Diatom Taxonomic Composition as a Biological Indicator of the Ecological Health and Status of a River Basin under Agricultural Influence




Abstract
:1. Introduction
2. Materials and Methods
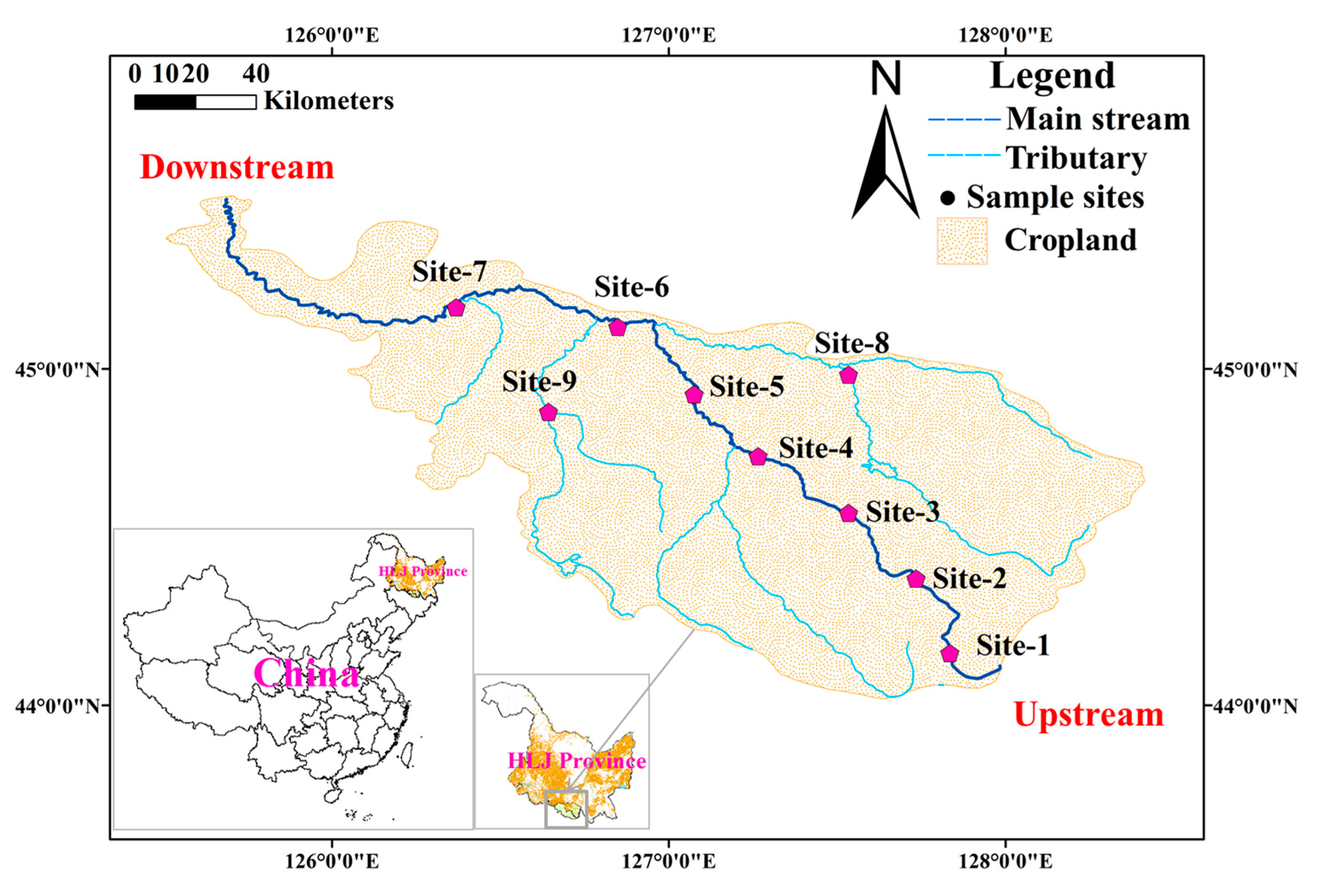
2.1. Study Area, Sample Collection and Laboratory Analyses
2.2. Multivariate Statistical Analysis
3. Results
3.1. Environmental Gradient of LLRB during Ice Cover
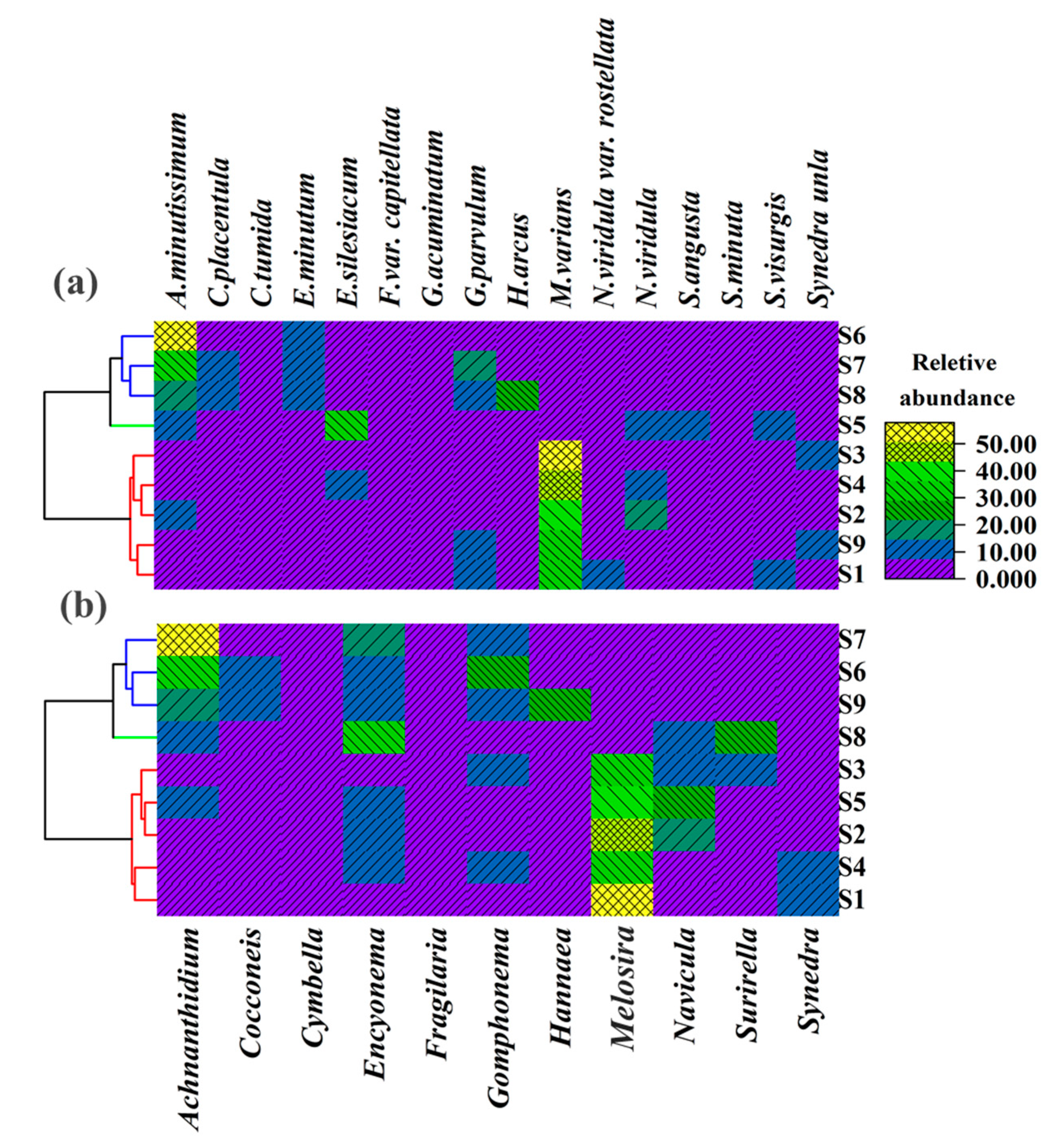
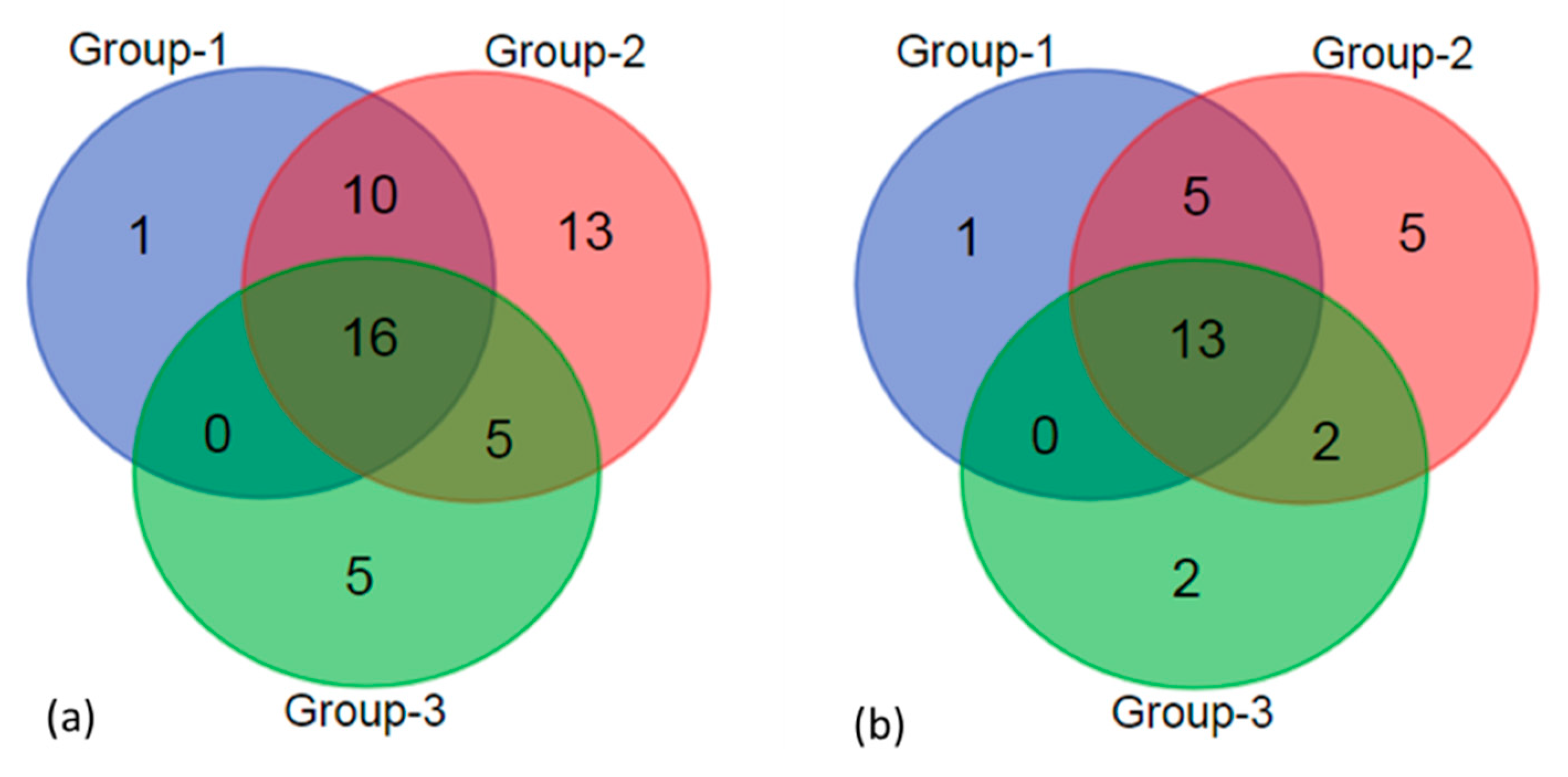
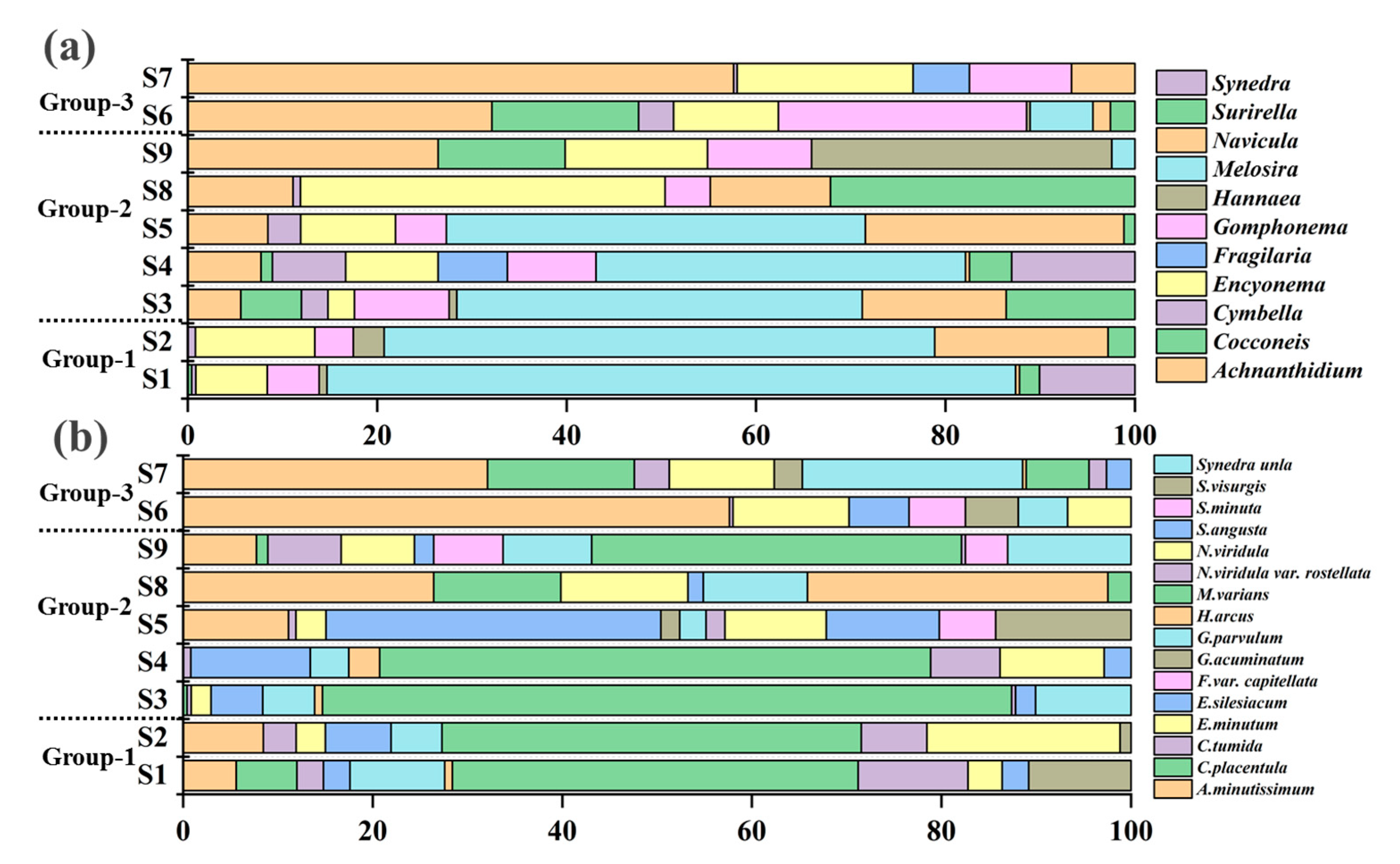
3.2. Diatom Taxonomic Composition at Genus And Species Levels
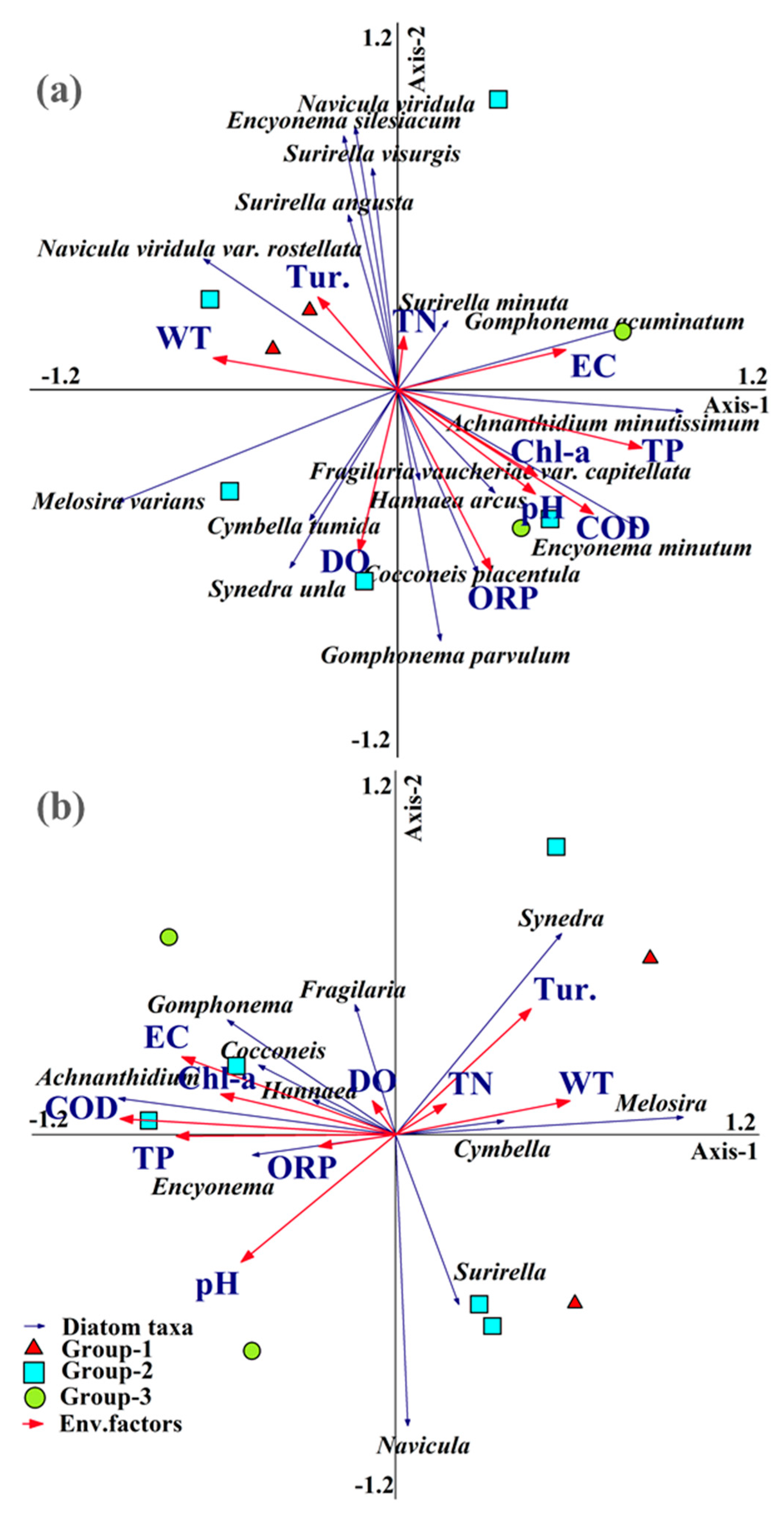
3.3. Diatom Taxonomic Composition Linked to Environmental Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, R.H.; Teng, Y.G.; Chen, H.Y.; Hu, B.; Yue, W.F. Groundwater pollution and risk assessment based on source apportionment in a typical cold agricultural region in Northeastern China. Sci. Total Environ. 2019, 696, 133972. [Google Scholar] [CrossRef] [PubMed]
- Hicks, M.B.; Taylor, J.M. Diatom Assemblage Changes in Agricultural Alluvial Plain Streams and Application for Nutrient Management. J. Environ. Qual. 2019, 48, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Bellinger, B.J.; Angradi, T.R.; Bolgrien, D.W.; Jicha, T.M.; Hill, B.H.; Reavie, E.D. Longitudinal variation and response to anthropogenic stress in diatom assemblages of the Lower Mississippi River, USA. River Syst. 2013, 21, 29–54. [Google Scholar] [CrossRef]
- Kamari, M.; Alho, P.; Veijalainen, N.; Aaltonen, J.; Huokuna, M.; Lotsari, E. River ice cover influence on sediment transportation at present and under projected hydroclimatic conditions. Hydrol. Process. 2015, 29, 4738–4755. [Google Scholar] [CrossRef]
- Keatley, B.E.; Douglas, M.S.V.; Smol, J.P. Prolonged ice cover dampens diatom community responses to recent climatic change in High Arctic lakes. Arct. Antarct. Alp. Res. 2008, 40, 364–372. [Google Scholar] [CrossRef] [Green Version]
- Walter, P. Environmental pollution Sea ice cover linked to toxic mercury. Chem. Ind. 2011, 3, 7-7. [Google Scholar]
- Binding, C.E.; Greenberg, T.A.; Bukata, R.P.; Smith, D.E.; Twiss, M.R. The MERIS MCI and its potential for satellite detection of winter diatom blooms on partially ice-covered Lake Erie. J. Plankton Res. 2012, 34, 569–573. [Google Scholar] [CrossRef] [Green Version]
- Katz, S.L.; Izmest’eva, L.R.; Hampton, S.E.; Ozersky, T.; Shchapov, K.; Moore, M.V.; Shimaraeva, S.V.; Silow, E.A. The “Melosira” years of Lake Baikal: Winter environmental conditions at ice onset predict under-ice algal blooms in spring. Limnol. Oceanogr. 2015, 60, 1950–1964. [Google Scholar] [CrossRef]
- Pardo, I.; Delgado, C.; Abrain, R.; Gomez-Rodriguez, C.; Garcia-Rosello, E.; Garcia, L.; Reynoldson, T.B. A predictive diatom-based model to assess the ecological status of streams and rivers of Northern Spain. Ecol. Indic. 2018, 90, 519–528. [Google Scholar] [CrossRef]
- Frenette, J.J.; Thibeault, P.; Lapierre, J.F.; Hamilton, P.B. Presence of algae in freshwater ice cover of fluvial LAC Saint-Pierre (St. Lawrence river, Canada). J. Phycol. 2008, 44, 284–291. [Google Scholar] [CrossRef]
- Chen, X.; McGowan, S.; Bu, Z.J.; Yang, X.D.; Cao, Y.M.; Bai, X.; Zeng, L.H.; Liang, J.; Qiao, Q.L. Diatom-based water-table reconstruction in Sphagnum peatlands of northeastern China. Water Res. 2020, 174, 115648. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.E.; Becker, T.J.; Bellucci, C.J. Diatom tolerance metrics to identify total phosphorus as candidate cause of aquatic life impairment in Connecticut, USA freshwater streams. Ecol. Indic. 2018, 93, 638–646. [Google Scholar] [CrossRef]
- Kim, H.K.; Cho, I.H.; Hwang, E.A.; Kim, Y.J.; Kim, B.H. Benthic Diatom Communities in Korean Estuaries: Species Appearances in Relation to Environmental Variables. Int. J. Environ. Res. Public Health 2019, 16, 2681. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Cao, J.X.; Pei, G.F.; Liu, G.X. Using benthic diatom assemblages to assess human impacts on streams across a rural to urban gradient. Environ. Sci. Pollut. Res. 2015, 22, 18093–18106. [Google Scholar] [CrossRef]
- Liu, Y.; Shen, J.M.; Chen, Z.L.; Ren, N.Q.; Li, Y.F. Distribution of polycyclic aromatic hydrocarbons in surface water and sediment near a drinking water reservoir in Northeastern China. Environ. Sci. Pollut. Res. 2013, 20, 2535–2545. [Google Scholar] [CrossRef]
- Chen, X.; Bu, Z.J.; Stevenson, M.A.; Cao, Y.M.; Zeng, L.H.; Qin, B. Variations in diatom communities at genus and species levels in peatlands (central China) linked to microhabitats and environmental factors. Sci. Total Environ. 2016, 568, 137–146. [Google Scholar] [CrossRef]
- Growns, I. Is genus or species identification of periphytic diatoms required to determine the impacts of river regulation? J. Appl. Phycol. 1999, 11, 273–283. [Google Scholar] [CrossRef]
- Xiao, W.; Huang, Y.; Mi, W.; Wu, H.; Bi, Y. Variation of Diatoms and Silicon in a Tributary of the Three Gorges Reservoir: Evidence of Interaction. Water 2019, 11, 1369. [Google Scholar] [CrossRef] [Green Version]
- Xue, H.; Zheng, B.; Meng, F.; Wang, Y.; Zhang, L.; Cheng, P. Assessment of Aquatic Ecosystem Health of the Wutong River Based on Benthic Diatoms. Water 2019, 11, 727. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.J.; Teng, Y.G.; Zhai, Y.Z.; Hu, L.T.; Zhao, X.B.; Zhang, M.L. Suitability for developing riverside groundwater sources along Songhua River, Northeast China. Hum. Ecol. Risk Assess. 2018, 24, 2088–2100. [Google Scholar] [CrossRef]
- Kim, H.K.; Kwon, Y.S.; Kim, Y.J.; Kim, B.H. Distribution of Epilithic Diatoms in Estuaries of the Korean Peninsula in Relation to Environmental Variables. Water 2015, 7, 6702–6718. [Google Scholar] [CrossRef] [Green Version]
- Wang, M. Evaluate method and classification standard on lake eutrophication. Environ. Monit. China 2002, 18, 47–49. [Google Scholar]
- Tan, X.; Ma, P.M.; Bunn, S.E.; Zhang, Q.F. Development of a benthic diatom index of biotic integrity (BD-IBI) for ecosystem health assessment of human dominant subtropical rivers, China. J. Environ. Manag. 2015, 151, 286–294. [Google Scholar] [CrossRef]
- Lange-Bertalot, H.; Cavacini, P.; Tagliaventi, N.; Alfinito, S. Diatoms of Sardinia. Rare and 76 New Species in Rock Pools and Other Ephemeral Waters; Iconographia Diatomologica 12; A.R.G. Gantner Verlag, KG: Ruggell, Germany, 2003; pp. 1–438. [Google Scholar]
- Lange-Bertalot, H.; Hofmann, G.; Werum, M.; Cantonati, M. Freshwater Benthic Diatoms of Central Europe:Over 800 Common Species Used in Ecological Assessment; English edition with updated taxonomy and addedspecies; Koeltz Botanical Books: Schmitten, Oberreifenberg, Germany, 2017; pp. 1–942. [Google Scholar]
- Wojtal, A.Z. Species Composition and Distribution of Diatom Assemblages in Spring Waters from Various Geological Formations in Southern Poland; Bibliotheca Diatomologica 59; J. Cramer: Stuttgart, Germany, 2013; pp. 1–436. [Google Scholar]
- Lai, J. Canoco 5: A new version of an ecological multivariate data ordination program. Biodivers. Sci. 2013, 21, 765–768. [Google Scholar]
- Gang, H.X.; Li, R.H.; Zhao, Y.M.; Liu, G.F.; Chen, S.; Jiang, J. Loss of GLK1 transcription factor function reveals new insights in chlorophyll biosynthesis and chloroplast development. J. Exp. Bot. 2019, 70, 3125–3138. [Google Scholar] [CrossRef]
- Twiss, M.R.; McKay, R.M.L.; Bourbonniere, R.A.; Bullerjahn, G.S.; Carrick, H.J.; Smith, R.E.H.; Winter, J.G.; D’Souza, N.A.; Furey, P.C.; Lashaway, A.R.; et al. Diatoms abound in ice-covered Lake Erie: An investigation of offshore winter limnology in Lake Erie over the period 2007 to 2010. J. Great Lakes Res. 2012, 38, 18–30. [Google Scholar] [CrossRef]
- Lotter, A.F.; Bigler, C. Do diatoms in the Swiss Alps reflect the length of ice-cover? Aquat. Sci. 2000, 62, 125–141. [Google Scholar] [CrossRef] [Green Version]
- Pei, G.F.; Liu, G.X.; Hu, Z.Y. Variation of benthic diatoms in a shallow, eutrophic Chinese Lake, Lake Donghu. J. Freshw. Ecol. 2006, 21, 277–284. [Google Scholar] [CrossRef]
- Soares, M.C.S.; Huszar, V.L.M.; Roland, F. Phytoplankton dynamics in two tropical rivers with different degrees of human impact (southeast Brazil). River Res. Appl. 2007, 23, 698–714. [Google Scholar] [CrossRef]
- Bona, F.; La Morgia, V.; Falasco, E. Predicting river diatom removal after shear stress induced by ice melting. River Res. Appl. 2012, 28, 1289–1298. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.X.; Wang, Z.H.; Guo, X.; Gu, Y.G.; Liang, W.B.; Liu, L. Impacts of metal contamination and eutrophication on dinoflagellate cyst assemblages along the Guangdong coast of southern China. Mar. Pollut. Bull. 2017, 120, 239–249. [Google Scholar] [CrossRef]
- Agawin, N.S.R.; Duarte, C.M.; Agusti, S.; Vaque, D. Effect of N: P ratios on response of Mediterranean picophytoplankton to experimental nutrient inputs. Aquat. Microb. Ecol. 2004, 34, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Wood, R.J.; Mitrovic, S.M.; Lim, R.P.; Warne, M.S.; Dunlop, J.; Kefford, B. Benthic diatoms as indicators of herbicide toxicity in rivers—A new SPEcies At Risk (SPEAR(herbicides)) index. Ecol. Indic. 2019, 99, 203–213. [Google Scholar] [CrossRef]
- Tan, X.; Zhang, Q.F.; Burford, M.A.; Sheldon, F.; Bunn, S.E. Benthic Diatom Based Indices for Water Quality Assessment in Two Subtropical Streams. Front. Microbiol. 2017, 8, 601. [Google Scholar] [CrossRef] [Green Version]
- Maslennikova, A.V. Development and application of an electrical conductivity transfer function, using diatoms from lakes in the Urals, Russia. J. Paleolimnol. 2020, 63, 129–146. [Google Scholar] [CrossRef]
- Tibby, J.; Gell, P.A.; Fluin, J.; Sluiter, I.R.K. Diatom-salinity relationships in wetlands: Assessing the influence of salinity variability on the development of inference models. Hydrobiologia 2007, 591, 207–218. [Google Scholar] [CrossRef]
- Pestryakova, L.A.; Herzschuh, U.; Gorodnichev, R.; Wetterich, S. The sensitivity of diatom taxa from Yakutian lakes (north-eastern Siberia) to electrical conductivity and other environmental variables. Polar Res. 2018, 37, 1485625. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.; Lee, H.; Kim, Y.K.; Sohn, K.; Lee, K. Hydrogen peroxide interference in chemical oxygen demand during ozone based advanced oxidation of anaerobically digested livestock wastewater. Int. J. Environ. Sci. Technol. 2011, 8, 381–388. [Google Scholar] [CrossRef] [Green Version]
- Zubaidah, T.; Karnaningroem, N.; Slamet, A. The Self-Purification Ability in the Rivers of Banjarmasin, Indonesia. J. Ecol. Eng. 2019, 20, 177–182. [Google Scholar] [CrossRef]
- Xiao, C.G.; Chen, J.; Chen, D.; Chen, R.D. Effects of river sinuosity on the self-purification capacity of the Shiwuli River, China. Water Sci. Technol. Water Supply 2019, 19, 1152–1159. [Google Scholar] [CrossRef]
- Wiese, R.; Renaudie, J.; Lazarus, D.B. Testing the accuracy of genus-level data to predict species diversity in Cenozoic marine diatoms. Geology 2016, 44, 1051–1054. [Google Scholar] [CrossRef]
- Lazarus, D.; Barron, J.; Renaudie, J.; Diver, P.; Turke, A. Cenozoic Planktonic Marine Diatom Diversity and Correlation to Climate Change. PLoS ONE 2014, 9, e84857. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stations | WT (°C) | DO (mg/L) | EC (μS/cm) | pH | ORP (mV) | Tur. (NTU) | Chl-a (μg/L) | TN (mg/L) | TP (mg/L) | COD (mg/L) |
|---|---|---|---|---|---|---|---|---|---|---|
| S1 | 1.44 | 1.55 | 54.47 | 8.57 | 405 | 27.37 | 0.58 | 3.60 | 0.06 | 5.67 |
| S2 | 0.16 | 2.20 | 61.73 | 8.23 | 404 | 26.77 | 0.37 | 3.63 | 0.05 | 7.33 |
| S3 | 3.54 | 1.70 | 164.5 | 8.38 | 394 | 26.53 | 1.71 | 5.00 | 0.22 | 10.67 |
| S4 | 1.44 | 1.72 | 168.2 | 7.66 | 409 | 27.1 | 1.17 | 4.93 | 0.29 | 17.00 |
| S5 | 0.97 | 0.46 | 194.6 | 8.17 | 406 | 27.57 | 1.22 | 3.37 | 0.44 | 16.67 |
| S6 | 0.06 | 1.94 | 263.5 | 9.27 | 410 | 23.87 | 2.52 | 5.03 | 1.67 | 72.00 |
| S7 | 0.49 | 1.54 | 267.63 | 8.42 | 415 | 27.93 | 6.51 | 8.03 | 1.77 | 83.33 |
| S8 | 0.49 | 2.09 | 270.47 | 8.44 | 436 | 26.57 | 0.83 | 2.07 | 0.58 | 20.67 |
| S9 | 0.03 | 1.43 | 34.07 | 9.00 | 449 | 24.67 | 1.09 | 1.30 | 0.34 | 67.67 |
| T-test | ||||||||||
| Group-1 × Group-2 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p < 0.05 | p > 0.05 |
| Group-1 × Group-3 | p > 0.05 | p > 0.05 | p < 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p < 0.05 | p < 0.05 |
| Group-2 × Group-3 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p < 0.05 | p < 0.05 |
| TSI (Chl-a) | TSI (TN) | TSI (TP) | TSI (COD) | TSI (∑) | Trophic Status | |
|---|---|---|---|---|---|---|
| S1 | 19 | 76 | 49 | 47 | 48 | mesotrophic |
| S2 | 14 | 76 | 46 | 54 | 48 | mesotrophic |
| S3 | 31 | 82 | 70 | 64 | 62 | middle eutrophic |
| S4 | 27 | 82 | 74 | 76 | 65 | middle eutrophic |
| S5 | 27 | 75 | 81 | 76 | 65 | middle eutrophic |
| S6 | 35 | 82 | 103 | 115 | 84 | high eutrophic |
| S7 | 45 | 90 | 104 | 119 | 89 | high eutrophic |
| S8 | 23 | 67 | 85 | 82 | 64 | middle eutrophic |
| S9 | 26 | 59 | 77 | 113 | 69 | middle eutrophic |
| Environmental Factors | r2 | p | ||||
|---|---|---|---|---|---|---|
| Species-Level | Genus-Level | Species-Level | Genus-Level | |||
| Axis-1 | Axis-2 | Axis-1 | Axis-2 | |||
| WT | −0.601 | 0.104 | 0.575 | 0.111 | 0.260 | 0.168 |
| DO | −0.127 | −0.472 | −0.075 | 0.112 | 0.422 | 0.912 |
| EC | 0.549 | 0.130 | −0.674 | −0.037 | 0.278 | 0.038 |
| pH | 0.449 | −0.340 | −0.509 | −0.419 | 0.232 | 0.204 |
| ORP | 0.306 | −0.482 | −0.253 | 0.257 | 0.298 | 0.816 |
| Tur. | −0.260 | 0.304 | 0.448 | 0.416 | 0.362 | 0.150 |
| Chl-a | 0.455 | −0.281 | −0.576 | 0.133 | 0.540 | 0.220 |
| TN | 0.020 | 0.175 | 0.167 | 0.102 | 0.954 | 0.832 |
| TP | 0.674 | −0.190 | −0.724 | −0.004 | 0.004 | 0.030 |
| COD | 0.638 | −0.406 | −0.793 | 0.053 | 0.076 | 0.002 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, X.; Liu, Y.; Fan, Y. Diatom Taxonomic Composition as a Biological Indicator of the Ecological Health and Status of a River Basin under Agricultural Influence. Water 2020, 12, 2067. https://doi.org/10.3390/w12072067
Lu X, Liu Y, Fan Y. Diatom Taxonomic Composition as a Biological Indicator of the Ecological Health and Status of a River Basin under Agricultural Influence. Water. 2020; 12(7):2067. https://doi.org/10.3390/w12072067
Chicago/Turabian StyleLu, Xinxin, Yan Liu, and Yawen Fan. 2020. "Diatom Taxonomic Composition as a Biological Indicator of the Ecological Health and Status of a River Basin under Agricultural Influence" Water 12, no. 7: 2067. https://doi.org/10.3390/w12072067
APA StyleLu, X., Liu, Y., & Fan, Y. (2020). Diatom Taxonomic Composition as a Biological Indicator of the Ecological Health and Status of a River Basin under Agricultural Influence. Water, 12(7), 2067. https://doi.org/10.3390/w12072067