retention in Ecologically Functional Floodplains: A Review. Water, 12(10), 2762. https://doi.org/10.3390/w12102762