Complex Undisturbed Riparian Zones Are Resistant to Colonisation by Invasive Alien Plant Species



Abstract
:1. Introduction
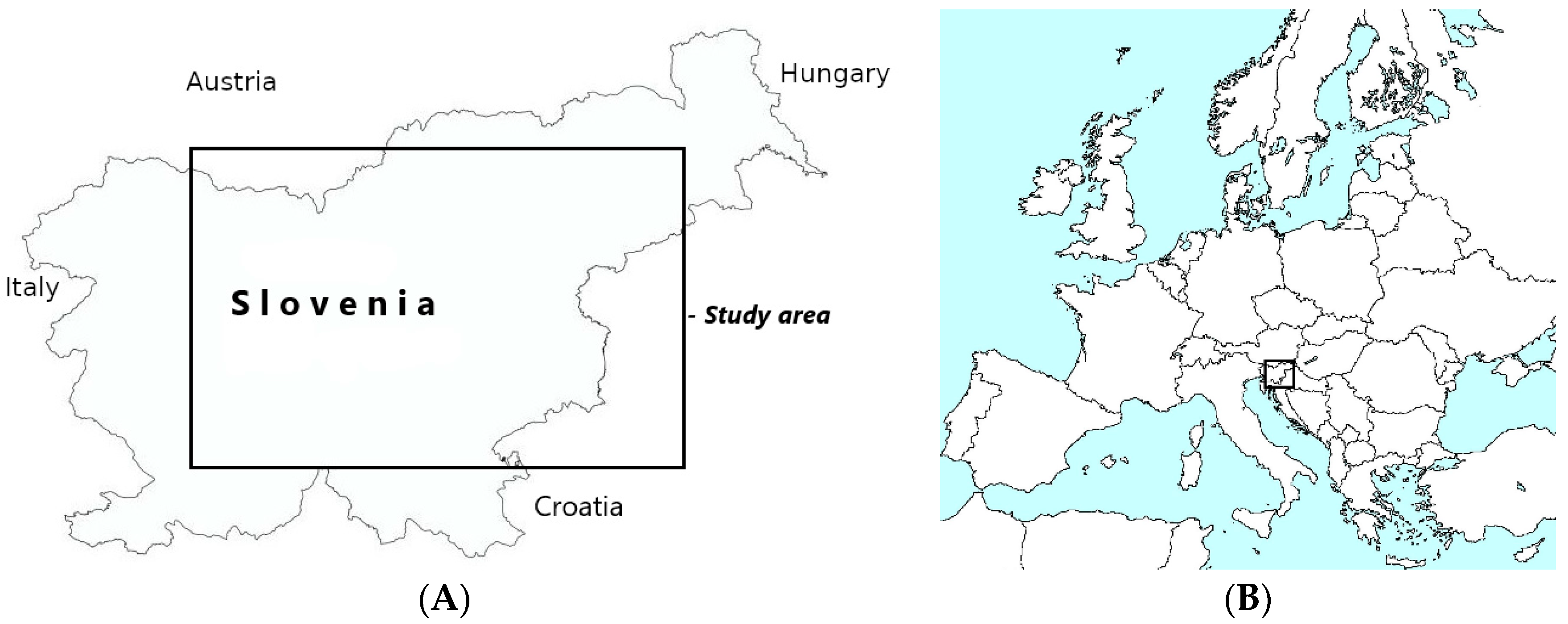
2. Materials and Methods
2.1. Field Survey
- (a)
- Riparian zone properties: width of RZ vegetation (>30 m/5–30 m/1–5 m/<1 m); completeness of RZ vegetation (complete/breaks at >50 m intervals/breaks at 50 m intervals/disturbances frequent); vegetation structure (riparian trees and/or marshy species/pioneer trees and shrubs/herbaceous species, few trees and shrubs/mainly herbaceous species); vegetation height (trees >4 m high/shrubs up to 4 m/herbaceous plants and shrubs 1–2.5 m/herbaceous plants < 1 m); bank structure (stable, firmly held by roots/firm but loosely held by roots/loose/unstable, easily disturbed); bank erosion (little or none/at curves or constrictions/frequent/in entire reach, banks falling in); bank changes (no visible changes/transverse changes/embankment with natural material/embankment with artificial material).
- (b)
- Channel properties: channel depth (width/depth ratio: <7/8–15/15–25/>25); channel changes (none/deepened or widened/dams from natural materials/dams from artificial material); riffles & pools (at every 5–7 stream widths/irregularly spaced/long pools following short riffles/channel regulated); current velocity (torrential/very fast/fast/moderate/slow/not visible); water depth (>1 m/0.3–1 m/<0.3 m).
- (c)
- Land-use: land-use adjacent to RZ (≤30 m) (forest, wetland/mixed pasture, wood, swamp, arable land/agricultural landscape/urban landscape, with gardens and ruderal areas); land-use in wider catchment area (≤100 m), which was estimated using Atlas of Environment [45].
2.2. Data Analyses
3. Results
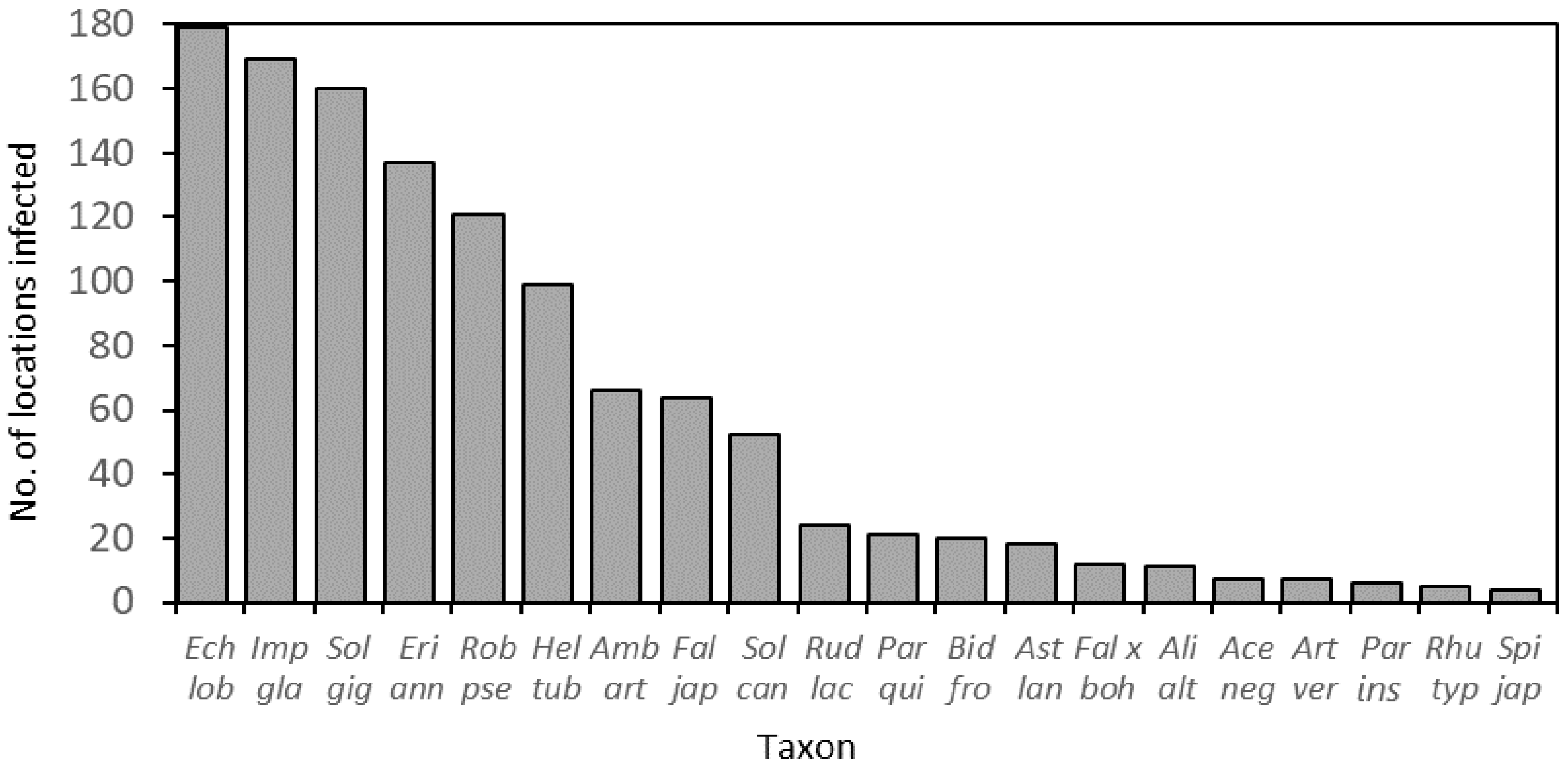
3.1. IAS Frequency, Abundance, Life Forms, and Distributions
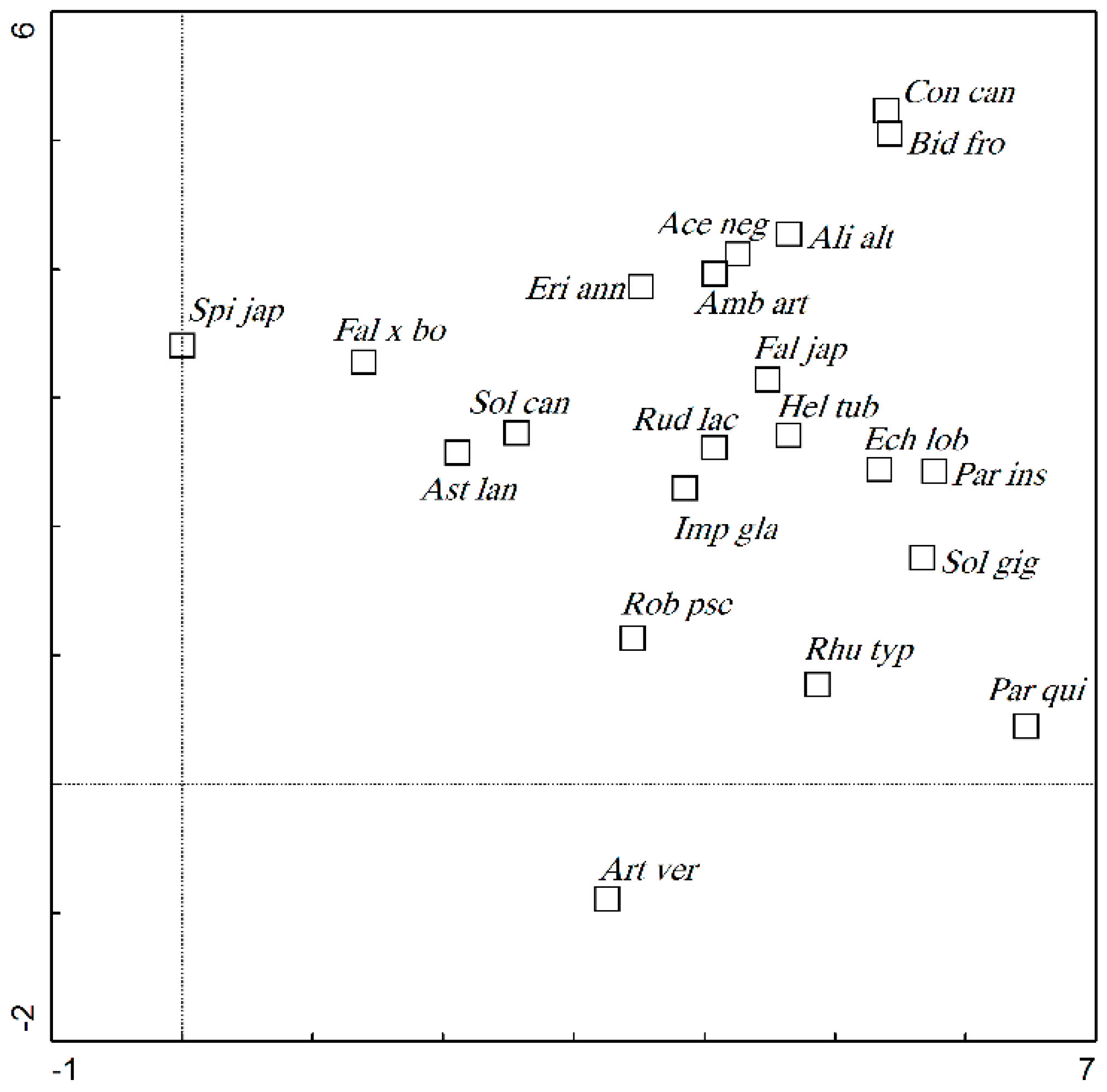
3.2. Relationships Between Environmental Factors and IAS Presence, Abundance, and Form
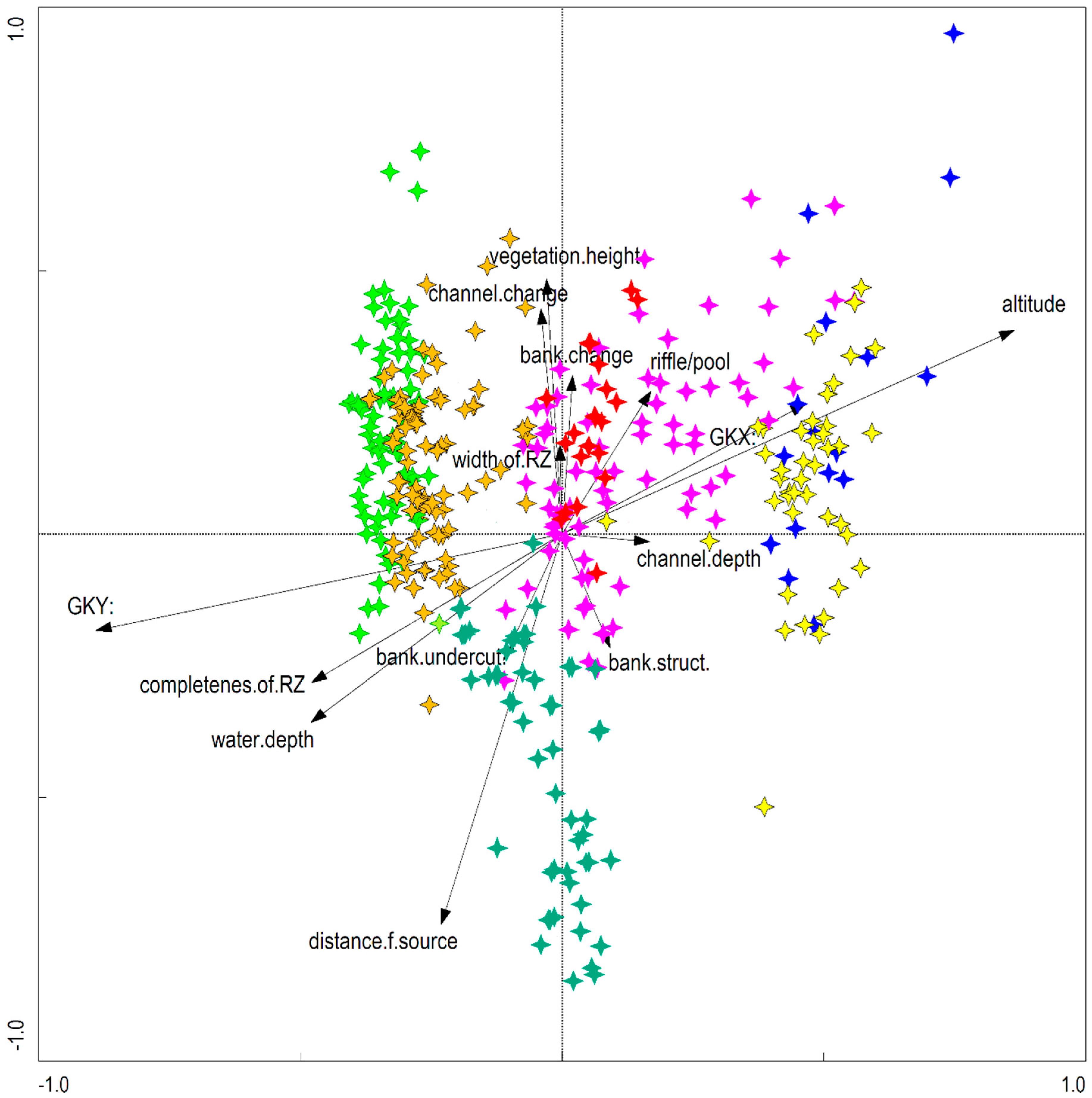
3.3. Impact of Environmental Factors on the Distribution of IAS in the Riparian Zones
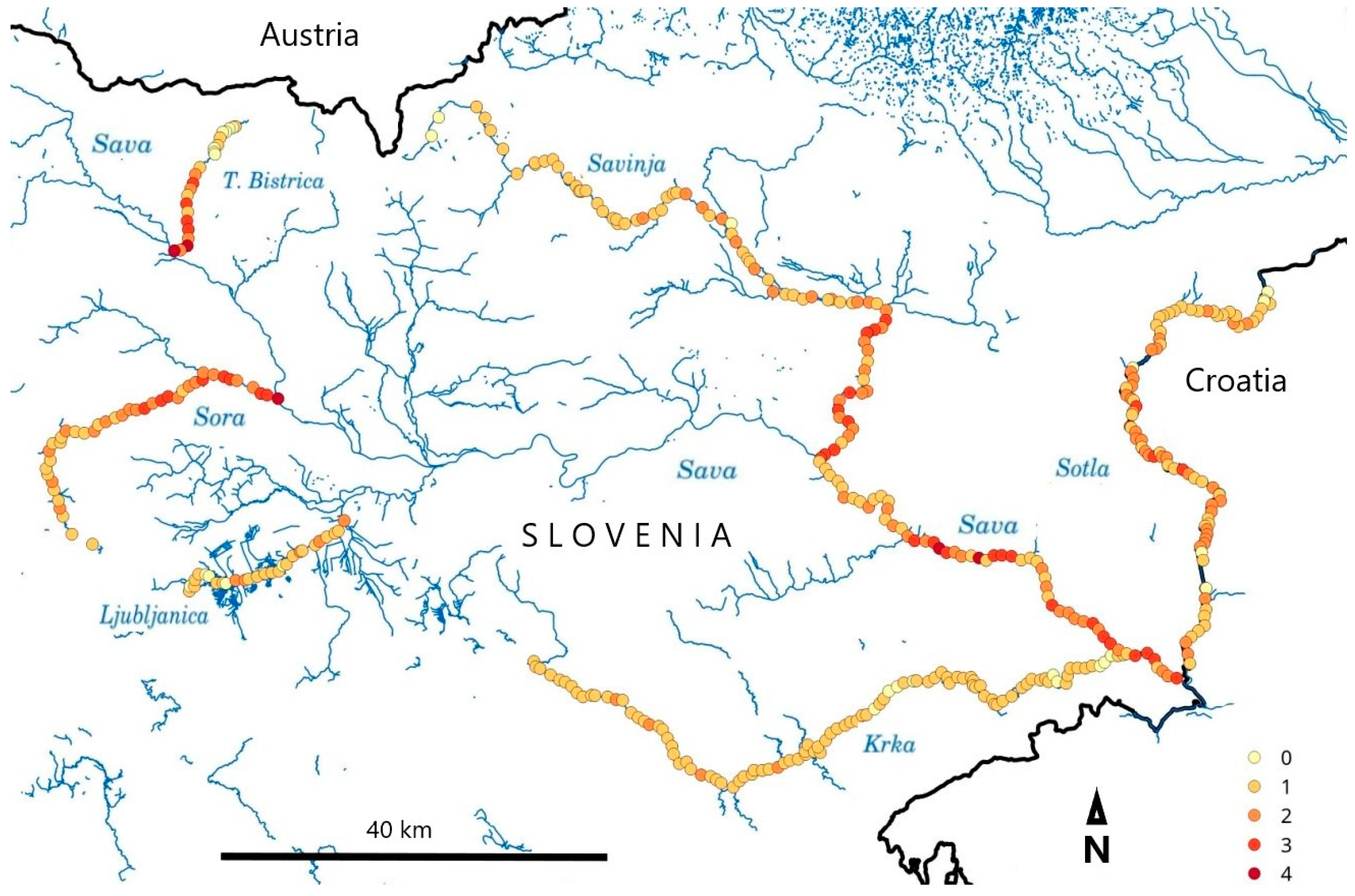
3.4. Comparisons of Reaches with Different IAS Abundance
4. Discussion
4.1. Presence and Abundance of IAS
4.2. Impact of Environmental Factors
4.3. Comparison of Reaches with Different IAS Abundance
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dufour, S.; Rodríguez-González, P.M.; Laslier, M. Tracing the scientific trajectory of riparian vegetation studies: Main topics, approaches and needs in a globally changing world. Sci. Total Environ. 2019, 653, 1168–1185. [Google Scholar] [CrossRef] [PubMed]
- Naiman, R.J.; Décamps, H. The ecology of interfaces: Riparian zones. Ann. Rev. Ecol. Syst. 1997, 28, 621–658. [Google Scholar] [CrossRef] [Green Version]
- Schnitzler, A.; Hale, B.W.; Alsum, E.M. Examining native and exotic species diversity in European riparian forests. Biol. Conserv. 2007, 138, 146–156. [Google Scholar] [CrossRef]
- Douda, J.; Boublík, K.; Slezák, M.; Biurrun, I.; Nociar, J.; Havrdová, A.; Doudová, J.; Aćić, S.; Brisse, H.; Brunet, J.; et al. Vegetation classification and biogeography of European floodplain forests and alder carrs. Appl. Veg. Sci. 2016, 19, 147–163. [Google Scholar] [CrossRef]
- Gaberščik, A. Meje rečnega ekosistema. In Urejenost Voda—Osnova Ali Omejitev, Proceedings of the Mišičev vodarski Dan, Maribor, Slovenia, December 6, 1996; Vodnogospodarski Biro: Maribor, Slovenia, 1996; pp. 72–77. [Google Scholar]
- Naiman, R.J.; Decamps, H.; McClain, M.E. Riparia: Ecology, Conservation, and Management of Streamside Communities; Elsevier: London, UK, 2010; p. 430. ISBN 978-0-08-047068-9. [Google Scholar]
- Kuhar, U.; Gregorc, T.; Renčelj, M.; Šraj-Kržič, N.; Gaberščik, A. Distribution of macrophytes and condition of the physical environment of streams flowing through agricultural landscape in north-eastern Slovenia. Limnologica 2007, 37, 146–154. [Google Scholar] [CrossRef] [Green Version]
- Šraj-Kržič, N.; Germ, M.; Urbanc-Berčič, O.; Kuhar, U.; Janauer, G.A.; Gaberščik, A. The quality of the aquatic environment and macrophytes of karstic watercourses. Plant Ecol. 2007, 192, 107–118. [Google Scholar] [CrossRef]
- Germ, M.; Urbanc-Berčič, O.; Janauer, G.A.; Filzmoser, P.; Exler, N.; Gaberščik, A. Macrophyte distribution pattern in the Krka River—The role of habitat quality. Large Rivers 2008, 18, 145–155. [Google Scholar] [CrossRef]
- Zelnik, I.; Haler, M.; Gaberščik, A. Vulnerability of a riparian zone towards invasion by alien plants depends on its structure. Biologia 2015, 70, 869–878. [Google Scholar] [CrossRef]
- Pollock, M.M.; Naiman, R.J.; Hanley, T.A. Plant species richness in riparian wetlands—A test of biodiversity theory. Ecology 1998, 79, 94–105. [Google Scholar] [CrossRef]
- Ward, J.V.; Tockner, K.; Arscott, D.B.; Claret, C. Riverine landscape diversity. Freshw. Biol. 2002, 47, 517–539. [Google Scholar] [CrossRef] [Green Version]
- Wierzcholska, S.; Dyderski, M.K.; Pielech, R.; Gazda, A.; Smoczyk, M.; Malicki, M.; Horodecki, P.; Kamczyc, J.; Skorupski, M.; Hachułka, M.; et al. Natural forest remnants as refugia for bryophyte diversity in a transformed mountain river valley landscape. Sci. Total Environ. 2018, 640, 954–964. [Google Scholar] [CrossRef] [PubMed]
- Forman, R.T.T. Land Mosaics: The Ecology of Landscapes and Regions, 1st ed.; Cambridge University Press: Cambridge, UK, 1995; ISBN 978-0-521-47980-6. [Google Scholar]
- Ward, J.V. Riverine landscapes: Biodiversity patterns, disturbance regimes, and aquatic conservation. Biol. Conserv. 1998, 83, 269–278. [Google Scholar] [CrossRef]
- Meyer, J.L.; Sale, M.J.; Mulholland, P.J.; Poff, N.L. Impacts of climate change on aquatic ecosystem functioning and health. J. Am. Water Resour. Assoc. 1999, 35, 1373–1386. [Google Scholar] [CrossRef]
- Tickner, D.P.; Angold, P.G.; Gurnell, A.M.; Mountford, J.O. Riparian plant invasions: Hydrogeomorphological control and ecological impacts. Prog. Phys. Geogr. Earth Environ. 2001, 25, 22–52. [Google Scholar] [CrossRef]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef]
- Saunders, D.A.; Hobbs, R.J.; Margules, C.R. Biological Consequences of Ecosystem Fragmentation: A Review. Conserv. Biol. 1991, 5, 18–32. [Google Scholar] [CrossRef]
- Turner, M.G. Landscape Ecology: What is the state of the science? Annu. Rev. Ecol. Evol. Syst. 2005, 36, 319–344. [Google Scholar] [CrossRef]
- Guirado, M.; Pino, J.; Rodà, F. Understorey plant species richness and composition in metropolitan forest archipelagos: Effects of forest size, adjacent land use and distance to the edge. Glob. Ecol. Biogeogr. 2006, 15, 50–62. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Humphries, S.E. An integrated approach to the ecology and management of plant invasions. Conserv. Biol. 1995, 9, 761–770. [Google Scholar] [CrossRef] [Green Version]
- Ohlemüller, R.; Walker, S.; Wilson, J.B. Local vs regional factors as determinants of the invasibility of indigenous forest fragments by alien plant species. Oikos 2006, 112, 493–501. [Google Scholar] [CrossRef]
- Gassó, N.; Sol, D.; Pino, J.; Dana, E.D.; Lloret, F.; Sanz-Elorza, M.; Sobrino, E.; Vilà, M. Exploring species attributes and site characteristics to assess plant invasions in Spain. Divers. Distrib. 2009, 15, 50–58. [Google Scholar] [CrossRef]
- Vilà, M.; Ibáñez, I. Plant invasions in the landscape. Landsc. Ecol. 2011, 26, 461–472. [Google Scholar] [CrossRef]
- González-Moreno, P.; Pino, J.; Gassó, N.; Vilà, M. Landscape context modulates alien plant invasion in Mediterranean forest edges. Biol. Invasions 2013, 15, 547–557. [Google Scholar] [CrossRef] [Green Version]
- Hood, W.G.; Naiman, R.J. Vulnerability of riparian zones to invasion by exotic vascular plants. Plant Ecol. 2000, 148, 105–114. [Google Scholar] [CrossRef]
- Washitani, I. Plant conservation ecology for management and restoration of riparian habitats of lowland Japan. Popul. Ecol. 2001, 43, 189–195. [Google Scholar] [CrossRef]
- Regulation (EU) No 1143/2014 of the European Parliament and of the Council of 22 October 2014 on the Prevention and Management of the Introduction and Spread of Invasive Alien Species; 2014; L 317/35. Available online: https://eur-lex.europa.eu/eli/reg/2014/1143/oj (accessed on 17 April 2015).
- Hejda, M.; Pyšek, P. What is the impact of Impatiens glandulifera on species diversity of invaded riparian vegetation? Biol. Conserv. 2006, 132, 143–152. [Google Scholar] [CrossRef]
- Dassonville, N.; Vanderhoeven, S.; Gruber, W.; Meerts, P. Invasion by Fallopia japonica increases topsoil mineral nutrient concentrations. Ecoscience 2007, 14, 230–240. [Google Scholar] [CrossRef] [Green Version]
- Hulme, P.E.; Bremner, E.T. Assessing the impact of Impatiens glandulifera on riparian habitats: Partitioning diversity components following species removal. Funct. Ecol. 2010, 43, 43–50. [Google Scholar] [CrossRef]
- Robertson, D.J.; Coll, M. Effects of Riparian Invasive Nonindigenous Plants on Freshwater Quantity and Ecological Functioning in Mesic Temperate Landscapes. Nat. Areas J. 2019, 39, 22–32. [Google Scholar] [CrossRef]
- Ewel, K.C.; Cressa, C.; Kneib, R.T.; Lake, P.S.; Levin, L.A.; Palmer, M.A.; Snelgrove, P.; Wall, D.H. Managing Critical Transition Zones. Ecosystems 2001, 4, 452–460. [Google Scholar] [CrossRef]
- Planty-Tabacchi, A.-M.; Tabacchi, E.; Naiman, R.J.; Deferrari, C.; Décamps, H. Invasibility of species-rich communities in riparian zones. Conserv. Biol. 1996, 10, 598–607. [Google Scholar] [CrossRef]
- Stachowicz, J.J.; Tilman, D. Species invasions and the relationships between species diversity, community saturation and ecosystem functioning. In Species Invasions: Insights into Ecology, Evolution, and Biogeography; Sax, D.F., Stachowicz, J.J., Gaines, S.D., Eds.; Sinauer Associates: Sunderland, MA, USA, 2005; pp. 41–46. ISBN 978-0-87893-811-7. [Google Scholar]
- McKinney, M.L.; Lockwood, J.L. Community composition and homogenization: Evenness and abundance of native and exotic plant species. In Species invasions: Insights into Ecology, Evolution, and Biogeography; Sax, D.F., Stachowicz, J.J., Gaines, S.D., Eds.; Sinauer Associates: Sunderland, MA, USA, 2005; pp. 365–380. ISBN 978-0-87893-811-7. [Google Scholar]
- Rowntree, K. An assessment of the potential impact of alien invasive vegetation on the geomorphology of river channels in South Africa. S. Afr. J. Aquat. Sci. 1991, 17, 28–43. [Google Scholar] [CrossRef]
- Merritt, D.M.; Wohl, E.E. Processes governing hydrochory along rivers: Hydraulics, hydrology, and dispersal phenology. Ecol. Appl. 2002, 12, 1071–1087. [Google Scholar] [CrossRef]
- Richardson, D.M.; Holmes, P.M.; Esler, K.J.; Galatowitsch, S.M.; Stromberg, J.C.; Kirkman, S.P.; Pyšek, P.; Hobbs, R.J. Riparian vegetation: Degradation, alien plant invasions, and restoration prospects. Divers. Distrib. 2007, 13, 126–139. [Google Scholar] [CrossRef]
- Zelnik, I. The presence of invasive alien plant species in different habitats: Case study from Slovenia. Acta Biol. Slov. 2012, 55, 25–38. [Google Scholar]
- Wagner, V.; Chytrý, M.; Jiménez-Alfaro, B.; Pergl, J.; Hennekens, S.; Biurrun, I.; Knollová, I.; Berg, C.; Vassilev, K.; Rodwell, J.S.; et al. Alien plant invasions in European woodlands. Divers. Distrib. 2017, 23, 969–981. [Google Scholar] [CrossRef]
- Marinšek, A.; Kutnar, L. Occurrence of invasive alien plant species in the floodplain forests along the Mura River in Slovenia. Period. Biol. 2017, 119, 251–260. [Google Scholar] [CrossRef]
- Strahler, A.N. Dynamic basis of geomorphology. Geol. Soc. Am. Bull. 1952, 63, 923–938. [Google Scholar] [CrossRef]
- Atlas of the Environment. Available online: http://gis.arso.gov.si/atlasokolja (accessed on 5 May 2016).
- Petersen, R.C. The RCE: A Riparian, Channel, and Environmental inventory for small streams in the agricultural landscape. Freshw. Biol. 1992, 27, 295–306. [Google Scholar] [CrossRef]
- Cook, B.J.; Hauer, F.R. Effects of hydrologic connectivity on water chemistry, soils, and vegetation structure and function in an intermontane depressional wetland landscape. Wetlands 2007, 27, 719–738. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Hudon, C.; Gagnon, P.; Amyot, J.-P.; Létourneau, G.; Jean, M.; Plante, C.; Rioux, D.; Deschênes, M. Historical changes in herbaceous wetland distribution induced by hydrological conditions in Lake Saint-Pierre (St. Lawrence River, Quebec, Canada). Hydrobiologia 2005, 539, 205–224. [Google Scholar] [CrossRef]
- QGIS—Free and Open-Source Desktop Geographic Information System Application. Available online: https://www.qgis.org/en/site/ (accessed on 9 February 2016).
- Stohlgren, T.J.; Bull, K.A.; Otsuki, Y.; Villa, C.A.; Lee, M. Riparian zones as havens for exotic plant species in the central grasslands. Plant Ecol. 1998, 138, 113–125. [Google Scholar] [CrossRef]
- Assini, S.; Bracco, F.; Sartori, F. Plant invasion in riparian habitats in Northern Italy. In Plant Invasions in Italy; Grapow, L., Pretto, F., Brundu, G., Carli, E., Blasi, C., Eds.; Palombi & Partner srl: Rome, Italy, 2009; p. 15. [Google Scholar]
- Essl, F.; Rabitsch, W. Neobiota in Österreich; Umweltbundesamt: Wien, Austria, 2002; ISBN 978-3-85457-658-7. [Google Scholar]
- Pyšek, P.; Chytrý, M.; Pergl, J.; Sádlo, J.; Wild, J. Plant invasions in the Czech Republic: Current state, introduction dynamics, invasive species and invaded habitats. Preslia 2012, 84, 575–629. [Google Scholar]
- Malanson, G.P. Riparian Landscapes; Cambridge University Press: Cambridge, UK, 1993; p. 296. ISBN 978-0-521-38431-5. [Google Scholar]
- Ewers, F.W.; Rosell, J.A.; Olson, M.E. Lianas as Structural Parasites. In Functional and Ecological Xylem Anatomy; Hacke, U., Ed.; Springer International Publishing Switzerland: Cham, Switzerland, 2015; pp. 163–188. ISBN 978-3-319-15783-2. [Google Scholar] [CrossRef]
- Flanagan, N.E.; Richardson, C.J.; Ho, M. Connecting differential responses of native and invasive riparian plants to climate change and environmental alteration. Ecol. Appl. 2015, 25, 753–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruno, J.F.; Fridley, J.D.; Bromberg, K.D.; Bertness, M.D. Insights into biotic interactions from studies of species invasions. In Species Invasions: Insights into Ecology, Evolution, and Biogeography; Sax, D.F., Stachowicz, J.J., Gaines, S.D., Eds.; Sinauer Associates: Sunderland, MA, USA, 2005; pp. 13–40. ISBN 978-0-87893-811-7. [Google Scholar]
- Grime, J.P. Plant strategies and vegetation processes. In Plant Strategies and Vegetation Processes; John Wiley & Sons: Chichester, UK, 1979; p. 417. [Google Scholar]
- Jauni, M.; Hyvönen, T. Interactions between alien plant species traits and habitat characteristics in agricultural landscapes in Finland. Biol. Invasions 2012, 14, 47–63. [Google Scholar] [CrossRef]
- Rehounková, K.; Prach, K. Spontaneous vegetation succession in gravel-sand pits: A potential for restoration. Restor. Ecol. 2008, 16, 305–312. [Google Scholar] [CrossRef]
- Cumming, G. Habitat shape, species invasions, and reserve design: Insights from simple models. Conserv. Ecol. 2002, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Lyon, J.; Gross, N.M. Patterns of plant diversity and plant–environmental relationships across three riparian corridors. For. Ecol. Manag. 2005, 204, 267–278. [Google Scholar] [CrossRef]
- Loo, S.E.; Nally, R.M.; O’Dowd, D.J.; Lake, P.S. Secondary invasions: Implications of riparian restoration for in-stream invasion by an aquatic grass. Restor. Ecol. 2009, 17, 378–385. [Google Scholar] [CrossRef]
- Allan, J.D.; Castillo, M.M. Stream Ecology: Structure and Function of Running Waters, 2nd ed.; Springer: Dordrecht, The Netherlands, 2007; ISBN 978-1-4020-5582-9. [Google Scholar]
- Pielech, R.; Anioł-Kwiatkowska, J.; Szczęśniak, E. Landscape-scale factors driving plant species composition in mountain streamside and spring riparian forests. Forest Ecol. Manag. 2015, 347, 217–227. [Google Scholar] [CrossRef]
- Lepori, F.; Hjerdt, N. Disturbance and Aquatic Biodiversity: Reconciling Contrasting Views. BioScience 2006, 56, 809–818. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization, Biodiversity, and Conservation: The impacts of urbanization on native species are poorly studied, but educating a highly urbanized human population about these impacts can greatly improve species conservation in all ecosystems. BioScience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Klinger, R.; Underwood, E.C.; Moore, P.E. The role of environmental gradients in non-native plant invasion into burnt areas of Yosemite National Park, California. Divers. Distrib. 2006, 12, 139–156. [Google Scholar] [CrossRef] [Green Version]
- Körner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| River | Altitude (m a.s.l.) | Distance from River’s source (km) | Nr. of Reaches | Year of Survey | Length of River in Slovenia | Max. Order of | Eco-Region | ||
|---|---|---|---|---|---|---|---|---|---|
| Max. | Min. | Min. | Max. | (km) | River | ||||
| Sava | 191 | 134 | 158 | 213 | 55 | 2013 | 221 | 6 | SP |
| Savinja | 779 | 191 | 0.1 | 98 | 80 | 2011 | 99 | 4 | AL, SP |
| Krka | 268 | 140 | 0.2 | 95 | 100 | 2012 | 95 | 4 | DN, SP |
| Sotla | 332 | 136 | 1 | 86 | 86 | 2009 | 90 | 3 | SP |
| Sora | 570 | 309 | 3.7 | 52 | 42 | 2012 | 52 | 4 | AL |
| Ljubljanica | 289 | 284.5 | 0.1 | 28 | 28 | 2011 | 41 | 4 | DN |
| Tržiška Bistrica | 766 | 371 | 8.1 | 27.5 | 23 | 2011 | 27.5 | 3 | AL |
| Parameter | Spatial | Land Use | Riparian Zone Vegetation | Banks | Channel | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IAS | Altitude | Distance from Source | Adjacent | Catchment Area | Width | Completeness | Structure | Height | Structure | Changes | Changes | Riffles & Pools | Current Velocity | Water Depth | |
| All | Total abundance | −0.042 | 0.332 ** | −0.052 | −0.090 | −0.023 | −0.042 | −0.029 | −0.181 * | 0.077 | −0.066 | −0.138 * | 0.027 | 0.139 * | 0.000 |
| number | −0.061 | 0.385 ** | −0.072 | −0.062 | 0.016 | −0.089 | −0.062 | −0.166 * | 0.030 | −0.101 * | −0.146 * | −0.040 | 0.108 * | 0.138 * | |
| presence | −0.074 | 0.155 * | −0.155 * | −0.158 * | −0.156 * | −0.164 * | −0.131 * | −0.157 * | 0.015 | −0.068 | −0.098 * | −0.008 | 0.017 | 0.078 | |
| Trees | Total abundance | −0.067 | 0.082 | −0.021 | −0.048 | 0.081 | 0.074 | 0.080 | 0.030 | 0.062 | −0.088 | −0.078 | −0.072 | 0.050 | 0.020 |
| number | −0.056 | 0.070 | −0.016 | −0.037 | 0.073 | 0.063 | 0.071 | 0.024 | 0.031 | −0.071 | −0.063 | −0.083 | 0.034 | 0.057 | |
| Annuals | Total abundance | 0.209 * | 0.169 * | −0.081 | 0.027 | −0.071 | −0.291 ** | −0.116 * | −0.172 * | 0.122 * | −0.063 | −0.072 | 0.118 * | 0.162 * | −0.021 |
| number | 0.179 * | 0.177 * | −0.100 * | 0.039 | −0.056 | −0.313 ** | −0.164 * | −0.205 * | 0.081 | −0.099 * | −0.089 | 0.075 | 0.123 * | 0.050 | |
| Perennials | Total abundance | 0.000 | 0.284 ** | −0.014 | −0.103 * | 0.005 | 0.070 | −0.017 | −0.157 * | 0.060 | −0.069 | −0.162 * | −0.085 | 0.132 * | −0.029 |
| number | 0.011 | 0.341 ** | −0.016 | −0.086 | 0.030 | 0.030 | −0.019 | −0.128 * | 0.052 | −0.092 | −0.173 * | −0.128 * | 0.125 * | 0.022 | |
| Vines | Total abundance | −0.437 ** | 0.287 ** | −0.076 | −0.107 * | 0.031 | 0.083 | 0.022 | −0.019 | −0.180 * | 0.010 | 0.020 | 0.027 | −0.156 * | 0.254 ** |
| number | −0.433 ** | 0.293 ** | −0.075 | −0.084 | 0.052 | 0.096 | 0.015 | −0.005 | −0.164 * | 0.000 | 0.010 | 0.021 | −0.163 * | 0.245 ** | |
| Parameter | Specific | 1—IAS Absent | 2—≤20 IAS Individuals | 3—Several IAS Stands | 4—IAS Dominant | ||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | S.D. | Mean | S.D. | Mean | S.D. | Mean | S.D. | ||
| Spatial | GKY | 501,096 | ±48,725 ab | 510,244 | ±36,273 a | 515,831 | ±47,810 b | 492,717 | ±52,087 a |
| GKX | 104,411 | ±24,122 a | 90,866 | ±16,860 b | 100,475 | ±13,755 ab | 106,636 | ±12,673 a | |
| Altitude (m a.s.l.) | 355 | ±236 ab | 242 | ±114 a | 229 | ±99 a | 280 | ±116 b | |
| Distance from source (km) | 34 | ±34 a | 57 | ±50 b | 76 | ±64 c | 63 | ±63 bc | |
| Land-use | Adjacent | 3.5 | ±1.3 a | 2.7 | ±1.3 b | 2.7 | ±1.0 b | 2.8 | ±0.9 b |
| Catchment | 2.9 | ±0.9 a | 2.3 | ±1.0 b | 2.2 | ±1.0 b | 2.2 | ±0.9 b | |
| Riparian zone | Width | 3.0 | ±0.9 a | 2.5 | ±0.7 bc | 2.5 | ±0.7 b | 2.3 | ±0.7 c |
| vegetation | Completeness | 3.5 | ±0.8 a | 2.6 | ±1.3 b | 3.1 | ±1.2 ac | 2.6 | ±1.3 bc |
| Structure | 3.3 | ±0.8 a | 2.9 | ±0.9 b | 3.0 | ±0.8 b | 2.8 | ±0.9 b | |
| Height | 3.7 | ±0.5 a | 3.3 | ±0.8 b | 3.1 | ±0.9 bc | 2.9 | ±1.0 c | |
| Banks | Structure | 3.3 | ±1.0 ab | 3.4 | ±0.7 a | 3.4 | ±0.8 ab | 3.6 | ±0.8 b |
| Changes | 3.6 | ±0.8 a | 3.3 | ±1.0 a | 3.4 | ±0.8 a | 3.3 | ±1.0 a | |
| Erosion | 3.1 | ±0.8 a | 3.5 | ±0.8 b | 3.4 | ±0.8 bc | 3.3 | ±0.9 ac | |
| Channel | Depth | 2.6 | ±1.2 ab | 2.7 | ±1.0 a | 3.0 | ±1.2 b | 3.5 | ±0.9 c |
| Changes | 3.8 | ±0.6 a | 3.5 | ±0.8 ab | 3.5 | ±0.7 b | 3.5 | ±0.8 b | |
| Riffles/pools present | 2.6 | ±1.2 ab | 2.6 | ±1.1 a | 2.5 | ±1.3 ab | 2.9 | ±1.3 b | |
| Current velocity | 3.2 | ±1.3 ab | 3.2 | ±0.8 a | 3.3 | ±0.8 ab | 3.5 | ±0.9 b | |
| Water depth | 2.1 | ±1.0 a | 2.6 | ±0.7 b | 2.2 | ±0.9 a | 2.1 | ±0.9 a | |
| IAS | Total number | 0 | 0 a | 2.7 | ±1.7 b | 3.7 | ±2.1 c | 3.6 | ±1.7 c |
| Total abundance | 0 | 0 a | 3.7 | ±2.5 b | 7.7 | ±3.5 c | 9.6 | ±3.7 d | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zelnik, I.; Mavrič Klenovšek, V.; Gaberščik, A. Complex Undisturbed Riparian Zones Are Resistant to Colonisation by Invasive Alien Plant Species. Water 2020, 12, 345. https://doi.org/10.3390/w12020345
Zelnik I, Mavrič Klenovšek V, Gaberščik A. Complex Undisturbed Riparian Zones Are Resistant to Colonisation by Invasive Alien Plant Species. Water. 2020; 12(2):345. https://doi.org/10.3390/w12020345
Chicago/Turabian StyleZelnik, Igor, Valentina Mavrič Klenovšek, and Alenka Gaberščik. 2020. "Complex Undisturbed Riparian Zones Are Resistant to Colonisation by Invasive Alien Plant Species" Water 12, no. 2: 345. https://doi.org/10.3390/w12020345
APA StyleZelnik, I., Mavrič Klenovšek, V., & Gaberščik, A. (2020). Complex Undisturbed Riparian Zones Are Resistant to Colonisation by Invasive Alien Plant Species. Water, 12(2), 345. https://doi.org/10.3390/w12020345