Studying Hydraulic Interconnections in Low-Permeability Media by Using Bacterial Communities as Natural Tracers

 ,
,  ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
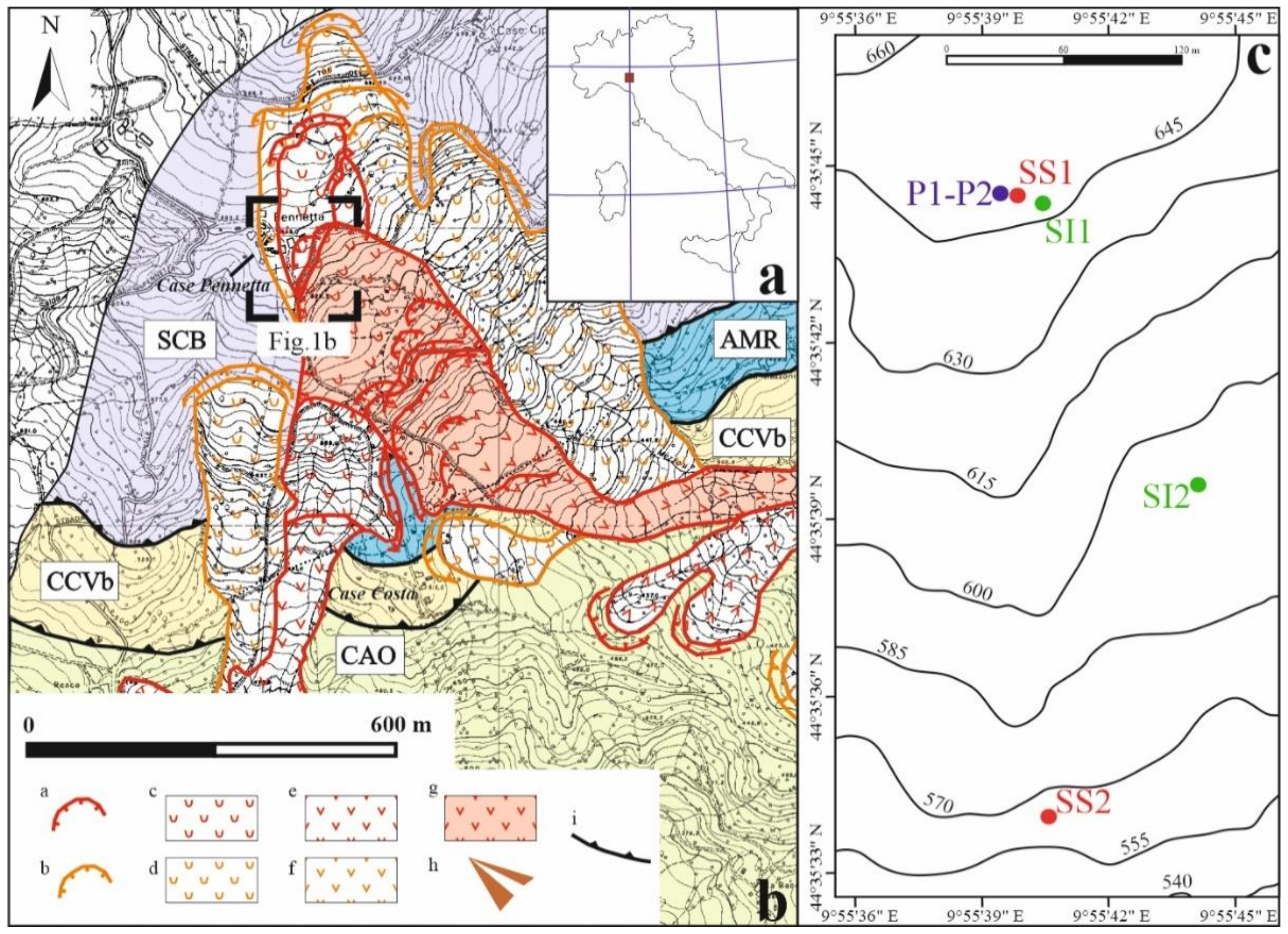
2.1. Test Site
2.2. Mineralogical Investigations
2.3. Hydrogeological Investigations
2.4. Next-Generation Sequencing (NGS) for Bacterial Community Analyses
3. Results
3.1. Mineralogical Investigations
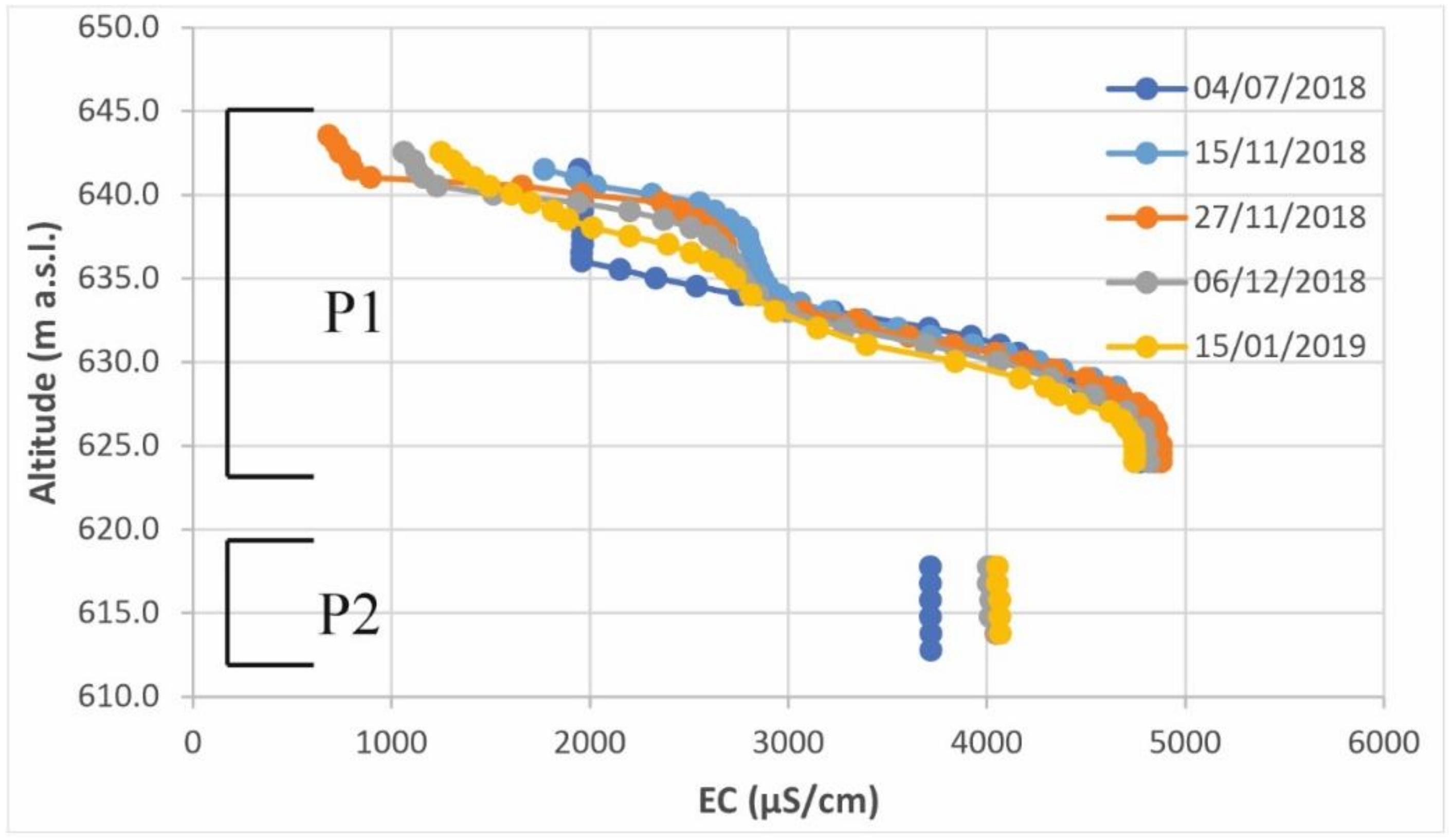
3.2. Hydrogeological Investigations
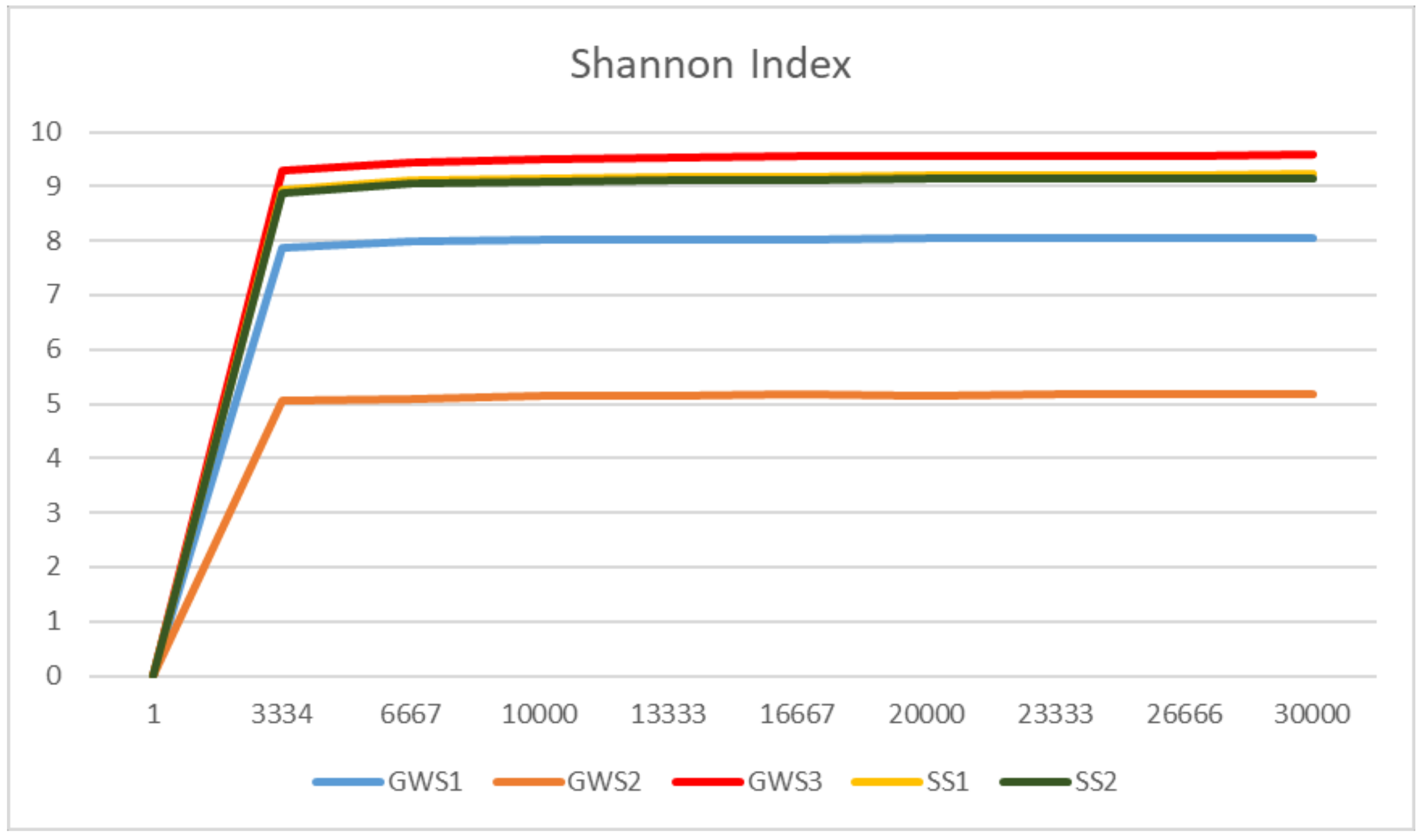
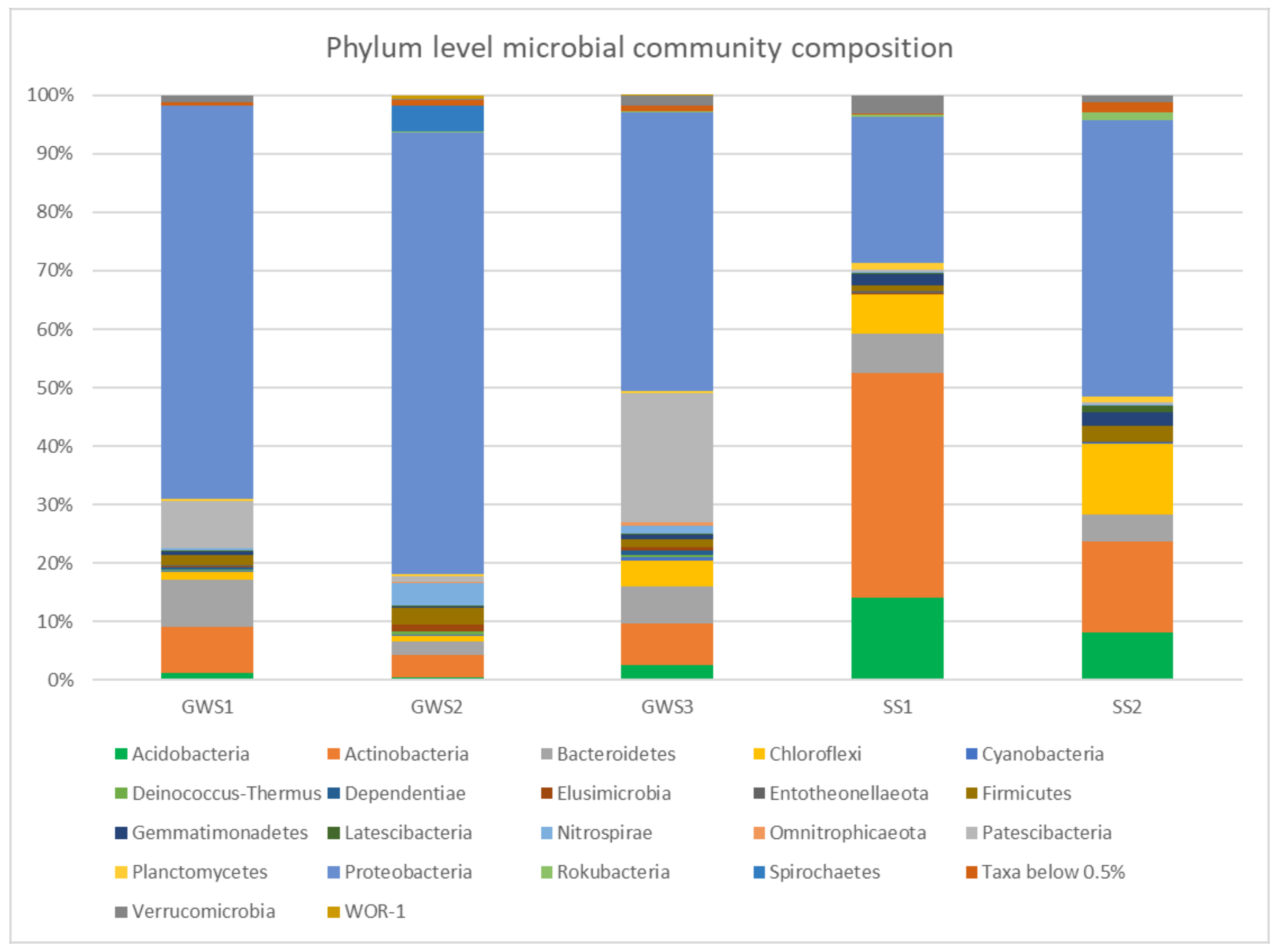
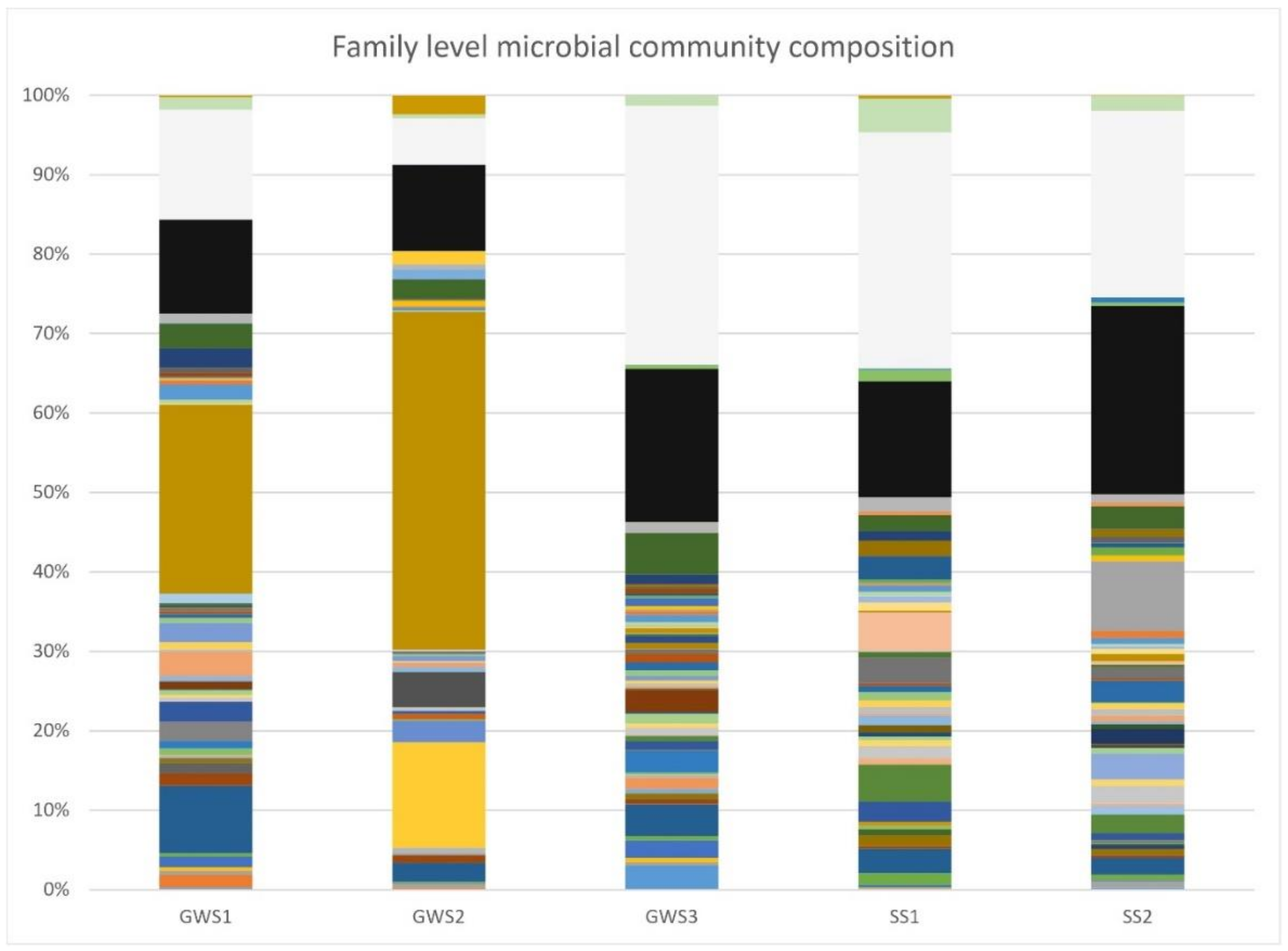
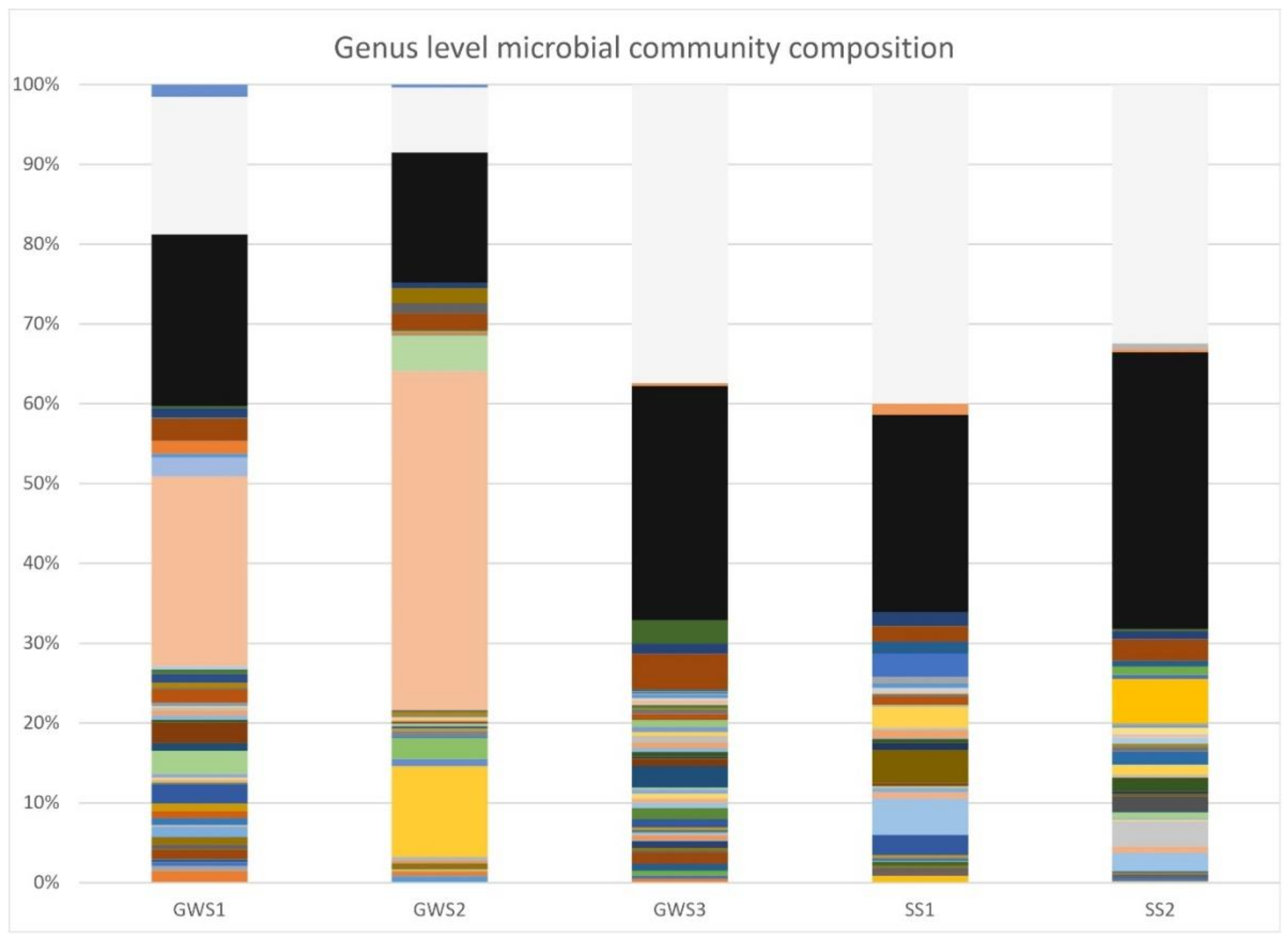
3.3. Groundwater and Soil Microbial Communities
4. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tufenkji, N.; Emelko, M.B. Fate and transport of microbial contaminants in groundwater. In Encyclopedia of Environmental Health; Nriagu, J.O., Ed.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 715–726. [Google Scholar]
- Bucci, A.; Petrella, E.; Celico, F.; Naclerio, G. Use of molecular approaches in hydrogeological studies: The case of carbonate aquifers in southern Italy. Hydrogeol. J. 2017, 25, 1017–1031. [Google Scholar] [CrossRef]
- Mills, A.L.; Herman, J.S.; Hornberger, G.M.; Dejesus, T.H. Effect of solution ionic-strength and iron coatings on mineral grains on the sorption of bacterial-cells to quartz sand. Appl. Environ. Microbiol. 1994, 60, 3300–3306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mccaulou, D.R.; Bales, R.C.; Arnold, R.G. Effect of temperature-controlled motility on transport of bacteria and microspheres through saturated sediment. Water Resour. Res. 1995, 31, 271–280. [Google Scholar] [CrossRef]
- Hendry, M.J.; Lawrence, J.R.; Maloszewski, P. Effects of velocity on the transport of two bacteria through saturated sand. Ground Water 1999, 37, 103–112. [Google Scholar] [CrossRef]
- Yee, N.; Fein, J.B.; Daughney, C.J. Experimental study of the pH, ionic strength, and reversibility behavior of bacteria-mineral adsorption. Geochim. Cosmochim. Acta 2000, 64, 609–617. [Google Scholar] [CrossRef]
- Dong, H.L.; Onstott, T.C.; Deflaun, M.F.; Fuller, M.E.; Scheibe, T.D.; Streger, S.H.; Rothmel, R.K.; Mailloux, B.J. Relative dominance of physical versus chemical effects on the transport of adhesion-deficient bacteria in intact cores from South Oyster, Virginia. Environ. Sci. Technol. 2002, 36, 891–900. [Google Scholar] [CrossRef]
- Bradford, S.A.; Šimůnek, J.; Walker, S.L. Transport and straining of E. coli O157: H7 in saturated porous media. Water Resour. Res. 2006, 42. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.X.; Walker, S.L. Role of solution chemistry and ion valence on the adhesion kinetics of groundwater and marine bacteria. Langmuir 2007, 23, 7162–7169. [Google Scholar] [CrossRef]
- Ochiai, N.; Kraft, E.L.; Selker, J.S. Methods for colloid transport visualization in pore networks. Water Resour. Res. 2006, 42. [Google Scholar] [CrossRef] [Green Version]
- Streit, W.R.; Schmitz, R.A. Metagenomics—The key to the uncultured microbes. Curr. Opin. Microbiol. 2004, 7, 492–498. [Google Scholar] [CrossRef]
- Kucharzyk, K.H.; Rectanus, H.V.; Bartling, C.M.; Rosansky, S.; Minard-Smith, A.; Mullins, L.A.; Neil, K. Use of omic tools to assess methyl tert-butyl ether (MTBE) degradation in groundwater. J. Hazard. Mater. 2019, 378, 120618. [Google Scholar] [CrossRef] [PubMed]
- Mosaddeghi, M.R.; Mahboubi, A.A.; Zandsalimi, S.; Unc, A. Influence of organic waste type and soil structure on the bacterial filtration rates in unsaturated intact soil columns. J. Environ. Manag. 2009, 90, 730–739. [Google Scholar] [CrossRef] [PubMed]
- Naclerio, G.; Fardella, G.; Marzullo, G.; Celico, F. Filtration of Bacillus subtilis and Bacillus cereus spores in a pyroclastic topsoil, carbonate Apennines, southern Italy. Colloids Surf. B Biointerfaces 2009, 70, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Safadoust, A.; Mahboubi, A.A.; Mosaddeghi, M.R.; Gharabaghi, B.; Voroney, P.; Unc, A.; Khodakaramian, G. Significance of physical weathering of two-texturally different soils for the saturated transport of Escherichia coli and bromide. J. Environ. Manag. 2012, 107, 147–158. [Google Scholar] [CrossRef]
- Moradi, A.; Mosaddeghi, M.R.; Chavoshi, E.; Safadoust, A.; Soleimani, M. Effect of Crude Oil-Induced Water Repellency on Transport of Escherichia coli and Bromide Through Repacked and Physically-Weathered Soil Columns. Environ. Pollut. 2019, 255, 113230. [Google Scholar] [CrossRef]
- Boschetti, T.; Falasca, A.; Bucci, A.; De Felice, V.; Naclerio, G.; Celico, F. Influence of soil on groundwater geochemistry in a carbonate aquifer, southern Italy. Int. J. Speleol. 2014, 43, 79–94. [Google Scholar] [CrossRef]
- Bucci, A.; Allocca, V.; Naclerio, G.; Capobianco, G.; Divino, F.; Fiorillo, F.; Celico, F. Winter survival of microbial contaminants in soil: An in situ verification. J. Environ. Sci. 2015, 27, 131–138. [Google Scholar] [CrossRef]
- Rizzo, P.; Bucci, A.; Sanangelantoni, A.M.; Iacumin, P.; Celico, F. Coupled Microbiological–Isotopic Approach for Studying Hydrodynamics in Deep Reservoirs: The Case of the Val d’Agri Oilfield (Southern Italy). Water 2020, 12, 1483. [Google Scholar] [CrossRef]
- Bucci, A.; Petrella, E.; Naclerio, G.; Gambatese, S.; Celico, F. Bacterial migration through low-permeability fault zones in compartmentalised aquifer systems: A case study in Southern Italy. Int. J. Speleol. 2014, 43, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Bucci, A.; Petrella, E.; Naclerio, G.; Allocca, V.; Celico, F. Microorganisms as contaminants and natural tracers: A 10-year research in some carbonate aquifers (southern Italy). Environ. Earth Sci. 2015, 74, 173–184. [Google Scholar] [CrossRef]
- Bucci, A.; Naclerio, G.; Allocca, V.; Celico, P.; Celico, F. Potential use of microbial community investigations to analyze hydrothermal systems behaviour: The case of Ischia island, southern Italy. Hydrol. Process. 2011, 25, 1866–1873. [Google Scholar] [CrossRef]
- Hernandez-Diaz, R.; Petrella, E.; Bucci, A.; Naclerio, G.; Feo, A.; Sferra, G.; Chelli, A.; Zanini, A.; Gonzales-Hernandez, P.; Celico, F. Integrating hydrogeological and microbiological data and modelling to characterize the hydraulic features and behavior of coastal carbonate aquifers: A case in Western Cuba. Water 2019, 11, 1989. [Google Scholar] [CrossRef] [Green Version]
- Vescovi, P.; Andreozzi, M.; De Nardo, M.T.; Lasagna, S.; Martelli, L.; Rio, D.; Tellini, C.; Vernia, L. Note illustrative della carta geologica d’Italia alla scala 1:50,000, foglio 216, Borgo Val di Taro. In Presidenza del Consigio dei Ministri, Servizio Geologico d’Italia, Regione Emilia-Romagna; S.El.Ca. SRLS.El.Ca. SRL: Florence, Italy, 2002. [Google Scholar]
- Remelli, S.; Petrella, E.; Chelli, A.; Conti, F.D.; Lozano Fondon, C.; Celico, F.; Francese, R.; Menta, C. Hydrodynamic and soil biodiversity characterization in an active landslide. Water 2019, 11, 1882. [Google Scholar] [CrossRef] [Green Version]
- Frouz, J. Use of soil dwelling Diptera (Insecta, Diptera) as bioindicators: A review of ecological requirements and response to disturbance. Agric. Ecosyst. Environ. 1999, 74, 167–186. [Google Scholar] [CrossRef]
- Edwards, C.A. The ecology of Symphyla. Entomol. Exp. Appl. 1958, 1, 308–319. [Google Scholar] [CrossRef]
- Menta, C.; Leoni, A.; Gardi, C.; Conti, F. Are grasslands important habitats for soil microarthropod conservation? Biodivers. Conserv. 2011, 20, 1073–1087. [Google Scholar] [CrossRef]
- Geoportale. Base Map: Technical Regional Map 1:5000, Emilia-Romagna Region 1998 (ed.). Available online: https://geoportale.regione.emilia-romagna (accessed on 23 June 2020).
- Morin, R.H.; Carleton, G.B.; Poirier, S. Fractured-aquifer hydrogeology from geophysical logs; the passaic formation, New Jersey. Ground Water 1997, 35, 328–338. [Google Scholar] [CrossRef] [Green Version]
- Cook, P.G.; Love, A.J.; Dighton, J.C. Inferring ground water flow in fractured rock from dissolved radon. Ground Water 1999, 37, 606–610. [Google Scholar] [CrossRef]
- Aquino, D.; Petrella, E.; Florio, M.; Celico, P.; Celico, F. Complex hydraulic interactions between compartmentalized carbonate aquifers and heterogeneous siliciclastic successions: A case study in southern Italy. Hydrol. Process. 2015, 29, 4252–4263. [Google Scholar] [CrossRef]
- Milani, C.; Hevia, A.; Foroni, E.; Duranti, S.; Turroni, F.; Lugli, G.A.; Sanchez, B.; Martín, R.; Gueimonde, M.; Van Sinderen, D.; et al. Assessing the fecal microbiota: An optimised ion torrent 16S rRNA gene-based analysis protocol. PLoS ONE 2013, 8, e68739. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, J.G. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Stackebrandt, E.; Koch, C.; Gvozdiak, O.; Schumann, P. Taxonomic Dissection of the Genus Micrococcus: Kocuria gen. nov., Nesterenkonia gen. nov., Kytococcus gen. nov., Dermacoccus gen. nov., and Micrococcus Cohn 1872 gen. emend. Int. J. Syst. Bacteriol. 1995, 45, 682–692. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.Q.; Chen, Y.G.; Li, W.J.; Tian, X.P.; Xu, L.H.; Jiang, C.L. Sphingomonas yunnanensis sp. nov., a novel Gram-negative bacterium from a contaminated plate. Int. J. Syst. Evol. Microbiol. 2005, 55, 2361–2364. [Google Scholar] [CrossRef]
- Baldani, J.I.; Rouws, L.; Cruz, L.M.; Olivares, F.L.; Schmid, M.; Hartmann, A. The family Oxalobacteraceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 919–974. [Google Scholar]
- Bernardet, J.F.; Bowman, J.P. The genus Flavobacterium. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 481–531. [Google Scholar]
- Berge, O.; Heulin, T.; Achouak, W.; Richard, C.; Bally, R.; Balandreau, J. Rahnella aquatilis, a nitrogen-fixing enteric bacterium associated with the rhizosphere of wheat and maize. Can. J. Microbiol. 1991, 37, 195–203. [Google Scholar] [CrossRef]
- Kaparullina, E.; Doronina, N.; Chistyakova, T.; Trotsenko, Y. Stenotrophomonas chelatiphaga sp. nov., a new aerobic EDTA-degrading bacterium. Syst. Appl. Microbiol. 2009, 32, 157–162. [Google Scholar] [CrossRef]
- Kloos, W.E.; Schleifer, K.H. Isolation and characterization of staphylococci from human skin II. descriptions of four new species: Staphylococcus warneri, Staphylococcus capitis, Staphylococcus hominis, and Staphylococcus simulans1. Int. J. Syst. Evol. Microbiol. 1975, 25, 62–79. [Google Scholar] [CrossRef] [Green Version]
- Higashioka, Y.; Kojima, H.; Watanabe, M.; Fukui, M. Desulfatitalea tepidiphila gen. nov., sp. nov., a sulfate-reducing bacterium isolated from tidal flat sediment. Int. J. Syst. Evol. Microbiol. 2013, 63, 761–765. [Google Scholar] [CrossRef]
- Fauque, G.D. Ecology of sulfate-reducing bacteria. In Sulfate-Reducing Bacteria; Barton, L.L., Ed.; Springer: Boston, MA, USA, 1995; Volume 8, pp. 217–241. [Google Scholar]
- Kojima, H.; Shinohara, A.; Fukui, M. Sulfurifustis variabilis gen. nov., sp nov., a sulfur oxidizer isolated from a lake, and proposal of Acidiferrobacteraceae fam. nov and Acidiferrobacterales ord. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 3709–3713. [Google Scholar] [CrossRef] [PubMed]
- Kao, P.M.; Tung, M.C.; Hsu, B.M.; Chiu, Y.C.; She, C.Y.; Shen, S.M.; Huang, Y.L.; Huang, W.C. Identification and quantitative detection of Legionella spp. in various aquatic environments by real-time PCR assay. Environ. Sci. Pollut. Res. Int. 2013, 20, 6128–6137. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.M.; Johnsen, K.; Sorensen, J.; Nielsen, P.; Jacobsen, C.S. Pseudomonas frederiksbergensis sp. nov., isolated from soil at a coal gasification site. Int. J. Syst. Evol. Microbiol. 2000, 50, 1957–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, J.H.; Kang, S.J.; Oh, H.W.; Oh, T.K. Pedobacter insulae sp. nov., isolated from soil. Int. J. Syst. Evol. Microbiol. 2007, 57, 1999–2003. [Google Scholar] [CrossRef] [Green Version]
- Rogel, M.A.; Hernandez-Lucas, I.; Kuykendall, L.D.; Balkwill, D.L.; Martinez-Romero, E. Nitrogen-fixing nodules with Ensifer adhaerens harboring Rhizobium tropici symbiotic plasmids. Appl. Environ. Microbiol. 2001, 67, 3264–3268. [Google Scholar] [CrossRef] [Green Version]
- Casida, L.E. Ensifer adhaerens gen. nov., sp. nov.: A bacterial predator of bacteria in soil. Int. J. Syst. Bacteriol. 1982, 32, 339–345. [Google Scholar] [CrossRef]
- Zhang, D.C.; Redzic, M.; Liu, H.C.; Zhou, Y.G.; Schinner, F.; Margesin, R. Devosia psychrophile sp. nov. and Devosia glacialis sp. nov., from alpine glacier cryoconite, and an emended description of the genus Devosia. Int. J. Syst. Evol. Microbiol. 2012, 62, 710–715. [Google Scholar] [CrossRef] [Green Version]
- La Scola, B.; Mallet, M.N.; Grimont, P.A.D.; Raoult, D. Description of Afipia birgiae sp. nov. and Afipia massiliensis sp. nov. and recognition of Afipia felis genospecies A. Int. J. Syst. Evol. Microbiol. 2002, 52, 1773–1782. [Google Scholar]
- Newton, H.J.; Ang, D.K.Y.; Van Driel, I.R.; Hartland, E. Molecular pathogenesis of infections caused by Legionella pneumophila. Clin. Microbiol. Rev. 2010, 23, 274–298. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Chuai, X.; Liang, M. Legionella feeleii: Pneumonia or Pontiac fever? Bacterial virulence traits and host immune response. Med. Microbiol. Immunol. 2019, 208, 25–32. [Google Scholar] [CrossRef]
- Kang, Y.S.; Jung, J.; Jeon, C.O.; Park, W. Acinetobacter oleivorans sp. nov. is capable of adhering to and growing on diesel-oil. J. Microbiol. 2011, 49, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Im, W.T.; Jung, H.M.; Ten, L.N.; Kim, M.K.; Bora, N.; Goodfellow, M.; Lim, S.; Jung, J.; Lee, S.T. Deinococcus aquaticus sp. nov., isolated from fresh water, and Deinococcus caeni sp. nov., isolated from activated sludge. Int. J. Syst. Evol. Microbiol. 2008, 58, 2348–2353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.C.; Jeon, C.O.; Lim, J.M.; Lee, S.M.; Lee, J.M.; Song, S.M.; Park, D.J.; Li, W.J.; Kim, C.J. Halomonas taeanensis sp. nov., a novel moderately halophilic bacterium isolated from a solar saltern in Korea. Int. J. Syst. Evol. Microbiol. 2005, 55, 2027–2032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Cánovas, M.J.; Quesada, E.; Llamas, I.; Bejar, V. Halomonas ventosae sp. nov., a moderately halophilic, denitrifying, exopolysaccharide-producing bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 733–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lincoln, S.P.; Fermor, T.R.; Tindall, B.J. Janthinobacterium agaricidamnosum sp. nov., a soft rot pathogen of Agaricus bisporus. Int. J. Syst. Evol. Microbiol. 1999, 49, 1577–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herwaldt, L.A.; Gorman, G.W.; McGrath, T.; Toma, S.; Brake, B.; Hightower, A.W.; Jones, J.; Reingold, A.L.; Boxer, P.A.; Tang, P.W. A new Legionella species, Legionella feeleii species nova, causes Pontiac fever in an automobile plant. Ann. Intern. Med. 1984, 100, 333–338. [Google Scholar] [CrossRef]
- Dennis, P.J.; Brenner, D.J.; Thacker, W.L.; Wait, R.; Vesey, G.; Steigerwalt, A.G.; Benson, R.F. Five new Legionella species isolated from water. Int. J. Syst. Evol. Microbiol. 1993, 43, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Orrison, L.H.; Cherry, W.B.; Fliermans, C.B.; Dees, S.B.; McDougal, L.K.; Dodd, D.J. Characteristics of environmental isolates of Legionella pneumophila. Appl. Environ. Microbiol. 1981, 42, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Clark, L.L.; Dajcs, J.J.; McLean, C.H.; Bartell, J.G.; Stroman, D.W. Pseudomonas otitidis sp. nov., isolated from patients with otic infections. Int. J. Syst. Evol. Microbiol. 2006, 56, 709–714. [Google Scholar] [CrossRef] [Green Version]
- Yumoto, I.; Kusano, T.; Shingyo, T.; Nodasaka, Y.; Matsuyama, H.; Okuyama, H. Assignment of Pseudomonas sp. strain E-3 to Pseudomonas psychrophila sp. nov., a new facultatively psychrophilic bacterium. Extremophiles 2001, 5, 343–349. [Google Scholar] [CrossRef]
- Romanenko, L.A.; Uchino, M.; Falsen, E.; Lysenko, A.M.; Zhukova, N.V.; Mikhailov, V.V. Pseudomonas xanthomarina sp. nov., a novel bacterium isolated from marine ascidian. J. Gen. Appl. Microbiol. 2005, 51, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Yandigeri, M.S.; Malviya, N.; Solanki, M.K.; Shrivastava, P.; Sivakumar, G. Chitinolytic Streptomyces vinaceusdrappus S5MW2 isolated from Chilika lake, India enhances plant growth and biocontrol efficacy through chitin supplementation against Rhizoctonia solani. World J. Microbiol. Biotechnol. 2015, 31, 1217–1225. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Li, W.J.; Jiao, W.C.; Li, Y.; Yuan, W.J.; Zhang, Y.Q.; Klenk, H.P.; Suh, J.W.; Bai, F.W. Streptomyces xinghaiensis sp. nov., isolated from marine sediment. Int. J. Syst. Evol. Microbiol. 2009, 59, 2870–2874. [Google Scholar] [CrossRef] [Green Version]
- Sorokin, D.Y.; Kovaleva, O.L.; Tourova, T.P.; Muyzer, G. Thiohalobacter thiocyanaticus gen. nov., sp. nov., a moderately halophilic, sulfur-oxidizing gammaproteobacterium from hypersaline lakes, that utilizes thiocyanate. Int. J. Syst. Evol. Microbiol. 2010, 60, 444–450. [Google Scholar] [CrossRef] [Green Version]
- Rutherford, D.W.; Chiou, C.T.; Eberl, D.D. Effects of exchanged cation on the microporosity of montmorillonite. Clays Clay Miner. 1997, 45, 534–543. [Google Scholar] [CrossRef]
- Kuila, U.; Prasad, M. Specific surface area and pore-size distribution in clays and shales. Geophys. Prospect. 2013, 61, 341–362. [Google Scholar] [CrossRef]
- Zaffar, M.; Lu, S.-G. Pore size distribution of clayey soils and its correlation with soil organic matter. Pedosphere 2015, 25, 240–249. [Google Scholar] [CrossRef]
- Aksu, I.; Bazilevskaya, E.; Karpyn, Z.T. Swelling of clay minerals in unconsolidated porous media and its impact on permeability. GeoResJ 2015, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Mueller, B. Experimental interactions between clay minerals and bacteria: A review. Pedosphere 2015, 25, 799–810. [Google Scholar] [CrossRef]
- Alimova, A.; Katz, A.; Steiner, N.; Rudolph, E.; Wei, H.; Steiner, J.C.; Gottlieb, P. Bacteria-clay interaction: Structural changes in smactite induced during biofilm formation. Clays Clay Miner. 2009, 57, 205–212. [Google Scholar] [CrossRef]
- Dexter, A.R. Heterogeneity of unsaturated, gravitational flow of water through beds of large particles. Water Resour. Res. 1993, 29, 1859–1862. [Google Scholar] [CrossRef]
- Wildenschild, D.; Jensen, K.H.; Villholth, K.; Illangasekare, T.H. A laboratory analysis of the effect of macropores on solute transport. Ground Water 1994, 32, 381–389. [Google Scholar] [CrossRef]
- McMurry, S.W.; Coyne, M.S.; Perfect, E. Fecal coliforms transport through intact soil blocks amended with poultry manure. J. Environ. Qual. 1998, 27, 86–92. [Google Scholar] [CrossRef] [Green Version]
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.; Freney, J.R.; Martinelli, L.A.; Seizinger, S.P.; Sutton, M.A. Transformation of the Nitrogen cycle: Recent trends, questions and potential solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef] [Green Version]
- Overeem, I.; Kettner, A.J.; Syvitski, J.P.M. 9.40 impacts of humans on river fluxes and morphology. In Treatise on Geomorphology; Shroder, J.F., Wohl, E., Eds.; Academic Press: Cambridge, MA, USA, 2013; pp. 828–842. [Google Scholar]
- Sutton, M.; Howard, C.; Erisman, J.W.; Billen, G.; Bleeker, A.; Grennfelt, P.; Van Grinsven, H.; Grizzetti, B. The European Nitrogen Assessment; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Barakata, M.; Cheviron, B.; Angulo-Jaramillo, R. Influence of the irrigation technique and strategies on the nitrogen cycle and budget: A review. Agric. Water Manag. 2016, 178, 225–238. [Google Scholar] [CrossRef]
- Racchetti, E.; Salmaso, F.; Pinardi, M.; Quadroni, S.; Soana, E.; Sacchi, E.; Severini, E.; Celico, F.; Viaroli, P.; Bartoli, M. Flood irrigation increases river-groundwater interactions and diffuse nitrate pollution in agricultural watersheds. Water 2019, 11, 2304. [Google Scholar] [CrossRef] [Green Version]
- Van Grinsven, H.J.M.; Ward, M.H.; Benjamin, N.; De Kok, T.M. Does the evidence about health risks associated with nitrate ingestion warrant an increase of the nitrate standard for drinking water? Environ. Health 2006, 5, 26. [Google Scholar] [CrossRef] [Green Version]
- Rivett, M.O.; Buss, S.R.; Morgan, P.; Smith, J.W.N.; Bemment, C.D. Nitrate attenuation in groundwater: A review of biogeochemical controlling processes. Water Res. 2008, 42, 4215–4232. [Google Scholar] [CrossRef]
- Howarth, R.; Chan, F.; Conley, D.J.; Garnier, J.; Doney, S.C.; Marino, R.; Billen, G. Coupled biogeochemical cycles: Eutrophication and hypoxia in temperate estuaries and coastal marine ecosystems. Front. Ecol. Environ. 2011, 9, 18–26. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Final Read Number |
|---|---|
| GWS1 | 46,615 |
| GWS2 | 94,558 |
| GWS3 | 56,571 |
| SS1 | 76,569 |
| SS2 | 61,852 |
| Taxonomy | Aerobic | Facultative Anaerobic | Halophilic/Halotolerant | Nitrate Reduction | Pathogen | Samples | Citations |
|---|---|---|---|---|---|---|---|
| Acinetobacter oleivorans | + | − | − | − | − | GWS1-2-3 | [57] |
| Afipia massiliensis | + | − | − | + | − | GWS1-2 | [54] |
| Deinococcus caeni | + | − | + | − | − | GWS1-2-3 | [58] |
| Devosia glacialis | + | − | − | + | − | GWS1-2-3; SS1 | [53] |
| Ensifer adhaerens | + | − | − | + | − | GWS1-2-3; SS1-2 | [52] |
| Halomonas taeanensis | + | − | + | + | − | SS2 | [59] |
| Halomonas ventosae | + | − | + | + | − | SS2 | [60] |
| Janthinobacterium agaricidamnosum | + | − | − | − | − | GWS1-3; SS1 | [61] |
| Legionella feeleii | + | − | − | − | + | GWS3; SS2 | [62] |
| Legionella nautarum | + | − | − | + | + | GWS3 | [63] |
| Legionella pneumophila | + | − | − | − | + | GWS3 | [64] |
| Pedobacter insulae | + | − | − | − | − | GWS1-2-3; SS1 | [50] |
| Pseudomonas frederiksbergensis | + | − | − | + | − | GWS1-2-3 | [49] |
| Pseudomonas otitidis | + | − | + | − | + | GWS2 | [65] |
| Pseudomonas psychrophila | + | − | + | + | − | GWS1-2-3; SS2 | [66] |
| Pseudomonas xanthomarina | + | − | + | + | − | GWS2 | [67] |
| Sphingomonas yunnanensis | + | − | − | + | − | GWS1-2-3; SS1-2 | [39] |
| Staphylococcus warneri | + | + | + | − | + | GWS2-3 | [44] |
| Stenotrophomonas chelatiphaga | + | − | − | − | − | GWS1-2; SS1 | [43] |
| Streptomyces vinaceusdrappus | + | − | + | + | − | GWS1-2-3; SS1-2 | [68] |
| Streptomyces xinghaiensis | + | − | + | − | − | GWS1-2-3; SS1-2 | [69] |
| Thiohalobacter thiocyanaticus | + | − | + | − | − | SS2 | [70] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rizzo, P.; Petrella, E.; Bucci, A.; Salvioli-Mariani, E.; Chelli, A.; Sanangelantoni, A.M.; Raimondo, M.; Quagliarini, A.; Celico, F. Studying Hydraulic Interconnections in Low-Permeability Media by Using Bacterial Communities as Natural Tracers. Water 2020, 12, 1795. https://doi.org/10.3390/w12061795
Rizzo P, Petrella E, Bucci A, Salvioli-Mariani E, Chelli A, Sanangelantoni AM, Raimondo M, Quagliarini A, Celico F. Studying Hydraulic Interconnections in Low-Permeability Media by Using Bacterial Communities as Natural Tracers. Water. 2020; 12(6):1795. https://doi.org/10.3390/w12061795
Chicago/Turabian StyleRizzo, Pietro, Emma Petrella, Antonio Bucci, Emma Salvioli-Mariani, Alessandro Chelli, Anna Maria Sanangelantoni, Melinda Raimondo, Andrea Quagliarini, and Fulvio Celico. 2020. "Studying Hydraulic Interconnections in Low-Permeability Media by Using Bacterial Communities as Natural Tracers" Water 12, no. 6: 1795. https://doi.org/10.3390/w12061795
APA StyleRizzo, P., Petrella, E., Bucci, A., Salvioli-Mariani, E., Chelli, A., Sanangelantoni, A. M., Raimondo, M., Quagliarini, A., & Celico, F. (2020). Studying Hydraulic Interconnections in Low-Permeability Media by Using Bacterial Communities as Natural Tracers. Water, 12(6), 1795. https://doi.org/10.3390/w12061795