Nutrient Release Dynamics Associated with Native and Invasive Leaf Litter Decomposition: A Mesocosm Experiment


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
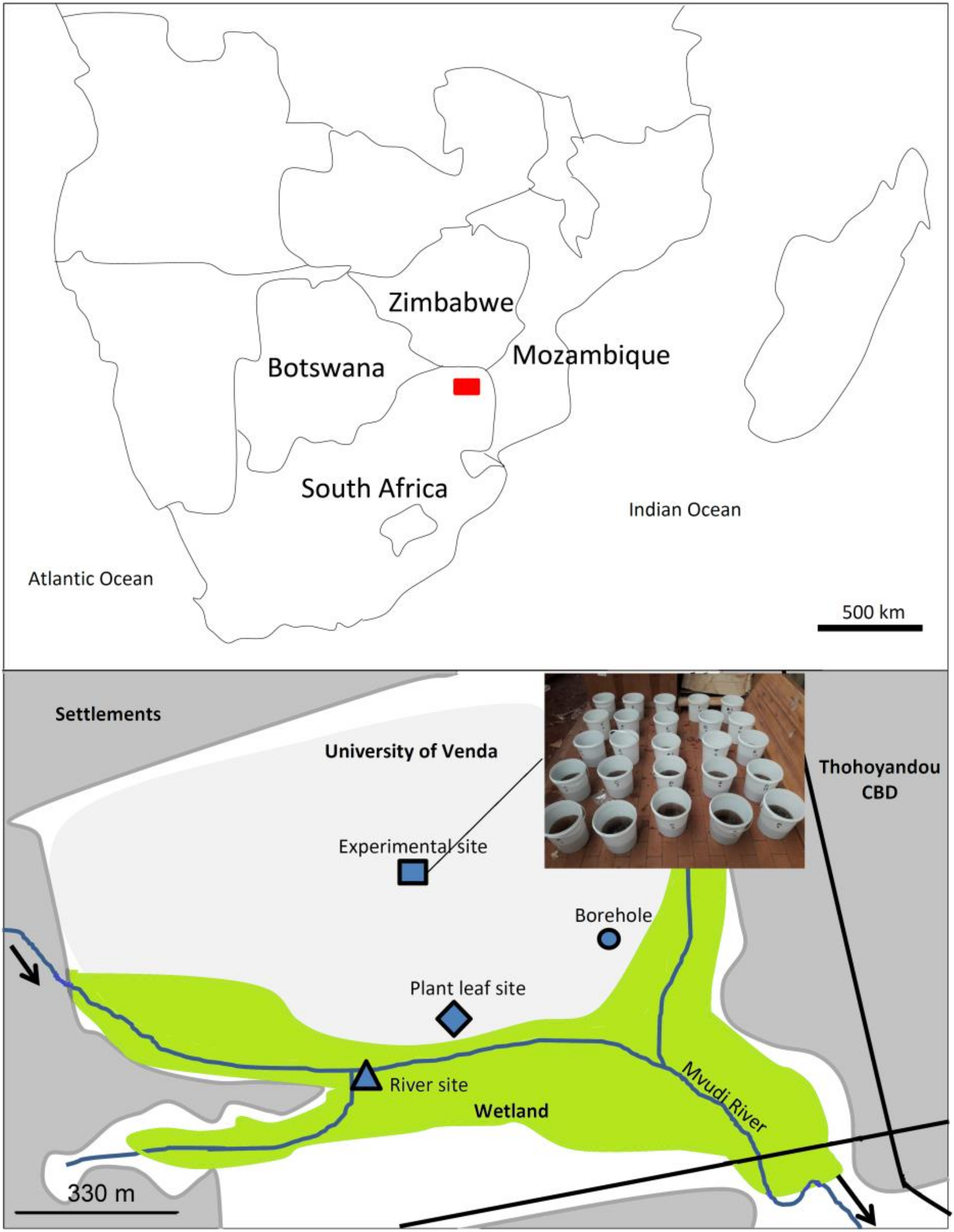
2.1. Study Area
2.2. Experimental Design
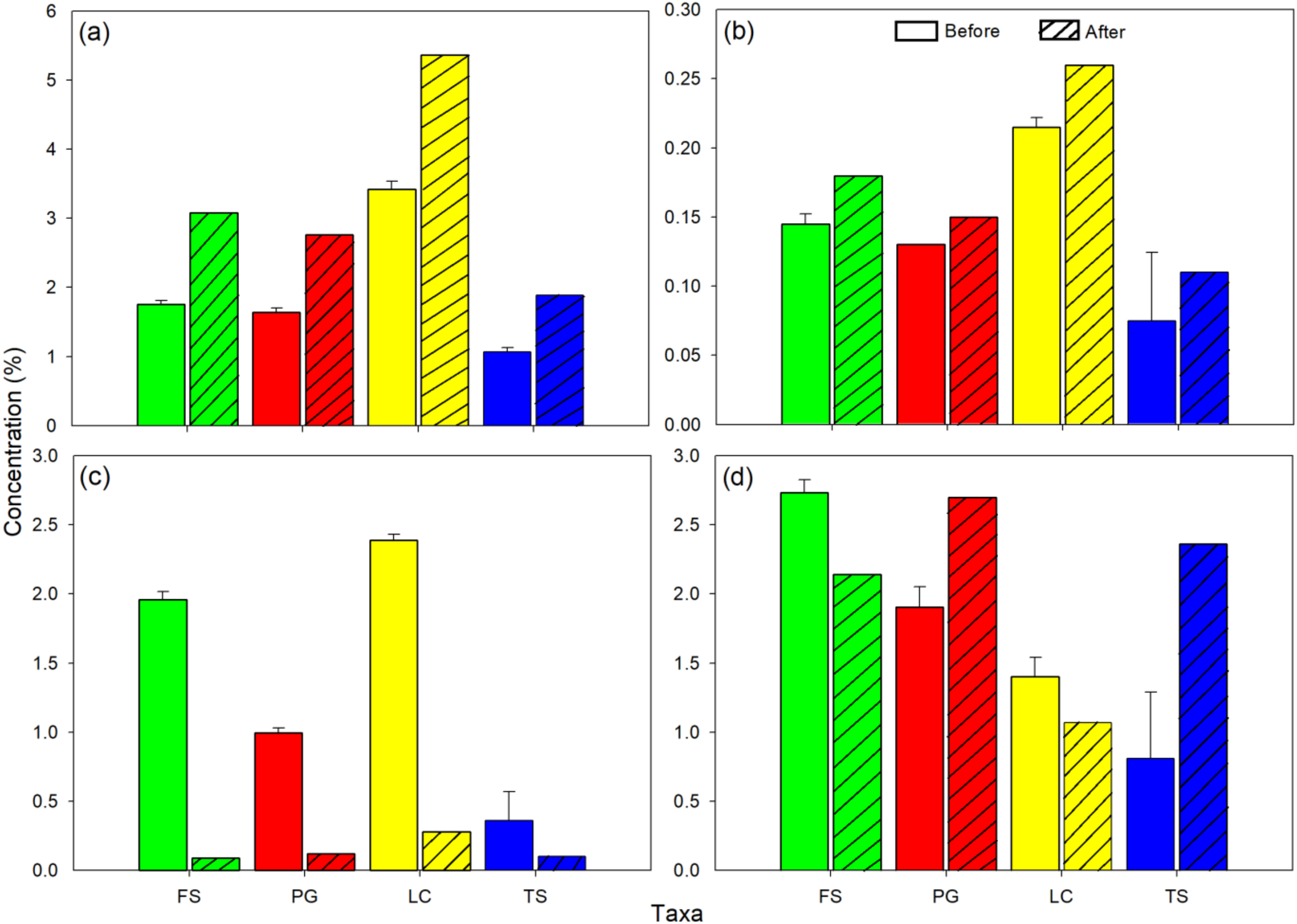
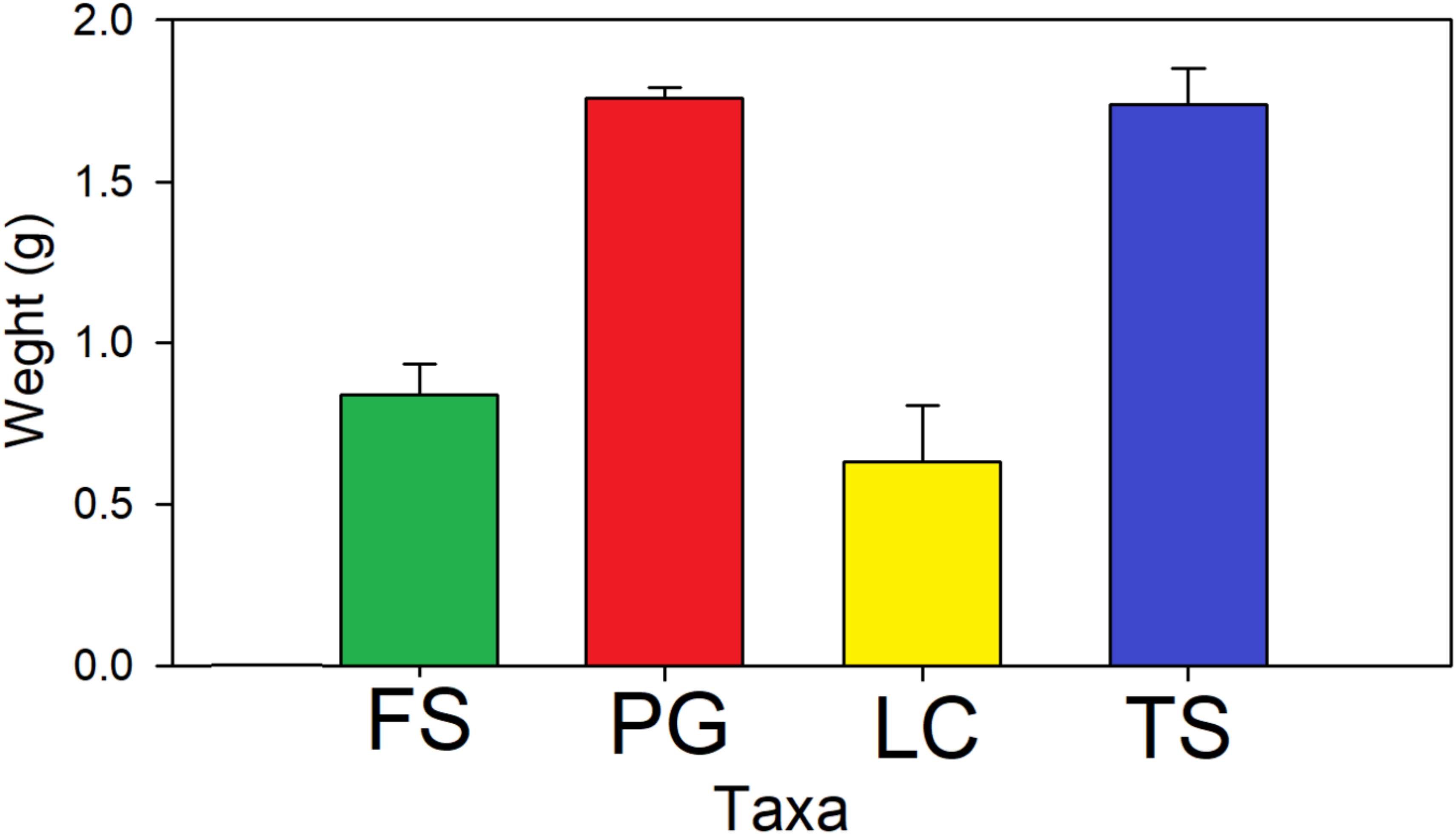
2.3. Sampling and Analyses
2.4. Statistical Analyses
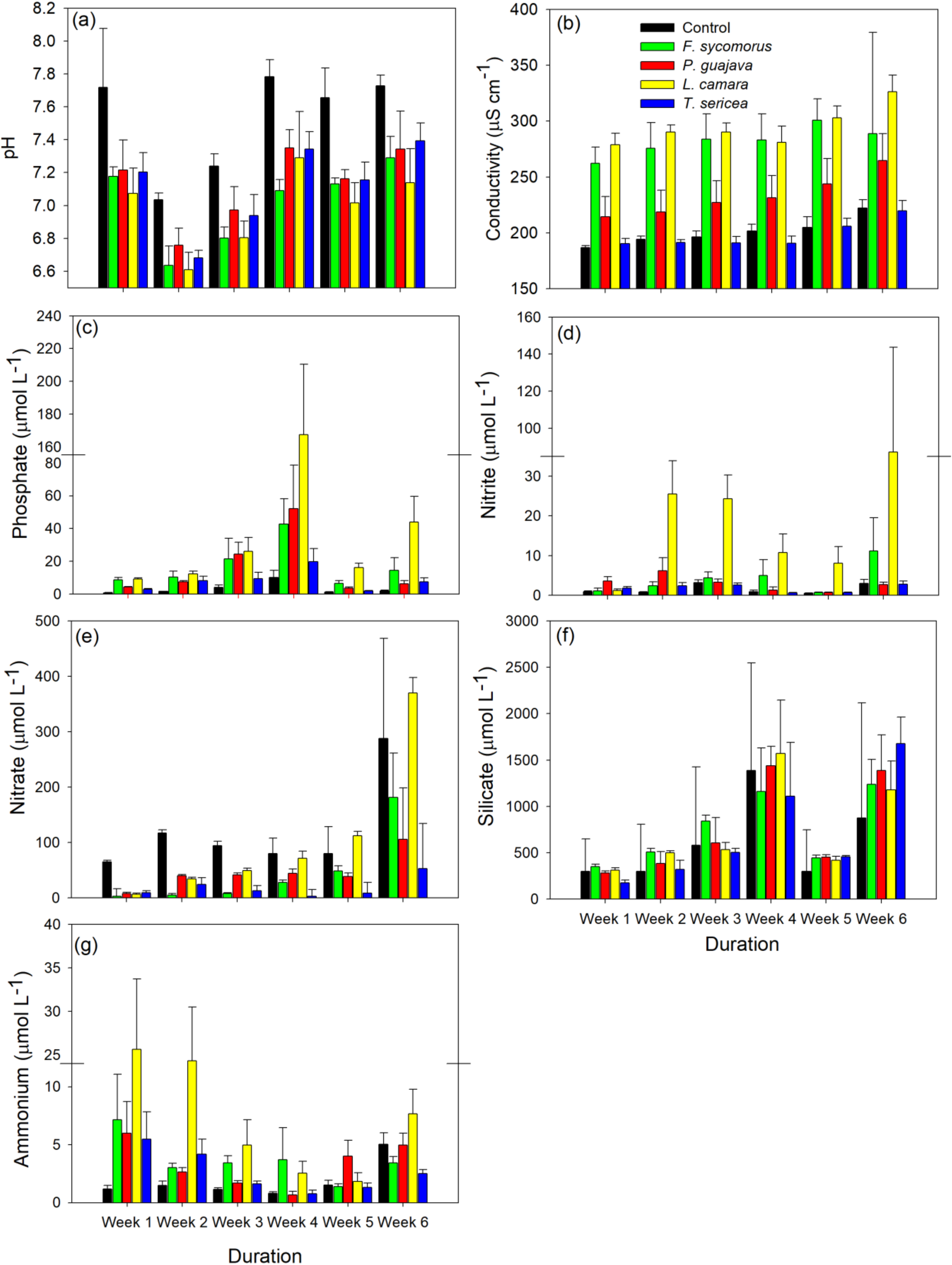
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vogt, K.A.; Grier, C.C.; Vogt, D.J. Production, turnover, and nutrient dynamics of above—And belowground detritus of world forests. Adv. Ecol. Res. 1986, 15, 303–377. [Google Scholar]
- DeGasparro, S.L.; Beresford, D.V.; Prater, C.; Frost, P.C. Leaf litter decomposition in boreal lakes: Variable mass loss and nutrient release ratios across a geographic gradient. Hydrobiologia 2020, 847, 819–830. [Google Scholar] [CrossRef]
- Aber, J.D.; Melillo, J.M. Nitrogen immobilization in decaying hardwood leaf litter as a function of initial nitrogen and lignin content. Can. J. Bot. 1982, 60, 2263–2269. [Google Scholar] [CrossRef]
- Zhang, D.; Hui, D.; Luo, Y.; Zhou, G. Rates of litter decomposition in terrestrial ecosystems: Global patterns and controlling factors. J. Plant Ecol. 2008, 1, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Bruder, A.; Schindler, M.H.; Moretti, M.S.; Gessner, M.O. Litter decomposition in a temperate and a tropical stream: The effects of species mixing, litter quality and shredders. Freshwater Biol. 2014, 59, 438–449. [Google Scholar] [CrossRef]
- Riggs, C.E.; Hobbie, S.E.; Cavender–Bares, J.; Savage, J.A.; Wei, X. Contrasting effects of plant species traits and moisture on the decomposition of multiple litter fractions. Oecologia 2015, 179, 573–584. [Google Scholar] [CrossRef]
- Chen, Y.; Ma, S.; Jiang, H.; Yangzom, D.; Cheng, G.; Lu, X. Decomposition time, chemical traits and climatic factors determine litter–mixing effects on decomposition in an alpine steppe ecosystem in Northern Tibet. Plant Soil 2019, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Krishna, M.P.; Mohan, M. Litter decomposition in forest ecosystems: A review. Energy Ecol. Environ. 2017, 2, 236–249. [Google Scholar] [CrossRef]
- Dilly, O.; Bartsch, S.; Rosenbrock, P.; Buscot, F.; Munch, J.C. Shifts in physiological capabilities of the microbiota during the decomposition of leaf litter in a black alder (Alnus glutinosa (Gaertn.) L.) forest. Soil Biol. Biochem. 2001, 33, 921–930. [Google Scholar] [CrossRef]
- Fernández, V.; Guzmán-Delgado, P.; Graça, J.; Santos, S.; Gil, L. Cuticle structure in relation to chemical composition: Re-assessing the prevailing model. Front. Plant Sci. 2016, 7, 427. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, B.E.; Klironomos, J.N. Breaking new ground: Soil communities and exotic plant invasion. BioScience 2005, 55, 477–487. [Google Scholar] [CrossRef]
- Drenovsky, R.E.; Martin, C.E.; Falasco, M.R.; James, J.J. Variation in resource acquisition and utilization traits between native and invasive perennial forbs. Am. J. Bot. 2008, 95, 681–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leishman, M.R.; Thomson, V.P.; Cooke, J. Native and exotic invasive plants have fundamentally similar carbon capture strategies. J. Ecol. 2010, 98, 28–42. [Google Scholar] [CrossRef]
- Webster, J.R.; Benfield, E.F. Vascular plant breakdown in freshwater ecosystems. Ann. Rev. Ecol. Syst. 1986, 17, 567–594. [Google Scholar] [CrossRef]
- McGroddy, M.E.; Daufresne, T.; Hedin, L.O. Scaling of C: N: P stoichiometry in forests worldwide: Implications of terrestrial Redfield-type ratios. Ecology 2004, 85, 2390–2401. [Google Scholar] [CrossRef]
- Brinson, M.M. Decomposition and nutrient exchange of litter in an alluvial swamp forest. Ecology 1977, 58, 601–609. [Google Scholar] [CrossRef]
- Richardson, D.M.; Van Wilgen, B.W. Invasive alien plants in South Africa: How well do we understand the ecological impacts? Working for water. S. Afr. J. Sci. 2004, 100, 45–52. [Google Scholar]
- Vardien, W.; Richardson, D.M.; Foxcroft, L.C.; Thompson, G.D.; Wilson, J.R.U.; Le Roux, J.J. Invasion dynamics of Lantana camara L. (sensu lato) in South Africa. S. Afr. J. Bot. 2012, 81, 81–94. [Google Scholar] [CrossRef] [Green Version]
- Urquía, D.; Gutierrez, B.; Pozo, G.; Pozo, M.J.; Espín, A.; de Lourdes Torres, M. Psidium guajava in the Galapagos Islands: Population genetics and history of an invasive species. PLoS ONE 2019, 14, e0203737. [Google Scholar] [CrossRef] [Green Version]
- Ramaswami, G.; Sukumar, R. Lantana camara L. (Verbenaceae) invasion along streams in a heterogeneous landscape. J. Biosci. 2014, 39, 717–726. [Google Scholar] [CrossRef]
- Pothasin, P.; Compton, S.G.; Wangpakapattanawong, P. Riparian Ficus tree communities: The distribution and abundance of riparian fig trees in Northern Thailand. PLoS ONE 2014, 9, 108945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venter, F.; Venter, J.A. Making the Most of Indigenous Trees; Briza Publications: Pretoria, South Africa, 2015. [Google Scholar]
- Sunil, C.; Somashekar, R.K.; Nagaraja, B.C. Diversity and composition of riparian vegetation across forest and agroecosystem landscapes of River Cauvery, southern India. Trop. Ecol. 2016, 57, 343–354. [Google Scholar]
- Mutshekwa, T.; Cuthbert, R.N.; Wasserman, R.J.; Murungweni, F.M.; Dalu, T. Macroinvertebrate colonisation associated with native and invasive leaf litter decomposition. Knowl. Manag. Aquat. Ecosyst. 2019, 421, 32. [Google Scholar]
- Staaf, H. Plant nutrient changes in beech leaves during senescence as influenced by site characteristics. Acta Oecol. 1982, 3, 161–170. [Google Scholar]
- Cuthbert, R.N.; Dalu, T.; Mutshekwa, T.; Wasserman, R.J. Leaf inputs from invasive and native plants drive differential mosquito abundances. Sci. Total Environ. 2019, 689, 652–654. [Google Scholar] [CrossRef]
- Grasshoff, K.; Ehrhardt, M.; Kremling, K. Methods of Seawater Analysis, 2nd ed.; Verlag Chemie: Weinheim, Germany, 1983. [Google Scholar]
- Armstrong, F.A.J.; Sterns, C.R.; Strickland, J.D.H. The measurement of upwelling and subsequent biological processes by means of the Technicon AutoAnalyzer and associated equipment. Deep Sea Res. 1967, 14, 381–389. [Google Scholar]
- Campbell, C.A.; Plank, C.O. Preparation of plant tissues for laboratory analysis. In Handbook of Reference Methods for Plant Analysis; Kalra, Y.P., Ed.; CRC Press: Boca Raton, FL, USA, 1998. [Google Scholar]
- Miller, M.K. Decomposition of bulk metallic glasses. Mat. Sci. Eng. A Struct. 1998, 250, 133–140. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R.V. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2018, 69, 1–33. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Core Development Team: Vienna, Austria, 2018. [Google Scholar]
- McNeish, R.E.; Benbow, M.E.; McEwan, R.W. Riparian forest invasion by a terrestrial shrub (Lonicera maackii) impacts aquatic biota and organic matter processing in headwater streams. Biol. Invasions 2012, 14, 1881–1893. [Google Scholar] [CrossRef]
- George, B.; Brandon, C.; Erwin, M. Degradation Rates of Native versus Exotic Leaves in a Tributary of the Yellow River in Georgia. Am. J. Plant. Sci. 2017, 8, 1967–1976. [Google Scholar] [CrossRef] [Green Version]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 436–443. [Google Scholar] [CrossRef]
- Abelho, M. From litterfall to breakdown in streams: A Review. Sci. World 2001, 1, 656–680. [Google Scholar] [CrossRef] [Green Version]
- Cornut, J.; Elger, A.; Lambrigot, D.; Mamonier, P.; Chauvet, E. Early stages of leaf decomposition are mediated by aquatic fungi in the hyporheic zone of woodland streams. Freshwater Biol. 2010, 55, 2541–2556. [Google Scholar] [CrossRef]
- Hladyz, S.; Gessner, M.O.; Giller, P.S.; Pozo, J.; Woodward, G. Resource quality and stoichiometric constraints on stream ecosystem functioning. Freshwater Biol. 2009, 54, 957–970. [Google Scholar] [CrossRef]
- Bottollier-Curtet, M.; Charcosset, J.; Planty-Tabacchi, A.; Tabacchi, E. Degradation of native and exotic riparian plant leaf litter in a floodplain pond. Freshwater Biol. 2011, 56, 1798–1810. [Google Scholar] [CrossRef] [Green Version]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. From tropics to tundra: Global convergence in plant functioning. Proc. Natl Acad. Sci. USA 1997, 94, 13730–13734. [Google Scholar] [CrossRef] [Green Version]
- Ong, C.S.; Juan, J.C.; Yule, C.M. The contribution of leaching to nutrient release from leaf litter of two emergent tree species in a Malaysian tropical peat swamp forest. Hydrobiologia 2017, 794, 125–137. [Google Scholar] [CrossRef]
- Cuassolo, F.; Navarro, M.B.; Balseiro, E.; Modenutti, B. Effect of light on particulate and dissolved organic matter production of native and exotic macrophyte species in Patagonia. Hydrobiologia 2016, 766, 29–42. [Google Scholar] [CrossRef]
- MacKenzie, R.A.; Wiegner, T.N.; Kinslow, F.; Cormier, N.; Strauch, A.M. Leaf-litter inputs from an invasive nitrogen-fixing tree influence organic-matter dynamics and nitrogen inputs in a Hawaiian river. Freshwater Sci. 2013, 32, 1036–1052. [Google Scholar] [CrossRef]
- Mineau, M.M.; Baxter, C.V.; Marcarelli, A.M.; Minshall, G.W. An invasive riparian tree reduces stream ecosystem efficiency via a recalcitrant organic matter subsidy. Ecology 2012, 93, 1501–1508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allan, J.D.; Castillo, M.M. Stream Ecology. Structure and Function of Running Waters; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Predictor | F-Value | p-Value |
|---|---|---|---|
| Temperature | Treatment | 1.52 | 0.23 |
| Week | 9155.33 | <0.001 | |
| Treatment:Week | 0.70 | 0.82 | |
| pH | Treatment | 42.03 | <0.001 |
| Week | 92.66 | <0.001 | |
| Treatment:Week | 1.15 | 0.31 | |
| Conductivity | Treatment | 69.99 | <0.001 |
| Week | 170.40 | <0.001 | |
| Treatment:Week | 2.70 | <0.001 | |
| Phosphate | Treatment | 30.51 | <0.001 |
| Week | 15.22 | <0.001 | |
| Treatment:Week | 1.00 | 0.47 | |
| Nitrite | Treatment | 23.33 | <0.001 |
| Week | 13.93 | <0.001 | |
| Treatment:Week | 1.96 | 0.01 | |
| Nitrate | Treatment | 18.18 | <0.001 |
| Week | 32.94 | <0.001 | |
| Treatment:Week | 4.91 | <0.001 | |
| Silicate | Treatment | 1.82 | 0.13 |
| Week | 31.80 | <0.001 | |
| Treatment:Week | 0.67 | 0.85 | |
| Ammonium | Treatment | 7.39 | 0.001 |
| Week | 11.11 | <0.001 | |
| Treatment:Week | 2.47 | 0.002 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mutshekwa, T.; Cuthbert, R.N.; Wasserman, R.J.; Murungweni, F.M.; Dalu, T. Nutrient Release Dynamics Associated with Native and Invasive Leaf Litter Decomposition: A Mesocosm Experiment. Water 2020, 12, 2350. https://doi.org/10.3390/w12092350
Mutshekwa T, Cuthbert RN, Wasserman RJ, Murungweni FM, Dalu T. Nutrient Release Dynamics Associated with Native and Invasive Leaf Litter Decomposition: A Mesocosm Experiment. Water. 2020; 12(9):2350. https://doi.org/10.3390/w12092350
Chicago/Turabian StyleMutshekwa, Thendo, Ross N. Cuthbert, Ryan J. Wasserman, Florence M. Murungweni, and Tatenda Dalu. 2020. "Nutrient Release Dynamics Associated with Native and Invasive Leaf Litter Decomposition: A Mesocosm Experiment" Water 12, no. 9: 2350. https://doi.org/10.3390/w12092350
APA StyleMutshekwa, T., Cuthbert, R. N., Wasserman, R. J., Murungweni, F. M., & Dalu, T. (2020). Nutrient Release Dynamics Associated with Native and Invasive Leaf Litter Decomposition: A Mesocosm Experiment. Water, 12(9), 2350. https://doi.org/10.3390/w12092350