Diatom Species Richness in Swiss Springs Increases with Habitat Complexity and Elevation

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
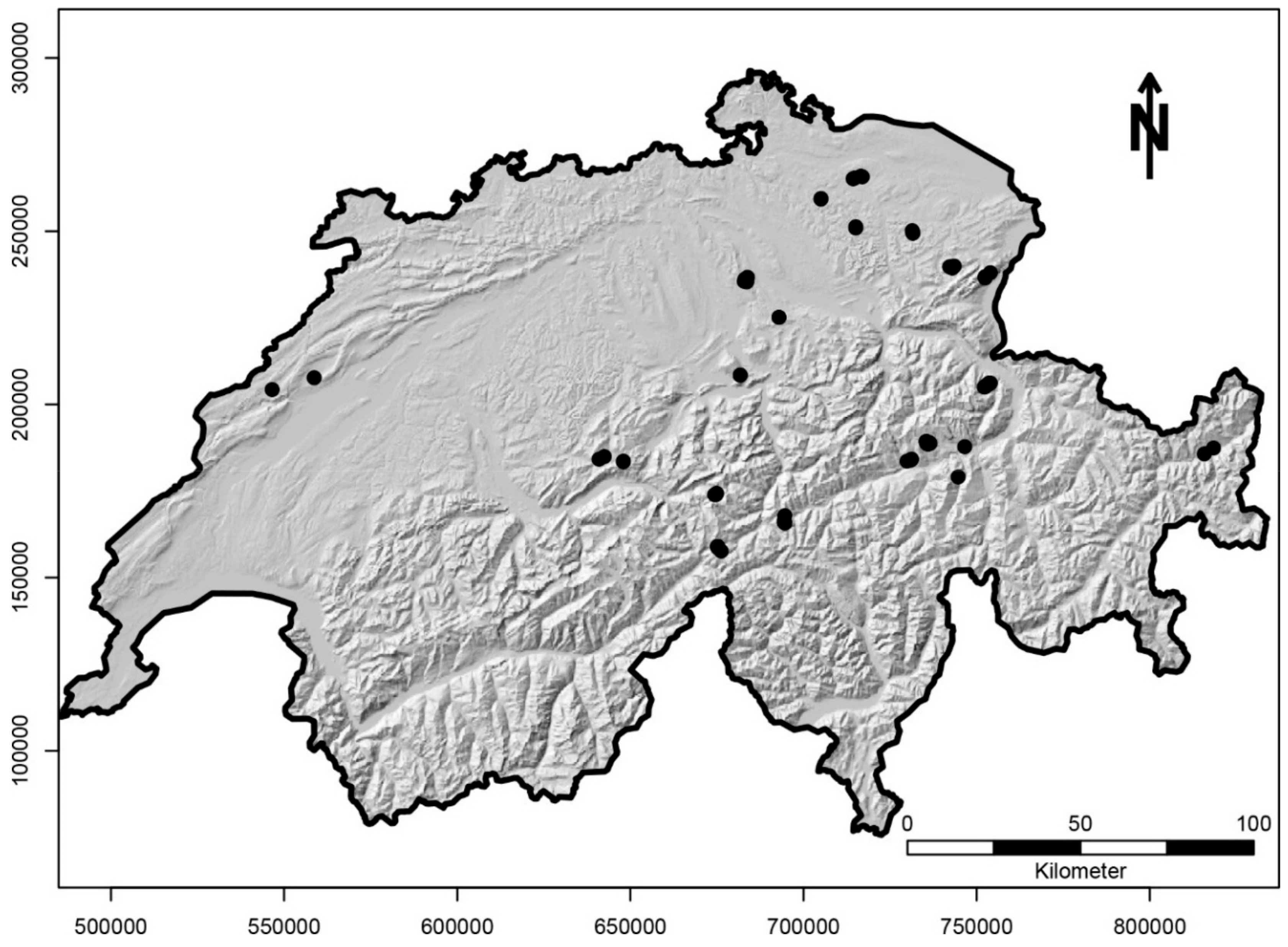
2.1. Study Area and Sites
2.2. Field Sampling
2.3. Laboratory Analyses
2.4. Species Richness and Compositional Turnover
2.5. Explanatory Factors
2.6. Statistical Analyses
3. Results
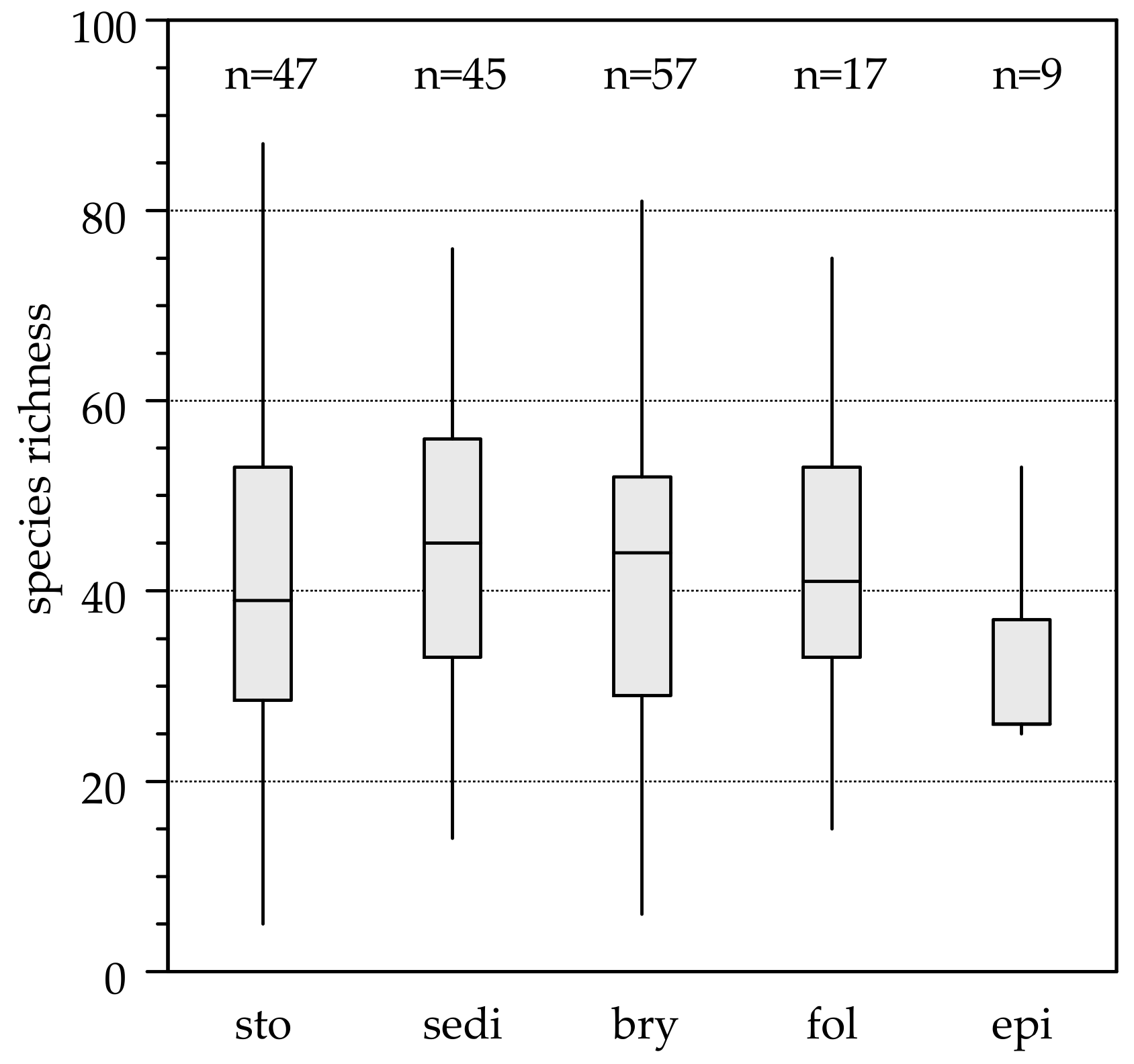
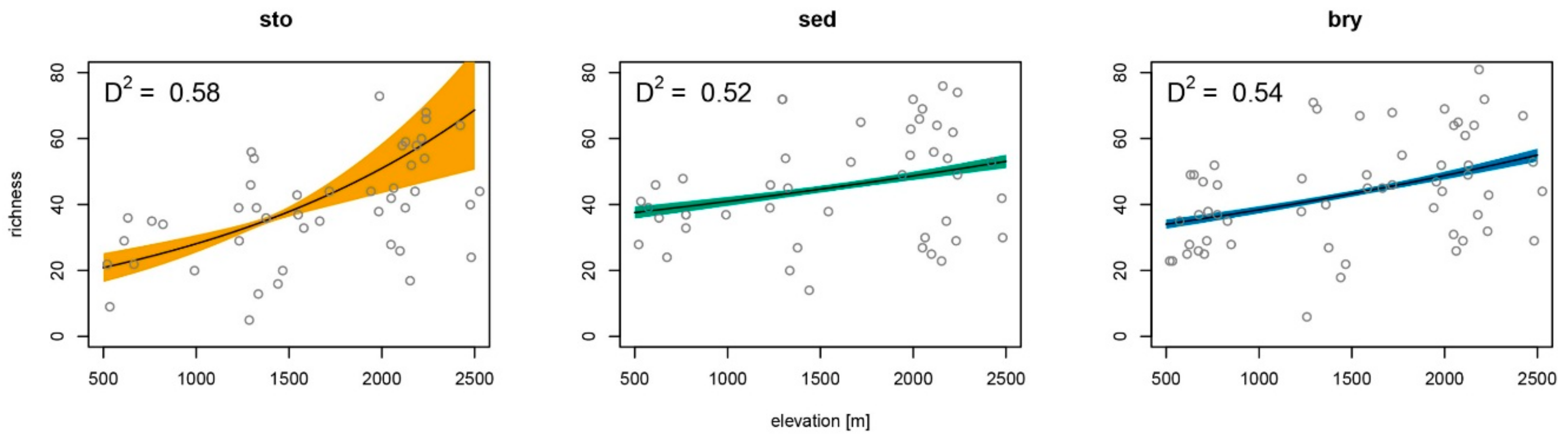
3.1. Habitat Diversity and Species Richness
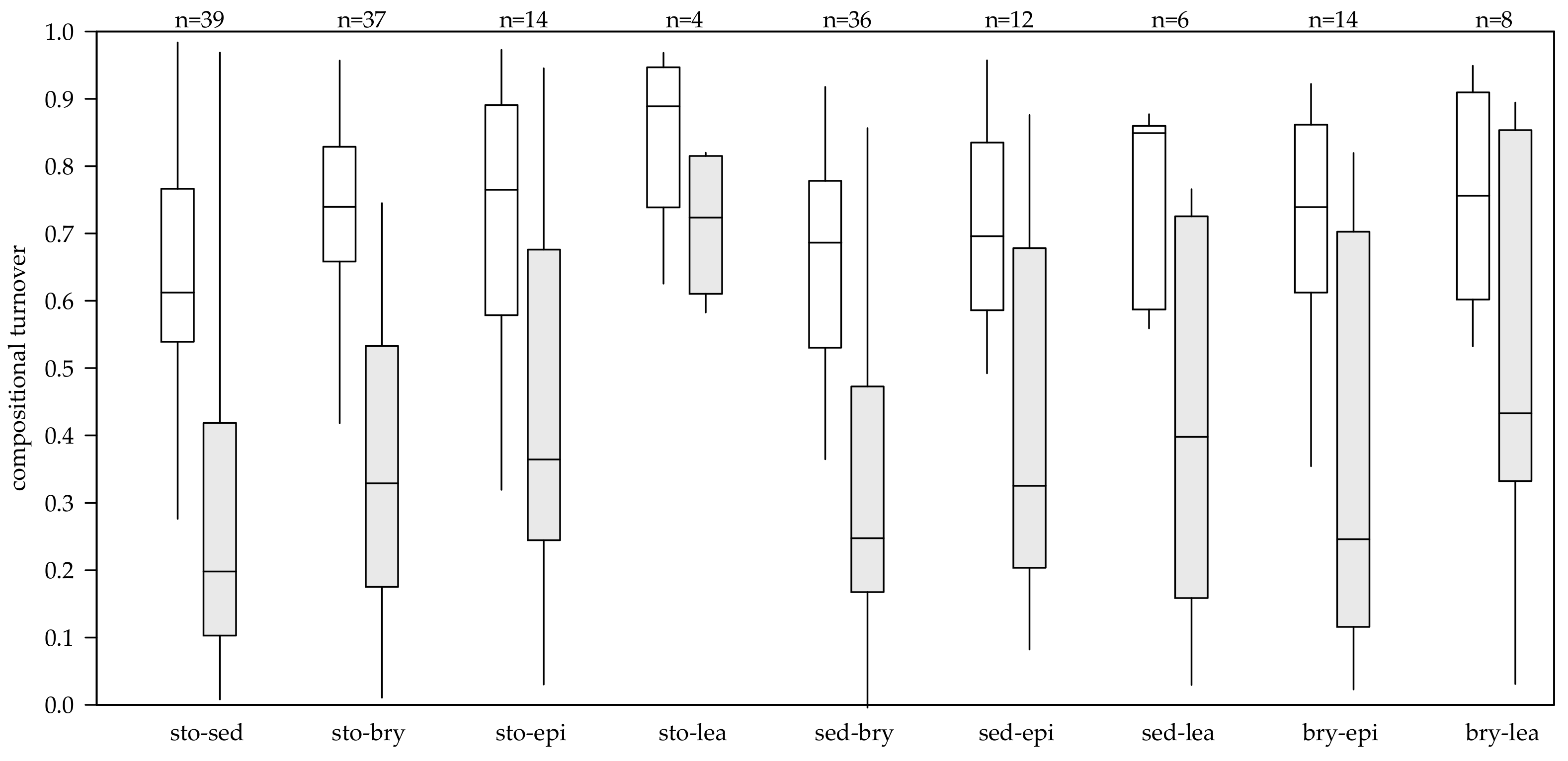
3.2. Compositional Turnover
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kluge, J.; Kessler, M.; Dunn, R.R. What drives elevational patterns of diversity? A test of geometric constraints, climate and species pool effects for pteridophytes on an elevational gradient in Costa Rica. Glob. Ecol. Biogeogr. 2006, 15, 358–371. [Google Scholar] [CrossRef]
- Lomolino, M.V. Elevation gradients of species-density: Historical and prospective views. Glob. Ecol. Biogeogr. 2001, 10, 3–13. [Google Scholar] [CrossRef]
- Rahbek, C. The elevational gradient of species richness: A uniform pattern? Ecography 1995, 18, 200–205. [Google Scholar] [CrossRef]
- Rahbek, C. The role of spatial scale and the perception of large-scale species-richness patterns. Ecol. Lett. 2005, 8, 224–239. [Google Scholar] [CrossRef]
- Grytnes, J.-A.; McCain, C.M. Elevational Trends in Biodiversity. In Encyclopedia of Biodiversity; Elsevier: Amsterdam, The Netherlands, 2007; pp. 1–8. [Google Scholar]
- Wang, J.; Soininen, J.; Zhang, Y.; Wang, B.; Yang, X.; Shen, J. Contrasting patterns in elevational diversity between microorganisms and macroorganisms. J. Biogeogr. 2011, 38, 595–603. [Google Scholar] [CrossRef]
- Wang, J.; Meier, S.; Soininen, J.; Casamayor, E.O.; Pan, F.; Tang, X.; Yang, X.; Zhang, Y.; Wu, Q.; Zhou, J.; et al. Regional and global elevational patterns of microbial species richness and evenness. Ecography 2017, 40, 393–402. [Google Scholar] [CrossRef]
- Round, F.E.; Crawford, R.M.; Mann, D.G. The diatoms. In Biology & Morphology of the Genera; Cambridge University Press: Cambridge, UK, 1990; p. 747. [Google Scholar]
- Smol, J.P.; Stoermer, E.F. The Diatoms: Applications for the Environmental and Earth Sciences, 2nd ed.; Cambridge University Press: Cambridge, UK, 2010; p. 686. [Google Scholar]
- Mann, D.G.; Droop, S.J.M. Biodiversity, biogeography and conservation of diatoms. Hydrobiologia 1996, 336, 19–32. [Google Scholar] [CrossRef]
- Baas Becking, L.G.M. Geobiologie of Inleiding tot de Milieukunde; W. P. Van Stockum & Zoon: The Hague, The Netherlands, 1934. [Google Scholar]
- Finlay, B.J.; Clarke, K.J. Ubiquitous dispersal of microbial species. Nature 1999, 400, 828. [Google Scholar] [CrossRef]
- Bouchard, G.; Gajewski, K.; Hamilton, P.B. Freshwater diatom biogeography in the Canadian Arctic Archipelago. J. Biogeogr. 2004, 31, 1955–1973. [Google Scholar] [CrossRef]
- Hustedt, F. Süßwasser-Diatomeen des indomalayischen Archipels und der Hawaii-Inseln. Nach dem Material der Wallacea-Expedition. Int. Rev. Hydrobiol. 1942, 42, 1–252. [Google Scholar] [CrossRef]
- Lange-Bertalot, H.; Metzeltin, D. Indicators of oligotropy. In 800 Taxa Representative of Three Ecologically Distinct Lake Types Carbonate Buffered, Oligodystrophic, Weakly Buffered Soft Water; Koeltz: Königstein, Germany, 1996; p. 390. [Google Scholar]
- Lowe, R.L.; Kociolek, J.P. New and rare diatoms from “Great-Smoky-Mountains-National-Park”. Nova Hedwig. 1984, 39, 465–476. [Google Scholar]
- Vyverman, W.; Verleyen, E.; Sabbe, K.; Vanhoutte, K.; Sterken, M.; Hodgson, D.A.; Mann, D.G.; Juggins, S.; Van De Vijver, B.; Jones, V.; et al. Historical processes constrain patterns in global diatom diversity. Ecology 2007, 88, 1924–1931. [Google Scholar] [CrossRef] [PubMed]
- Vanormelingen, P.; Verleyen, E.; Vyverman, W. The diversity and distribution of diatoms: From cosmopolitanism to narrow endemism. Biodivers. Conserv. 2008, 17, 393–405. [Google Scholar] [CrossRef]
- Verleyen, E.; Vyverman, W.; Sterken, M.; Hodgson, D.A.; De Wever, A.; Juggins, S.; Van De Vijver, B.; Jones, V.J.; Vanormelingen, P.; Roberts, D.; et al. The importance of dispersal related and local factors in shaping the taxonomic structure of diatom metacommunities. Oikos 2009, 118, 1239–1249. [Google Scholar] [CrossRef]
- Teittinen, A.; Soininen, J. Testing the theory of island biogeography for microorganisms—Patterns for spring diatoms. Aquat. Microb. Ecol. 2015, 75, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Teittinen, A.; Wang, J.; Strömgård, S.; Soininen, J. Local and geographical factors jointly drive elevational patterns in three microbial groups across subarctic ponds. Glob. Ecol. Biogeogr. 2017, 26, 973–982. [Google Scholar] [CrossRef] [Green Version]
- Cantonati, M.; Spitale, D. The role of environmental variables in structuring epiphytic and epilithic diatom assemblages in springs and streams of the Dolomiti Bellunesi National Park (south-eastern Alps). Fundam. Appl. Limnol. 2009, 174, 117–133. [Google Scholar] [CrossRef]
- Cantonati, M.; Lange-Bertalot, H.; Scalfi, A.; Angeli, N. Cymbella tridentina sp. nov. (Bacillariophyta), a crenophilous diatom from carbonate springs of the Alps. J. N. Am. Benthol. Soc. 2010, 29, 775–788. [Google Scholar] [CrossRef]
- Cantonati, M.; Lange-Bertalot, H.; Decet, F.; Gabrieli, J. Diatoms in very-shallow pools of the site of community importance Danta di Cadore Mires (south-eastern Alps), and the potential contribution of these habitats to diatom biodiversity conservation. Nova Hedwig. 2011, 93, 475–507. [Google Scholar] [CrossRef]
- Cantonati, M.; Angeli, N.; Bertuzzi, E.; Spitale, D.; Lange-Bertalot, H. Diatoms in springs of the Alps: Spring types, environmental determinants, and substratum. Freshw. Sci. 2012, 31, 499–524. [Google Scholar] [CrossRef]
- Fránková, M.; Bojková, J.; Poulíčková, A.; Hájek, M. The structure and species richness of the diatom assemblages of the Western Carpathian spring fens along the gradient of mineral richness. Fottea 2009, 9, 355–368. [Google Scholar] [CrossRef] [Green Version]
- Żelazna-Wieczorek, J. Diatom Flora in Springs of Lódz Hills (Central Poland). In Biodiversity, Taxonomy, and Temporal Changes of Epipsammic Diatom Assemblages in Springs Affected by Human Impact; A.R.G. Gantner Verlag, K.G.: Ruggell, Liechtenstein, 2011; p. 419. [Google Scholar]
- Kembel, S.W. Disentangling niche and neutral influences on community assembly: Assessing the performance of community phylogenetic structure tests. Ecol. Lett. 2009, 12, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Lortie, C.J.; Brooker, R.W.; Choler, P.; Kikvidze, Z.; Michalet, R.; Pugnaire, F.I.; Callaway, R.M. Rethinking plant community theory. Oikos 2004, 107, 433–438. [Google Scholar] [CrossRef]
- Tuomisto, H.; Ruokolainen, K.; Aguilar, M.; Sarmiento, A. Floristic patterns along a 43-km long transect in an Amazonian rain forest. J. Ecol. 2003, 91, 743–756. [Google Scholar] [CrossRef]
- Tuomisto, H.; Ruokolainen, K.; Yli-Halla, M. Dispersal, Environment, and Floristic Variation of Western Amazonian Forests. Science 2003, 299, 241–244. [Google Scholar] [CrossRef]
- Vergnon, R.; Dulvy, N.K.; Freckleton, R.P. Niches versus neutrality: Uncovering the drivers of diversity in a species-rich community. Ecol. Lett. 2009, 12, 1079–1090. [Google Scholar] [CrossRef]
- Karger, D.N.; Cord, A.F.; Kessler, M.; Kreft, H.; Kühn, I.; Pompe, S.; Sandel, B.; Cabral, J.S.; Smith, A.B.; Svenning, J.; et al. Delineating probabilistic species pools in ecology and biogeography. Glob. Ecol. Biogeogr. 2016, 25, 489–501. [Google Scholar] [CrossRef] [Green Version]
- Karger, D.N.; Kluge, J.; Krömer, T.; Hemp, A.; Lehnert, M.; Kessler, M. The effect of area on local and regional elevational patterns of species richness. J. Biogeogr. 2011, 38, 1177–1185. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Comita, L.S.; Chase, J.M.; Sanders, N.J.; Swenson, N.G.; Crist, T.O.; Stegen, J.C.; Vellend, M.; Boyle, B.; Anderson, M.J.; et al. Disentangling the drivers of beta diversity along latitudinal and elevational gradients. Science 2011, 333, 1755–1758. [Google Scholar] [CrossRef] [Green Version]
- Lessard, J.-P.; Belmaker, J.; Myers, J.A.; Chase, J.M.; Rahbek, C. Inferring local ecological processes amid species pool influences. Trends Ecol. Evol. 2012, 27, 600–607. [Google Scholar] [CrossRef]
- Ricklefs, R.E. Community Diversity: Relative Roles of Local and Regional Processes. Science 1987, 235, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.J.; Willis, K.J.; Field, R. Scale and species richness: Towards a general, hierarchical theory of species diversity. J. Biogeogr. 2001, 28, 453–470. [Google Scholar] [CrossRef] [Green Version]
- Hubbell, S.P. The Unified Neutral Theory of Biodiversity and Biogeography; Princeton University Press: Princeton, NJ, USA, 2001; p. 375. [Google Scholar]
- Plotkin, J.B.; Muller-Landau, H.C. Sampling the species composition of a landscape. Ecology 2002, 83, 3344–3356. [Google Scholar] [CrossRef]
- Scheiner, S.M. Affinity analysis: Effects of sampling. Vegetatio 1990, 86, 175–181. [Google Scholar] [CrossRef]
- Tuomisto, H. A diversity of beta diversities: Straightening up a concept gone awry. Part 2. Quantifying beta diversity and related phenomena. Ecography 2010, 33, 23–45. [Google Scholar] [CrossRef]
- Tuomisto, H. A diversity of beta diversities: Straightening up a concept gone awry. Part 1. Defining beta diversity as a function of alpha and gamma diversity. Ecography 2010, 33, 2–22. [Google Scholar] [CrossRef]
- Wolda, H. Similarity indexes, sample-size and diversity. Oecologia 1981, 50, 296–302. [Google Scholar] [CrossRef]
- Cantonati, M.; Lange-Bertalot, H. Achnanthidium dolomiticum sp. Nov. (bacillariophyta) from oligotrophic mountain springs and lakes fed by dolomite aquifers 1. J. Phycol. 2006, 42, 1184–1188. [Google Scholar] [CrossRef]
- Cox, E.J. Has the role of the substratum been underestimated for algal distribution patterns in freshwater ecosystems? Biofouling 1988, 1, 49–63. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publisher: Hoboken, NJ, USA, 2013; p. 264. [Google Scholar]
- Chao, A.; Chazdon, R.L.; Colwell, R.K.; Shen, T.-J. A new statistical approach for assessing similarity of species composition with incidence and abundance data. Ecol. Lett. 2004, 8, 148–159. [Google Scholar] [CrossRef]
- Cantonati, M.; Ortler, K. Using spring biota of pristine mountain areas for long-term monitoring. Hydrol. Water Resour. Ecol. Headwaters 1998, 248, 379–385. [Google Scholar]
- Cantonati, M.; Komárek, J.; Montejano, G. Cyanobacteria in ambient springs. Biodivers. Conserv. 2015, 24, 865–888. [Google Scholar] [CrossRef]
- Gerecke, R.; Franz, H.; Cantonati, M. Invertebrate diversity in springs of the National Park Berchtesgaden (Germany): Relevance for long-term monitoring. SIL Proc. 1922–2010 2009, 30, 1229–1233. [Google Scholar] [CrossRef]
- Zechmeister, H.; Mucina, L. Vegetation of European springs: High-rank syntaxa of the Montio-Cardaminetea. J. Veg. Sci. 1994, 5, 385–402. [Google Scholar] [CrossRef]
- Werum, M.; Lange-Bertalot, H. Diatoms in Springs from Central Europe and Elsewhere Under the Influence of Hydrogeology and Anthropogenic Impacts; Gantner: Ruggell, Liechtenstein, 2004; p. 480. [Google Scholar]
- Van der Kamp, G. The hydrogeology of springs in relation to the biodiversity of spring fauna—A review. J. Kans. Entomol. Soc. 1995, 68, 4–17. [Google Scholar]
- Hürlimann, J.; Niederhauser, P. Methoden zur Untersuchung und Beurteilung der Fliessgewässer. In Kieselalgen Stufe F (Flächendeckend); Bundesamt für Umwelt: Bern, Germany, 2007; p. 130. [Google Scholar]
- Hofmann, G.; Werum, M.; Lange-Bertalot, H. Diatomeen im Süßwasser-Benthos Von Mitteleuropa. Bestimmungsflora Kieselalgen Für Die Ökologische Praxis. Über 700 Der Häufigsten Arten Und Ihre Ökologie; A.R.G. Gantner Verlag K.G.: Ruggell, Germany, 2011; p. 908. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. 1. Teil: Naviculaceae; Gustav Fischer Verlag: Stuttgart, NY, USA, 1986; Volume 2/1, p. 876. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. 3. Teil: Centrales, Fragilariaceae, Eunotiaceae; Gustav Fischer Verlag: Stuttgart, NY, USA, 1991; Volume 2, p. 576. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. 4. Teil: Achnanthaceae, Kritische Ergänzungen Zu Navicula (Lineolatae) Und Gomphonema. Gesamtliteraturverzeichnis Teil 1–4; Gustav Fischer Verlag: Stuttgart, NY, USA, 1991; Volume 2/4, p. 437. [Google Scholar]
- Krammer, K. Die Cymbelloiden Diatomeen. Teil 2. Encyonema part., Encyonopsis und Cymbellopsis. Eine Monographie Der Weltweit Bekannten Taxa. J; Cramer: Berlin, Germany; Stuttgart, NY, USA, 1997; Volume 37, p. 469. [Google Scholar]
- Krammer, K. Die Cymbelloiden Diatomeen. Teil 1. Allgemeines Und Encyonema Part. Eine Monographie Der Weltweit Bekannten Taxa. J; Cramer: Berlin, Germany; Stuttgart, NY, USA, 1997; Volume 36, p. 382. [Google Scholar]
- Krammer, K. Cymbopleura, Delicata, Navicymbula, Gomphocymbellopsis, Afrocymbella; A.R.G. Gantner Verlag, K.G.: Ruggell, Liechtenstein, 2000; Volume 4, p. 530. [Google Scholar]
- Lange-Bertalot, H. Navicula Sensu Stricto, 10 Genera Separated from Navicula Sensu Lato, Frustulia; A.R.G. Gantner Verlag, K.G.: Ruggell, Liechtenstein, 2001; Volume 2, p. 526. [Google Scholar]
- Krammer, K. Cymbella; A.R.G. Gantner Verlag, K.G.: Ruggell, Liechtenstein, 2002; Volume 3, p. 584. [Google Scholar]
- Krammer, K. Pinnularia; A.R.G. Gantner Verlag, K.G.: Ruggell, Liechtenstein, 2002; Volume 1, p. 703. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae. Unveränderter Nachdruck; Gustav Fischer Verlag: Stuttgart, Liechtenstein, 2007; Volume 2/2, p. 611. [Google Scholar]
- Levkov, Z. Amphora sensu Lato; A.R.G. Gantner Verlag, K.G.: Ruggell, Liechtenstein, 2009; Volume 5, p. 916. [Google Scholar]
- Lange-Bertalot, H.; Bak, M.; Witkowski, A. Eunotia and Some Related Genera; A.R.G. Gantner Verlag, K.G.: Ruggell, Liechtenstein, 2011; Volume 6, p. 747. [Google Scholar]
- Reichardt, E. Zur Revision Der Gattung Gomphonema. Die Arten Um G. Affine/Insigne, G. Angustatum/Micropus, G. Acuminatum Sowie Gomphonemoide Diatomeen Aus Dem Oberoligozän in Böhmen; Gantner: Ruggell, Liechtenstein, 1999; p. 203. [Google Scholar]
- Chao, A.; Chazdon, R.L.; Colwell, R.K.; Shen, T. A new statistical approach for assessing similarity of species composition with incidence and abundance data. Ecol. Lett. 2005, 8, 148–159. [Google Scholar] [CrossRef]
- Cantonati, M.; Rott, E.; Spitale, D.; Angeli, N.; Komárek, J. Are benthic algae related to spring types? Freshw. Sci. 2012, 31, 481–498. [Google Scholar] [CrossRef]
- Oksanen, J.; Guillaume Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R package version 2.0-7; Creative Commons: San Francisco, CA, USA, 2013. [Google Scholar]
- Cribari-Neto, F.; Zeileis, A. Beta Regression in R. J. Stat. Softw. 2010, 34, 1–24. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing, 3.1.2; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Leingärtner, A.; Krauss, J.; Steffan-Dewenter, I. Species richness and trait composition of butterfly assemblages change along an altitudinal gradient. Oecologia 2014, 175, 613–623. [Google Scholar] [CrossRef]
- Wohlgemuth, T. Modelling floristic species richness on a regional scale: A case study in Switzerland. Biodivers. Conserv. 1998, 7, 159–177. [Google Scholar] [CrossRef]
- Taxböck, L.; Linder, H.P.; Cantonati, M. To what extant are swiss springs refugial habitats for rare and endangered diatom taxa? Water 2017, 9, 967. [Google Scholar] [CrossRef] [Green Version]
- Cantonati, M.; Poikane, S.; Pringle, C.M.; Stevens, L.E.; Heino, E.T.J.; Richardson, J.S.; Bolpagni, R.; Borrini, A.; Cid, N. Special Issue: Multiplicity, Characteristics, Main Impacts, and Stewardship of Natural and Artificial Freshwater Environments: Consequences for Biodiversity Conservation. Water 2020, 12, 260. [Google Scholar] [CrossRef] [Green Version]
- Gremmen, N.J.; Van De Vijver, B.; Frenot, Y.; Lebouvier, M. Distribution of moss-inhabiting diatoms along an altitudinal gradient at sub-Antarctic Îles Kerguelen. Antarct. Sci. 2007, 19, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Ormerod, S.; Rundle, S.; Wilkinson, S.; Daly, G.; Dale, K.; Juttner, I. Altitudinal trends in the diatoms, bryophytes, macroinvertebrates and fish of a Nepalese river system. Freshw. Boil. 1994, 32, 309–322. [Google Scholar] [CrossRef]
- Smucker, N.J.; Vis, M.L. Contributions of habitat sampling and alkalinity to diatom diversity and distributional patterns in streams: Implications for conservation. Biodivers. Conserv. 2011, 20, 643–661. [Google Scholar] [CrossRef]
- Soininen, J.; Eloranta, P. Seasonal persistence and stability of diatom communities in rivers: Are there habitat specific differences? Eur. J. Phycol. 2004, 39, 153–160. [Google Scholar] [CrossRef]
- Potapova, M.; Charles, D.F. Choice of substrate in algae-based water-quality assessment. J. North Am. Benthol. Soc. 2005, 24, 415–427. [Google Scholar] [CrossRef]
- Jüttner, I.; Rothfritz, H.; Ormerod, S. Diatoms as indicators of river quality in the Nepalese Middle Hills with consideration of the effects of habitat-specific sampling. Freshw. Boil. 1996, 36, 475–486. [Google Scholar] [CrossRef]
- Angeli, N.; Cantonati, M.; Spitale, D.; Lange-Bertalot, H. A comparison between diatom assemblages in two groups of carbonate, low-altitude springs with different levels of anthropogenic disturbances. Fottea 2010, 10, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Wojtal, A.Z.; Sobczyk, Ł. The influence of substrates and physicochemical factors on the composition of diatom assemblages in karst springs and their applicability in water-quality assessment. Hydrobiol. 2012, 695, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Prygiel, J.; Carpentier, P.; Almeida, S.; Coste, M.; Druart, J.-C.; Ector, L.; Guillard, D.; Honoré, M.-A.; Iserentant, R.; Ledeganck, P.; et al. Determination of the biological diatom index (IBD NF T 90–354): Results of an intercomparison exercise. Environ. Boil. Fishes 2002, 14, 27–39. [Google Scholar]
- Cantonati, M.; Gerecke, R.; Juttner, I.; Cox, E.J. Springs: Neglected key habitats for biodiversity conservation introduction to the special issue. J. Limnol. 2011, 70, 1. [Google Scholar] [CrossRef]
- Kociolek, J.P.; Stoermer, E.F. Oligotrophy: The forgotten end of an ecological spectrum. Acta Bot. Croat. 2009, 68, 465–472. [Google Scholar]
- Potapova, M.; Charles, D.F. Potential use of rare diatoms as environmental indicators in U.S.A. Rivers. In Seventeenth International Diatom Symposium; Poulin, M., Ed.; Biopress Limited: Ottawa, ON, Canada, 2002; pp. 281–295. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Richness | Elevation | Cond | Shading | pH | Flow | Temp | P | N | Cl | HD | n |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sall | 2.221 * | −1.543 | 2.511 * | 0.982 | 1.117 | 4.953 *** | 47 | ||||
| Ssto | 5.667 *** | 3.51 ** | 3.845 *** | 3.027 ** | 1.439 | 2.645 * | −1.87 | 32 | |||
| Ssed | 2.188 * | 1.777 | −1.705 | 0.968 | 31 | ||||||
| Sbry | 2.64 * | 1.728 | −0.788 | −0.061 | 39 | ||||||
| Sepi | −2.38 * | −1.749 | 2.194 | 14 | |||||||
| Slea | −3.59 * | 6.582 ** | 7 |
| sto-sed | sto-bry | sto-epi | sto-lea | sed-bry | sed-epi | sed-lea | bry-epi | bry-lea | all | |
|---|---|---|---|---|---|---|---|---|---|---|
| Jaccard index | 0.65 | 0.73 | 0.71 | 0.84 | 0.65 | 0.71 | 0.76 | 0.72 | 0.75 | 0.70 |
| Chao index | 0.30 | 0.36 | 0.46 | 0.72 | 0.33 | 0.41 | 0.42 | 0.35 | 0.52 | 0.37 |
| Z Value | ||||||
|---|---|---|---|---|---|---|
| Jaccard | Chao | |||||
| sto-sed | sto-bry | sed-bry | sto-sed | sto-bry | sed-bry | |
| Elevation | −1.587 | 1.534 | −0.313 | −2.581 ** | 1.213 | −1.019 |
| Cond | −0.034 | 3.101 ** | 2.387 * | −0.626 | 4.200 *** | 0.734 |
| Shading | 0.55 | −0.061 | 1.63 | −0.694 | −0.744 | 1.152 |
| pH | −0.186 | −0.853 | 1.153 | −0.373 | −3.383 *** | 1.248 |
| Flow | −1.682 | −1.573 | −5.817 *** | −2.798 ** | −2.705 ** | −3.086 ** |
| Temp | −0.209 | −0.319 | −3.694 *** | −1.286 | −1.457 | −1.611 |
| N | −0.358 | −0.381 | −2.156 * | −0.566 | −1.789 | −1.047 |
| P | 0.38 | 2.518 * | 2.933 ** | 1.014 | 4.151 *** | 2.171 * |
| Cl | −0.791 | −2.368 * | −0.021 | −0.59 | −3.592 *** | −0.954 |
| HD | −1.01 | 0.587 | −0.936 | −1.013 | −0.2 | −1.766 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taxböck, L.; Karger, D.N.; Kessler, M.; Spitale, D.; Cantonati, M. Diatom Species Richness in Swiss Springs Increases with Habitat Complexity and Elevation. Water 2020, 12, 449. https://doi.org/10.3390/w12020449
Taxböck L, Karger DN, Kessler M, Spitale D, Cantonati M. Diatom Species Richness in Swiss Springs Increases with Habitat Complexity and Elevation. Water. 2020; 12(2):449. https://doi.org/10.3390/w12020449
Chicago/Turabian StyleTaxböck, Lukas, Dirk Nikolaus Karger, Michael Kessler, Daniel Spitale, and Marco Cantonati. 2020. "Diatom Species Richness in Swiss Springs Increases with Habitat Complexity and Elevation" Water 12, no. 2: 449. https://doi.org/10.3390/w12020449
APA StyleTaxböck, L., Karger, D. N., Kessler, M., Spitale, D., & Cantonati, M. (2020). Diatom Species Richness in Swiss Springs Increases with Habitat Complexity and Elevation. Water, 12(2), 449. https://doi.org/10.3390/w12020449