Effects of Combined High Temperature and Waterlogging Stress at Booting Stage on Root Anatomy of Rice (Oryza sativa L.)

 ,
, 
Abstract
:1. Introduction
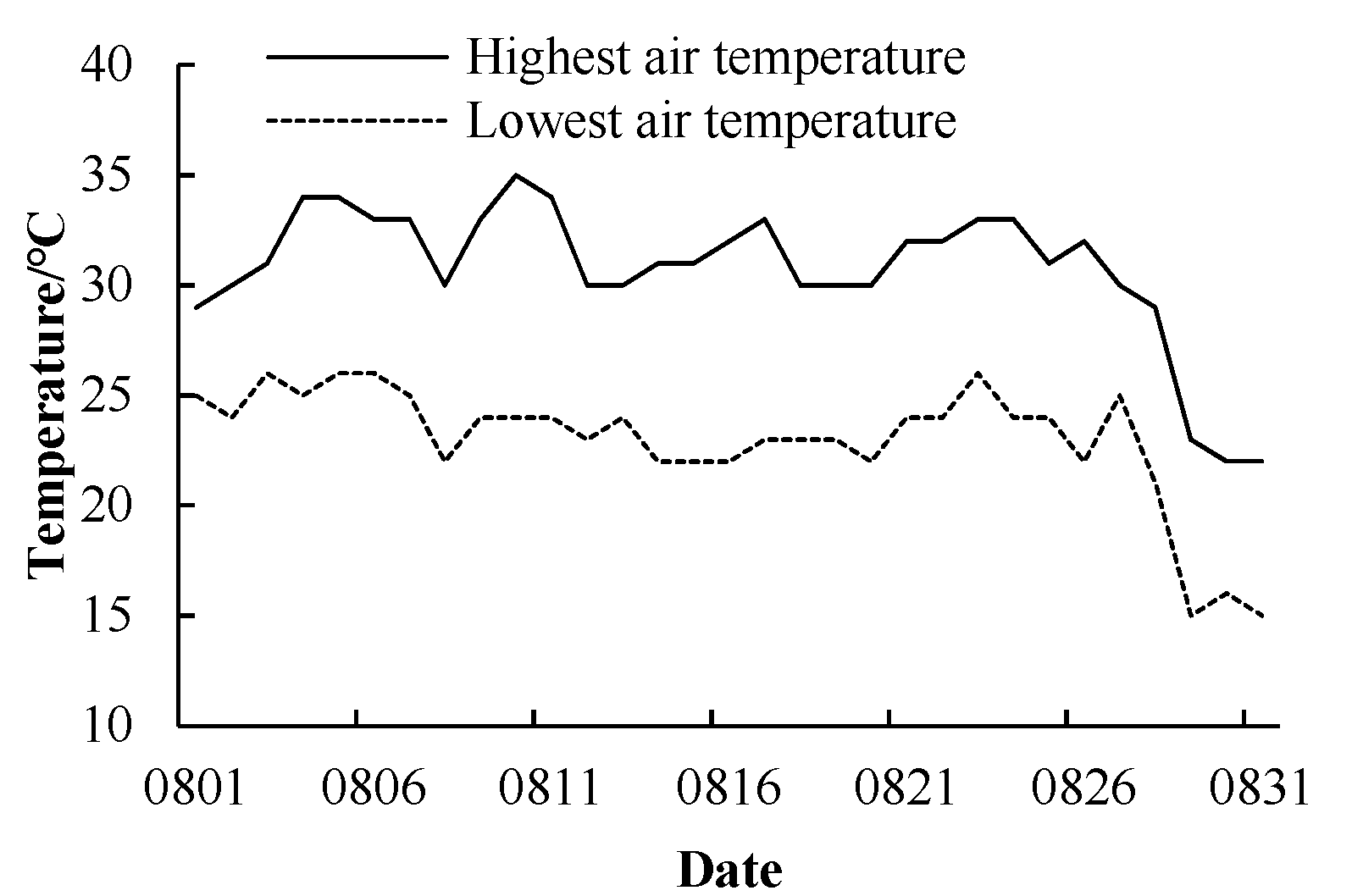
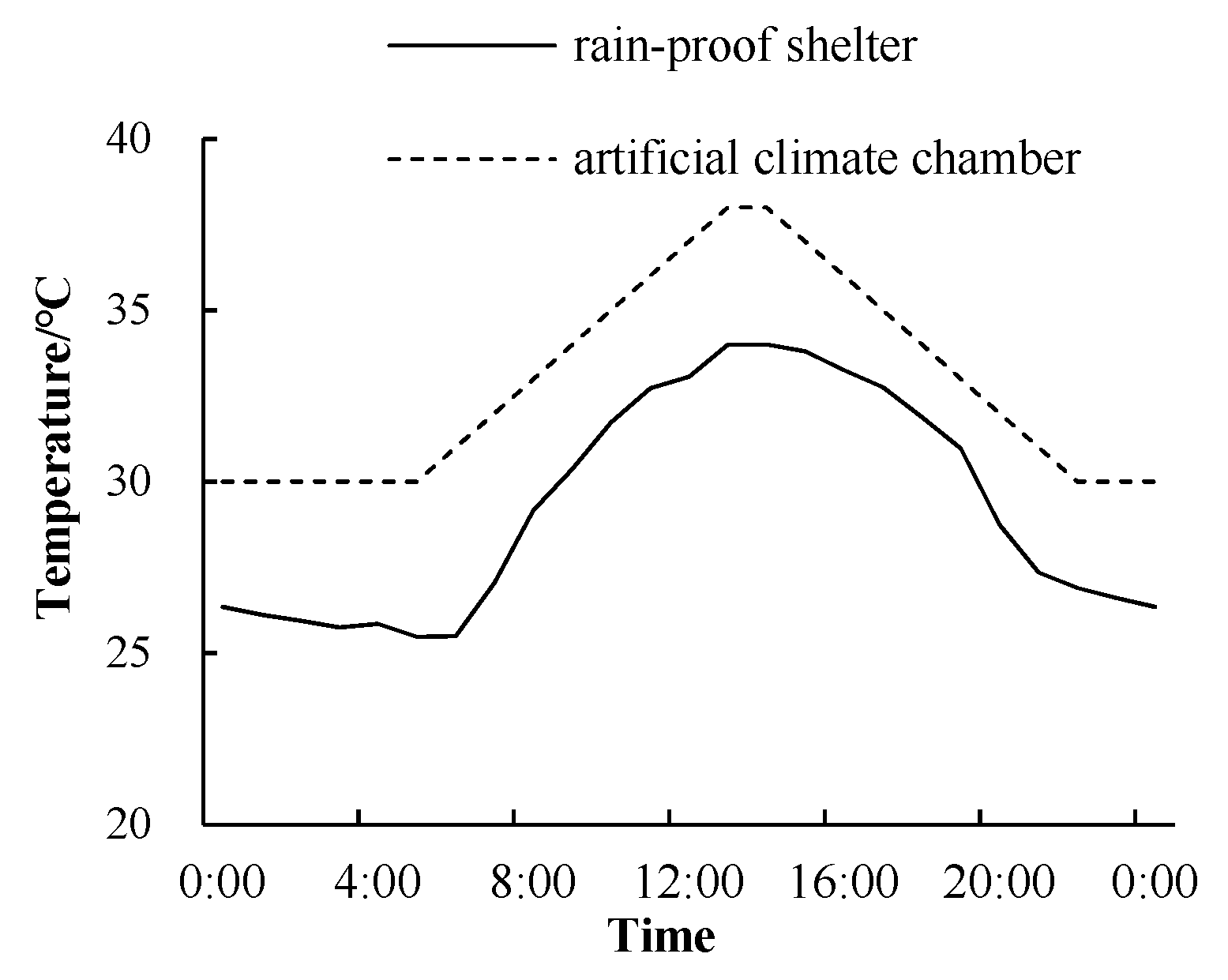
2. Materials and Methods
2.1. Plant Material and Test Site
2.2. Experimental Design
2.3. Measurement of Different Root Anatomical Parameters and Their Analysis
2.3.1. Root Anatomical Observations
2.3.2. Data Statistical Methods
2.3.3. Data Processing and Analysis
3. Results
3.1. Effects of High Temperature and Waterlogging on Anatomical Structure Parameters of Rice Roots
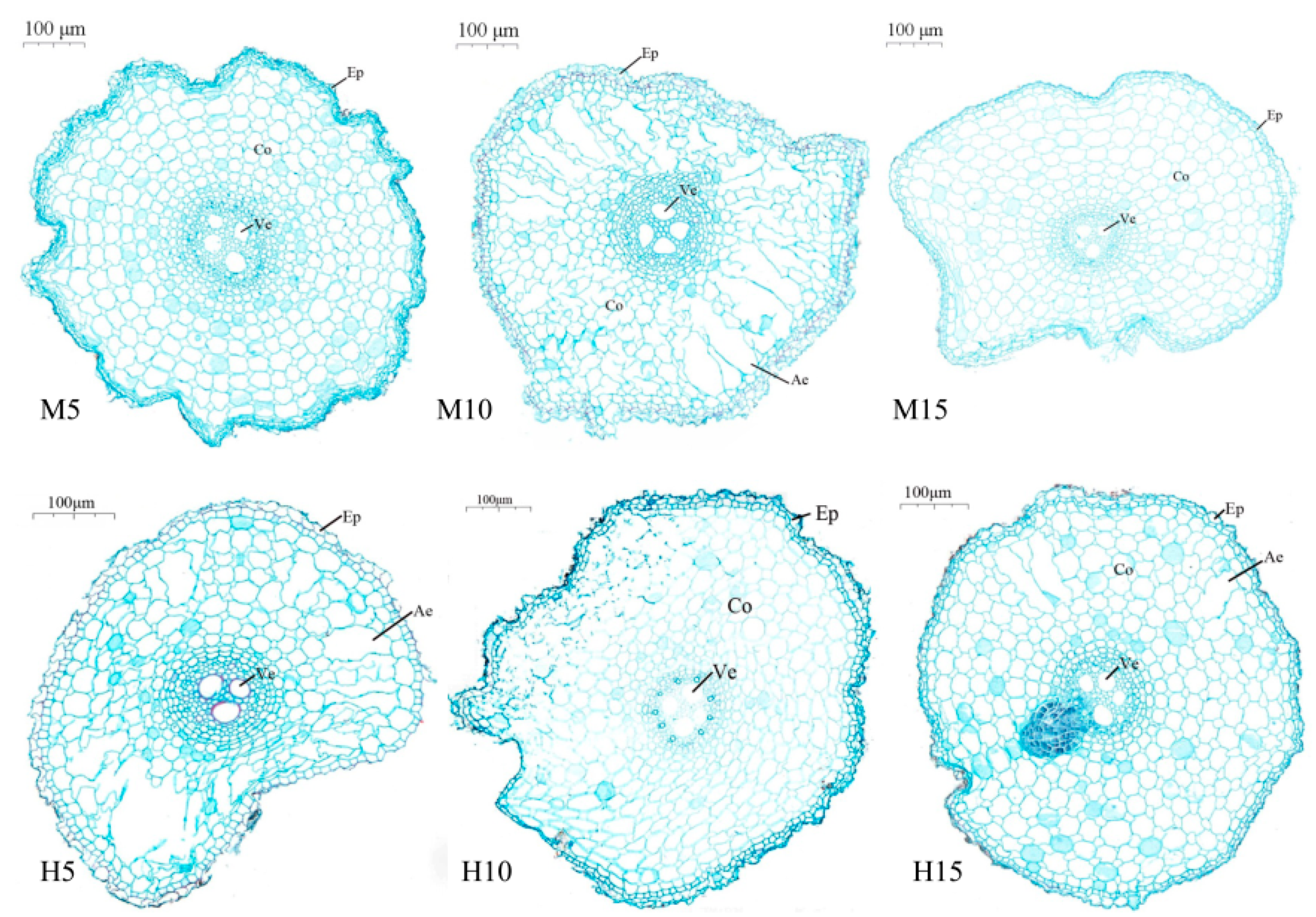
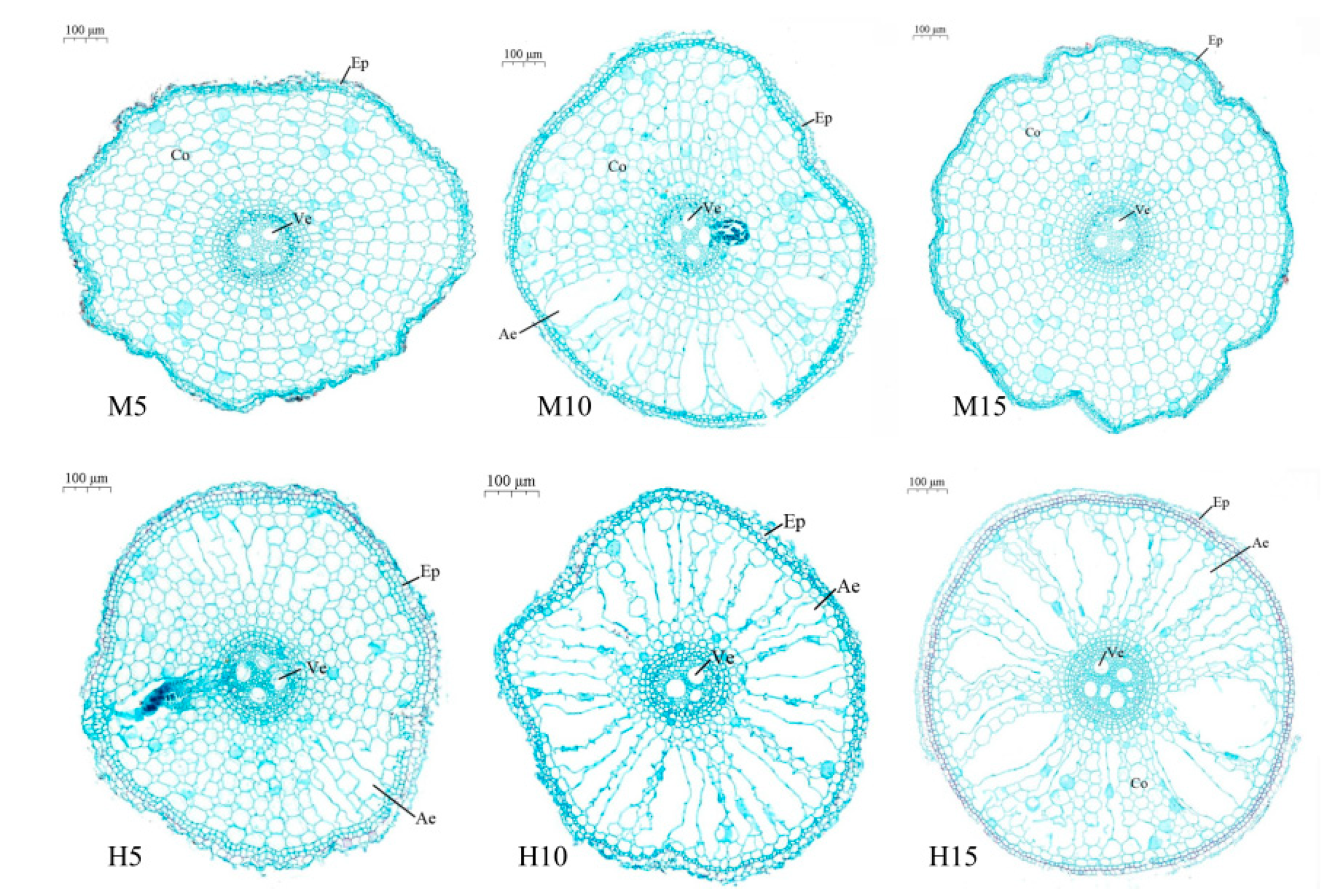
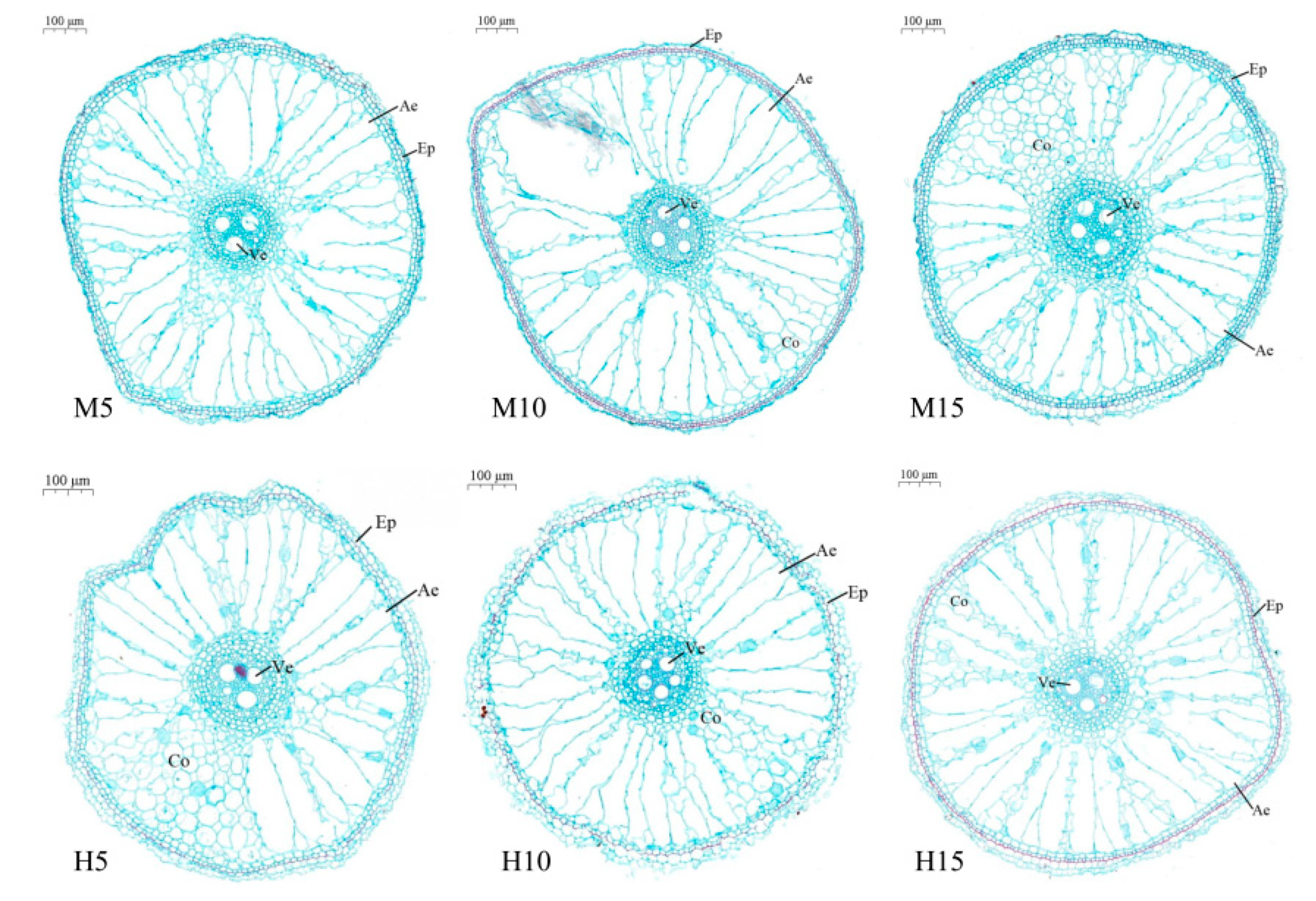
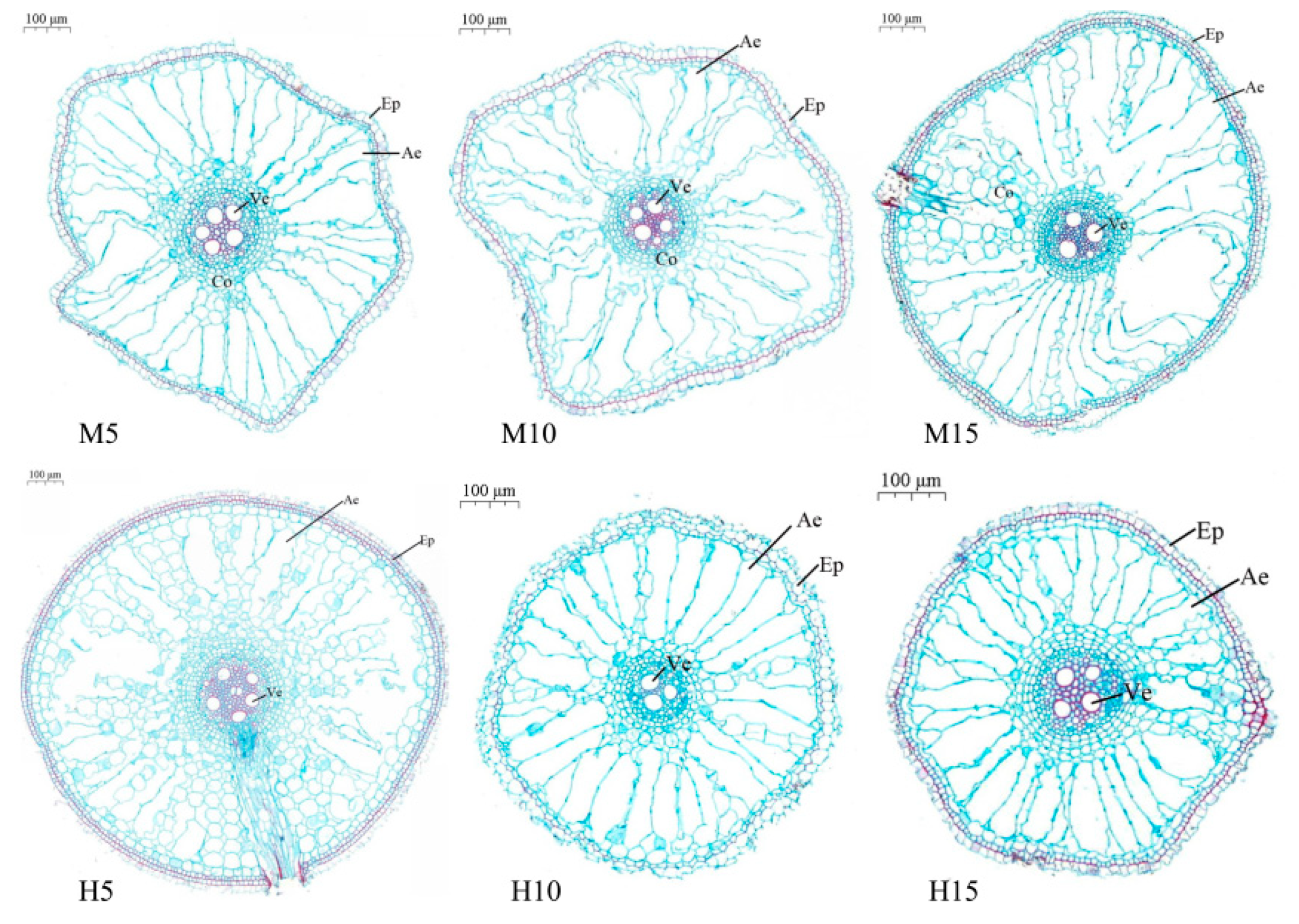
3.2. Aerenchyma Formation in Plants Exposed to High Temperature and Waterlogging Stress
3.2.1. Zero Day after the End of Stress
3.2.2. Five Days after the End of Stress
3.2.3. Ten Days after the End of Stress
3.2.4. Twenty Days after the End of Stress
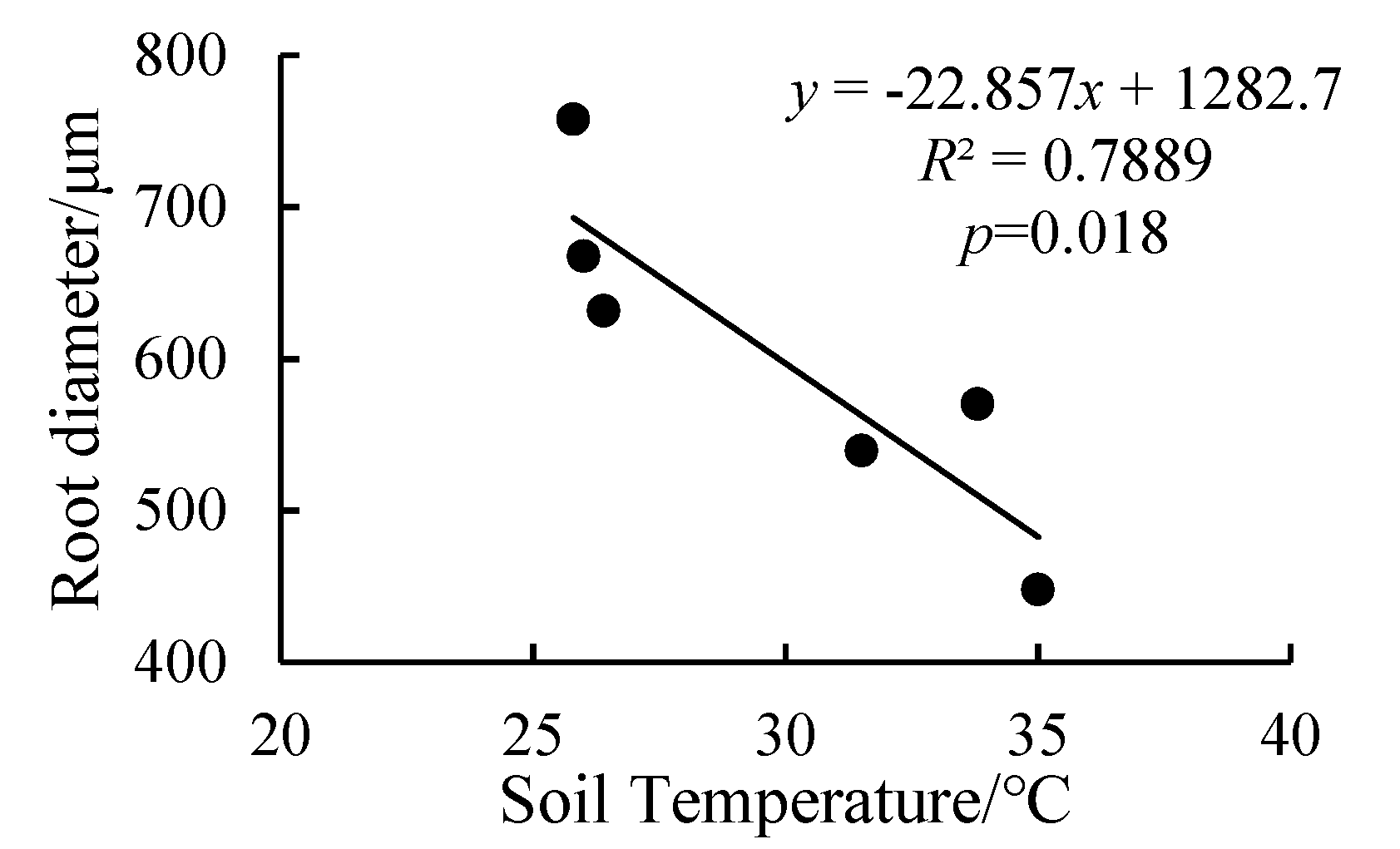
3.3. Effects of Combined High Temperature and Waterlogging Stresses on Root Diameter
3.4. Effects of Combined High Temperature and Waterlogging Stresses on Thickness of the Outer Root
3.5. Effects of Combined High Temperature and Waterlogging Stresses on Stele Diameter
3.6. Effects of High Temperature and Waterlogging on Vessel Number and Diameter
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhu, D.F.; Zhang, Y.P.; Chen, H.Z.; Xiang, J.; Zhang, Y.K. Innovation and Practice of High-Yield Rice Cultivation Technology in China. Sci. Agric. Sin. 2015, 17, 3404–3414. [Google Scholar] [CrossRef]
- IPCC (The Intergovernmental Panel on Climate Change). Climate Change 2013: The Physical Science Basis. Available online: http://www.ipcc.ch/publications_and_data/publications_and_dataShtml (accessed on 20 December 2016).
- Oreskes, N. Beyond the ivory tower: The Scientific Consensus on Climate Change. Science 2004, 5702, 1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, J.Y.; Chen, J.Q.; Piao, S.L.; Ciais, P.; Luo, Y.Q.; Wan, S.Q. Terrestrial carbon cycle affected by non-uniform climate warming. Nat. Geosci. 2014, 3, 173–180. [Google Scholar] [CrossRef]
- Butardo, V.M., Jr.; Sreenivasulu, N.; Juliano, B.O. Improving rice grain quality: State-of-the-art and future prospects. Rice Grain Qual. 2019, 1892, 19–55. [Google Scholar] [CrossRef]
- Hakata, M.; Kuroda, M.; Miyashita, T.; Yamaguchi, T.; Kojima, M.; Sakakibara, H.; Yamakawa, H. Suppression of α-amylase genes improves quality of rice grain ripened under high temperature. Plant Biotechnol. J. 2012, 9, 1110–1117. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.J.; Li, C.Y.; Lin, S.K. Influence of high temperature during grain filling on the accumulation of storage proteins and grain quality in rice (Oryza sativa L.). J. Agric. Food Chem. 2010, 19, 10545–10552. [Google Scholar] [CrossRef]
- Shi, W.J.; Li, X.; Schmidt, R.C.; Struik, P.C.; Yin, X.Y.; Jagadish, S.V.K. Pollen germination and in vivo fertilization in response to high temperature during flowering in hybrid and inbred rice. Plant Cell Environ. 2018, 6, 1287–1297. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.H.; Matsui, T.; Lin, S.H.; Lin, J.C. High temperature stress on rice anthesis: Research progress and prospects. Chin. J. Appl. Ecol. 2007, 11, 2632–2636. [Google Scholar] [CrossRef]
- Tsukaguchi, T.; Iida, Y. Effects of assimilate supply and high temperature during grain-filling period on the occurrence of various types of chalky kernels in rice plants (Oryza sativa L.). Plant Prod. Sci. 2008, 11, 203–210. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Shi, J.; Ding, Y.H.; Cui, L.L. Climatic characteristics and their changing law during summer high-temperature times in East China. Acta Geogr. Sin. 2008, 3, 237–246. [Google Scholar] [CrossRef]
- Wu, Q.X.; Su, R.R.; Liu, K.W.; Zhu, J.Q.; Yang, W.; Zhou, Y. Hot damage and concomitance of hot and waterlogging in Four-lake Basin in Jianghan Plain during the Last 50 Years. Chin. J. Agrometeorol. 2012, 4, 609–614. [Google Scholar] [CrossRef]
- Yang, S.N.; He, L.F. Analysis of atmosphere circulation and weather in August 2013. Meteorol. Mon. 2013, 11, 1521–1528. [Google Scholar] [CrossRef]
- Tran, T.; Kano-Nakata, M.; Suralta, R.; Menge, D.; Mitsuya, S.; Inukai, Y.; Yamauchi, A. Root plasticity and its functional roles were triggered by water deficit but not by the resulting changes in the forms of soil in rice. Plant Soil 2015, 386, 65–76. [Google Scholar] [CrossRef]
- Hubeau, M.; Steppe, K. Plant-PET scans: In vivo mapping of xylem and phloem functioning. Trends Plant Sci. 2015, 20, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y.; Liu, B.Y.; Wang, Y.F.; Xu, Z.W.; Zhang, X.Q. Research Progress of Plant Roots. Tianjin Agric. Sci. 2016, 11, 11–18. [Google Scholar] [CrossRef]
- Sauter, M. Root responses to flooding. Curr. Opin. Plant Biol. 2013, 16, 282–286. [Google Scholar] [CrossRef]
- Zhen, B.; Guo, X.P.; Lu, H.F. Effects of alternative stress of drought and waterlogging at tillering stage on rice root anatomical structure. Trans. Chin. Soc. Agric. Eng. Trans. 2015, 9, 107–113. [Google Scholar] [CrossRef]
- Kang, S.Z.; Zhang, J.H.; Liang, J.S. Combined effects of soil water content and temperature on plant root hydraulic conductivity. Acta Phytoecol. Sin. 1999, 3, 211–219. [Google Scholar]
- Huang, M. Responses of Rice Root Anatomy Characters to Water and Nitrogen Supply and Its Relationships with Water and Nitrogen Absorptions. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2016. [Google Scholar]
- Tian, X.H.; Deng, Y. Characterizing the rice field climatic factors under high temperature stress at anthesis. Int. Crop Sci. 2008, 4, 19–27. [Google Scholar]
- Meng, D.M. Study on the Capability of Rice Root Aerenchyma to Oxygen Transportation. Master’s Thesis, Beijing Forestry University, Beijing, China, 2008. [Google Scholar]
- Lan, X.; Gu, Z.D.; Ding, Y.F.; Wang, K.; Jiang, Q.; Zhu, C. Effect of high temperature stress on physiological characteristics of spikelet of rice during flowering stage. Chin. J. Rice Sci. 2016, 30, 637–646. [Google Scholar] [CrossRef]
- Cai, C.; Yin, X.; He, S. Responses of wheat and rice to factorial combinations of ambient and elevated CO2 and temperature in FACE experiments. Glob. Chang. Biol. 2016, 2, 856–874. [Google Scholar] [CrossRef] [PubMed]
- Yuan, A.; Wan, S.Q.; Zhou, X.H.; Afzal, A. Plant nitrogen concentration, use efficiency, and contents in a tallgrass prairie ecosystem under experimental warming. Glob. Chang. Biol. 2005, 10, 1733–1744. [Google Scholar] [CrossRef]
- Cheng, W.G.; Sakai, H.; Yagi, K.; Hasegawa, T. Combined effects of elevated [CO2] and high night temperature on carbon assimilation, nitrogen absorption, and the allocations of C and N by rice (Oryza sativa L.). Agric. For. Meteorol. 2010, 9, 1174–1181. [Google Scholar] [CrossRef]
- Evans, D.E. Aerenchyma formation. New Phytol. 2003, 1, 35–49. [Google Scholar] [CrossRef]
- Kenta, Y.; Ichirou, K. Role of ethylene signalling in the formation of constitutive aerenchyma in primary roots of rice. Aob Plants 2014, 6, plu043. [Google Scholar] [CrossRef]
- Abiko, T.; Obara, M. Enhancement of porosity and aerenchyma formation in nitrogen-deficient rice roots. Plant Sci. 2004, 215, 76–83. [Google Scholar] [CrossRef]
- Suralta, R.R.; Kano-Nakata, M.; Niones, J.M.; Inukai, Y.; Kameoka, E.; Tran, T.T.; Menge, D.; Mitsuya, S.; Yamauchi, A. Root plasticity for maintenance of productivity under abiotic stressed soil environments in rice: Progress and prospects. Field Crops Res. 2018, 220, 57–66. [Google Scholar] [CrossRef]
- Nada, R.M.; Abo-Hegazy, S.E.; Budran, E.G. The interaction of genes controlling root traits is required for the developmental acquisition of deep and thick root traits and improving root architecture in response to low water or nitrogen content in rice (Oryza sativa L.) cultivars. Plant Physiol. Biochem. 2019, 141, 122–132. [Google Scholar] [CrossRef]
- Fitter, A.; Williamson, L.; Linkohr, B.; Leyser, O. Root System Architecture Determines Fitness in an Arabidopsis Mutant in Competition for Immobile Phosphate Ions but Not for Nitrate Ions. Proc. Biol. Sci. 2002, 1504, 2017–2022. [Google Scholar] [CrossRef] [Green Version]
- Eissenstat, D.M.; Yanai, R.D. The Ecology of Root Lifespan. In Advances in Ecological Research; Academic Press: Cambridge, MA, USA, 1997; Volume 27, pp. 1–60. [Google Scholar] [CrossRef]
- Wu, Y.B.; Zhang, J.; Deng, Y.C.; Wu, J.; Wang, S.P.; Tang, Y.H.; Cui, X.Y. Effects of warming on root diameter, distribution and longevity in an alpine meadow. Plant Ecol. 2014, 215, 1057–1066. [Google Scholar] [CrossRef]
- Majdi, H.; Ohrvik, J. Interactive effects of soil warming and fertilization on root production, mortality, and longevity in a Norway spruce stand in Northern Sweden. Glob. Chang. Biol. 2004, 10, 182–188. [Google Scholar] [CrossRef]
- Niu, X.L.; Hu, T.T.; Zhang, F.C.; Duan, A.W.; Liu, Z.D.; Shen, X.J. Effect of partial water resupply at seeding stage on maize growth, water absorption capacity and anatomical structure. Sci. Agric. Sin. 2017, 16, 3110–3121. [Google Scholar] [CrossRef]
- Wang, Z.F.; Zhang, S.Q.; Liu, X.F. Root system hydraulic conductivity of different genotype maize and its relationship with root anatomy. Chin. J. Appl. Ecol. 2005, 12, 2349–2352. [Google Scholar] [CrossRef]
- Boughalleb, F.; Abdellaoui, R.; Ben-Brahim, N.; Neffati, M. Anatomical adaptations of Astragalus gombiformis Pomel. under drought stress. Central Eur. J. Biol. 2014, 12, 1215–1225. [Google Scholar] [CrossRef]
- Enstone, D.E.; Peterson, C.A.; Ma, F.S. Root Endodermis and Exodermis: Structure, Function, and Responses to the Environment. J. Plant Growth Regul. 2002, 21, 335–351. [Google Scholar] [CrossRef]
- Miyamoto, N.; Steudle, E.; Hirasawa, T.; Lafitte, R. Hydraulic conductivity of rice roots. J. Exp. Bot. 2001, 362, 1835–1846. [Google Scholar] [CrossRef]
- Qian, L.; Wang, X.; Luo, Y.; Sun, H.; Luo, W. Responses of Cotton at Different Growth Stages to Aeration Stress under the Influence of High Temperature. Crop Sci. 2018, 58, 342–353. [Google Scholar] [CrossRef]
- Zhen, B.; Zhou, X.; Lu, H.; Li, H. The Effects of Alternate Hot Wave and Waterlogging on Physiological Traits of Rice at Booting Stage. J. Irrig. Drain. 2019, 3, 1–7. [Google Scholar] [CrossRef]
- Zhen, B.; Zhou, X.G.; Lu, H.F.; Li, H.Z. Effect of interaction of high temperature at jointing stage and waterlogging on growth and development of rice. Trans. Chin. Soc. Agric. Eng. Trans. CSAE 2018, 21, 105–111. [Google Scholar] [CrossRef]
- Takahashi, N.; Kurata, K. Effects of Silica Application on Transpiration from Rice Panicle under Elevated CO2 Concentration. J. Agric. Meteorol. 2016, 6, 1133–1135. [Google Scholar] [CrossRef]
- Chen, J.Z.; Chen, M.L.; He, Y.Q. Effects of soil water condition and microclimate on transpiration rate of rice. J. Appl. Ecol. 2001, 1, 63–67. [Google Scholar] [CrossRef]
- Qi, D.L.; Wu, Q.X.; Zhu, J.Q. Nitrogen and phosphorus losses from paddy fields and the yield of rice with different water and nitrogen management practices. Sci. Rep. 2020, 10, 9734. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days after Stress Ended | Experimental Factor | Aerenchyma Area | Root Diameter | Thickness of Outer Root | Stele Diameter | Vessel Diameter |
|---|---|---|---|---|---|---|
| 0 day | T | 0.000 * | 0.000 * | 0.011 * | 0.010 * | 0.000 * |
| W | 0.000 * | 0.065 | 0.102 | 0.003 * | 0.006 * | |
| T × W | 0.000 * | 0.378 | 0.238 | 0.333 | 0.001 * | |
| 5 days | T | 0.000 * | 0.165 | 0.096 | 0.559 | 0.016 * |
| W | 0.000 * | 0.156 | 0.000 * | 0.032 * | 0.000 * | |
| T × W | 0.000 * | 0.052 | 0.000 * | 0.083 | 0.001 * | |
| 10 days | T | 0.042 | 0.255 | 0.237 | 0.141 | 0.043 * |
| W | 0.000 * | 0.128 | 0.007 * | 0.085 | 0.744 | |
| T × W | 0.000 * | 0.026 * | 0.000 * | 0.024 * | 0.000 * | |
| 20 days | T | 0.002 * | 0.001 * | 0.000 * | 0.965 | 0.000 * |
| W | 0.003 * | 0.047 * | 0.000 * | 0.000 * | 0.012 * | |
| T × W | 0.002 * | 0.427 | 0.000 * | 0.000 * | 0.000 * |
| Treatment | 0 Day after the End of Stress | 5 Days after the End of Stress | 10 Days after the End of Stress | 20 Days after the End of Stress |
|---|---|---|---|---|
| M5 | 0.00d | 0.00d | 318,980.13b | 262,343.33a |
| M10 | 62,725.87a | 76,833.10c | 442,274.67a | 242,150.33a |
| M15 | 0.00d | 0.00d | 223,713.24b | 308,096.25a |
| H5 | 31,830.90b | 35,345.34d | 306,063.57b | 300,848.94a |
| H10 | 0.00d | 191,419.50b | 274,994.44b | 147,754.71b |
| H15 | 12,859.79c | 317,942.06a | 315,199.43b | 172,162.13b |
| Treatment | 0 Day after the End of Stress | 5 Days after the End of Stress | 10 Days after the End of Stress | 20 Days after the End of Stress |
|---|---|---|---|---|
| M5 | 631.83ab | 864.60abc | 888.82b | 788.77ab |
| M10 | 667.91ab | 932.11ab | 993.03a | 766.98bc |
| M15 | 758.19a | 861.05abc | 857.13b | 872.25a |
| H5 | 448c | 732.30c | 834.56b | 675.14c |
| H10 | 570.42bc | 744.79bc | 872.84b | 701.25bc |
| H15 | 539.70bc | 972.12a | 937.45ab | 731.28bc |
| Treatment | 0 Day after the End of Stress | 5 Days after the End of Stress | 10 Days after the End of Stress | 20 Days after the End of Stress |
|---|---|---|---|---|
| M5 | 43.2ab | 42.64c | 50.6b | 47.48b |
| M10 | 47.58a | 55.3b | 47.42bc | 45.12bc |
| M15 | 40.44b | 49.34b | 47.54bc | 56.84a |
| H5 | 41.00b | 35.06d | 41.30c | 44.06c |
| H10 | 39.26b | 39.3cd | 51.08b | 41.78c |
| H15 | 37.82b | 63.52a | 59.64a | 42.32c |
| Treatment | 0 Day after the End of Stress | 5 Days after the End of Stress | 10 Days after the End of Stress | 20 Days after the End of Stress |
|---|---|---|---|---|
| M5 | 129.98b | 149.03b | 151.43b | 151.73bc |
| M10 | 155.04a | 163.43ab | 155.20b | 144.29bcd |
| M15 | 138.95ab | 157.46b | 174.27a | 167.89b |
| H5 | 110.12c | 147.11b | 156.59b | 205.29a |
| H10 | 135.26b | 153.37b | 154.55b | 121.26d |
| H15 | 134.84b | 179.23a | 153.06b | 138.16cd |
| Treatment | 0 Day after the End of Stress | 5 Days after the End of Stress | 10 Days after the End of Stress | 20 Days after the End of Stress | ||||
|---|---|---|---|---|---|---|---|---|
| VN | VD/μm | VN | VD/μm | VN | VD/μm | VN | VD/μm | |
| M5 | 3.67a | 29.50ab | 4.00ab | 34.44cd | 3.00b | 37.06a | 4.33a | 35.31a |
| M10 | 3.67a | 31.02a | 4.00ab | 37.39b | 4.33a | 36.67a | 5.00a | 30.70bc |
| M15 | 3.67a | 31.37a | 3.33b | 33.48de | 5.00a | 34.00c | 4.00a | 35.09a |
| H5 | 3.33a | 28.48b | 4.00ab | 35.18c | 4.33a | 33.53c | 4.67a | 32.17b |
| H10 | 4.00a | 30.21ab | 3.33b | 40.42a | 4.00a | 34.57bc | 3.00b | 31.85b |
| H15 | 3.67a | 25.34c | 5.00a | 32.65e | 4.00a | 36.25ab | 4.00a | 29.21c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhen, B.; Li, H.; Niu, Q.; Qiu, H.; Tian, G.; Lu, H.; Zhou, X. Effects of Combined High Temperature and Waterlogging Stress at Booting Stage on Root Anatomy of Rice (Oryza sativa L.). Water 2020, 12, 2524. https://doi.org/10.3390/w12092524
Zhen B, Li H, Niu Q, Qiu H, Tian G, Lu H, Zhou X. Effects of Combined High Temperature and Waterlogging Stress at Booting Stage on Root Anatomy of Rice (Oryza sativa L.). Water. 2020; 12(9):2524. https://doi.org/10.3390/w12092524
Chicago/Turabian StyleZhen, Bo, Huizhen Li, Qinglin Niu, Husen Qiu, Guangli Tian, Hongfei Lu, and Xinguo Zhou. 2020. "Effects of Combined High Temperature and Waterlogging Stress at Booting Stage on Root Anatomy of Rice (Oryza sativa L.)" Water 12, no. 9: 2524. https://doi.org/10.3390/w12092524
APA StyleZhen, B., Li, H., Niu, Q., Qiu, H., Tian, G., Lu, H., & Zhou, X. (2020). Effects of Combined High Temperature and Waterlogging Stress at Booting Stage on Root Anatomy of Rice (Oryza sativa L.). Water, 12(9), 2524. https://doi.org/10.3390/w12092524