
Effects of Plastic Debris on the Biofilm Bacterial Communities in Lake Water



Abstract
:1. Introduction
2. Materials and Methods
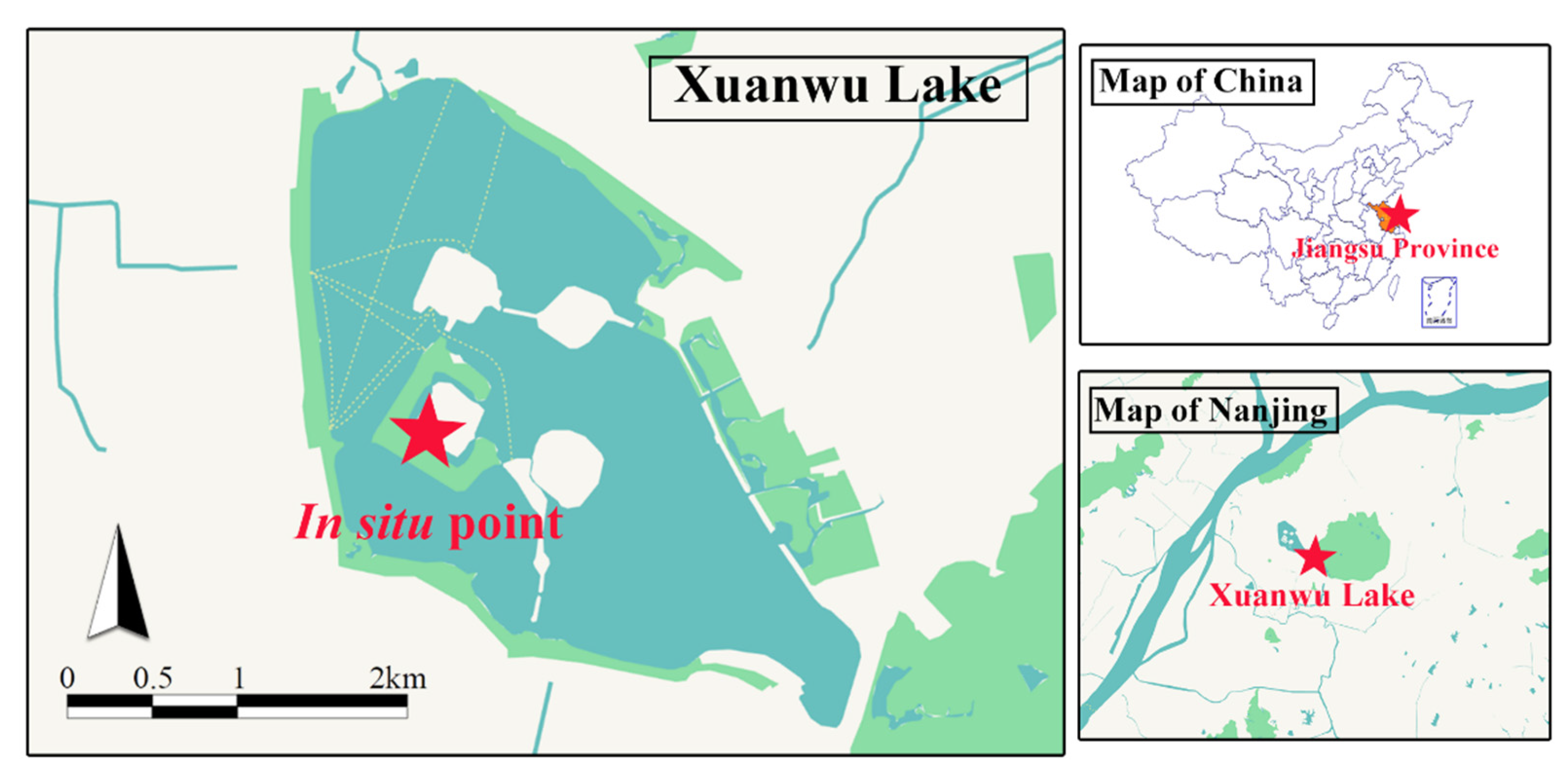
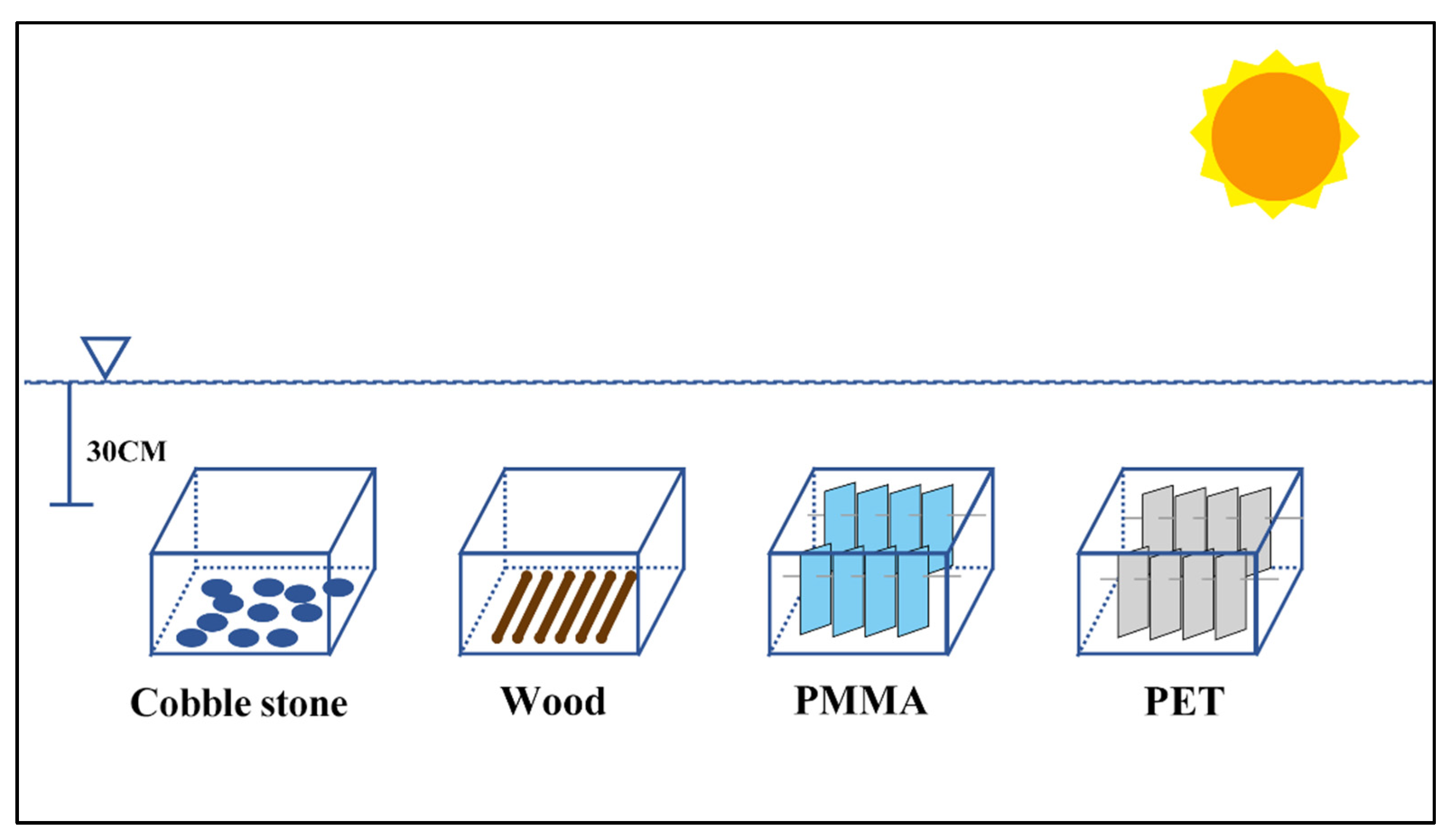
2.1. Experimental Design and Setup
2.2. Analyses of Water Chemistry
2.3. DNA Extraction, Amplification and Sequencing
2.4. Sequencing Processing and Data Analysis
2.5. Statistical Analysis
3. Results
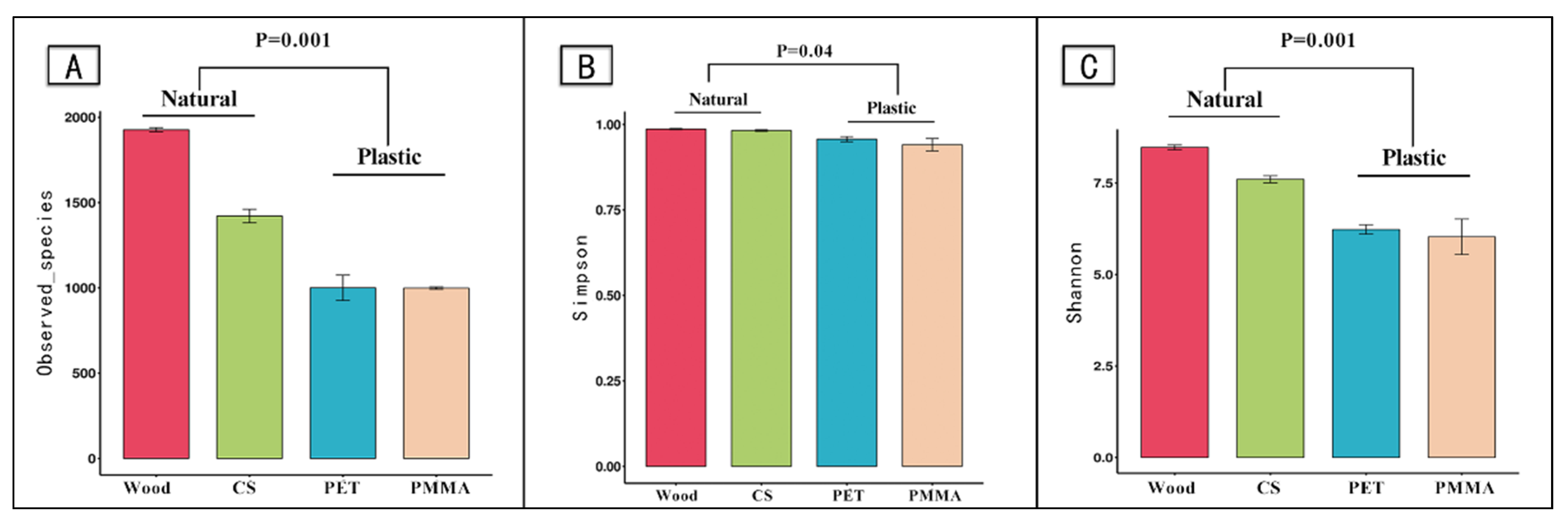
3.1. Taxonomic Annotation and Alpha Diversity
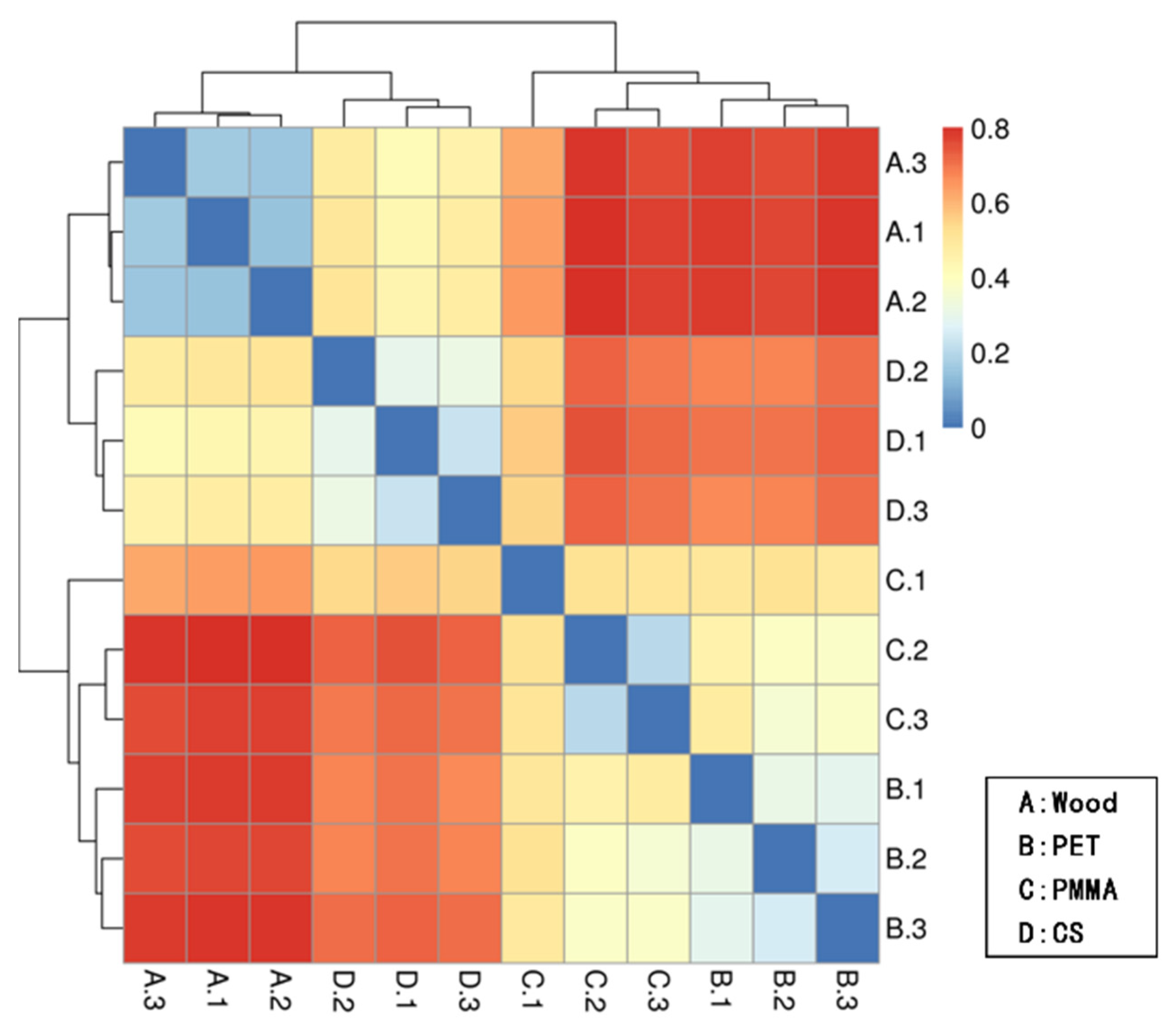
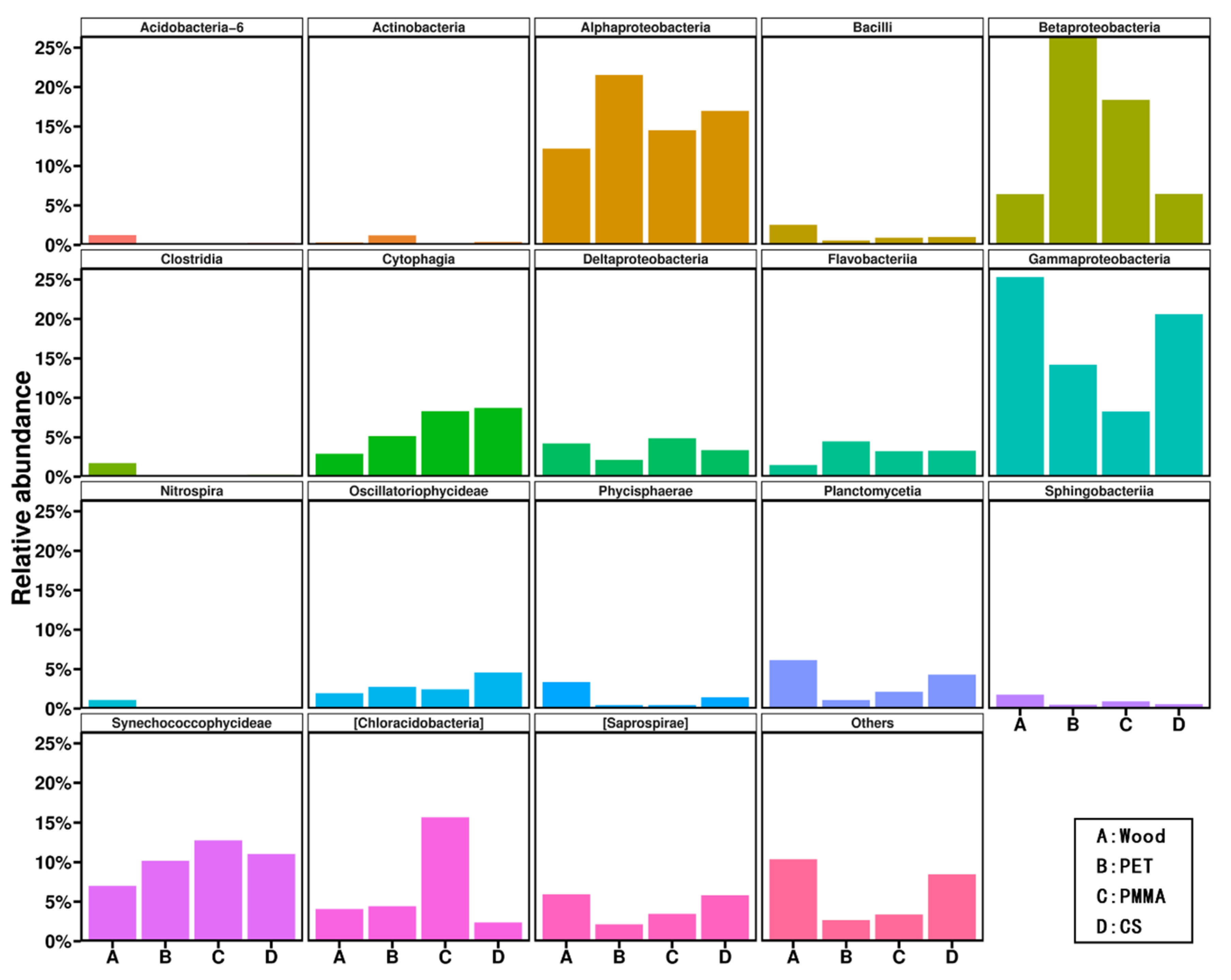
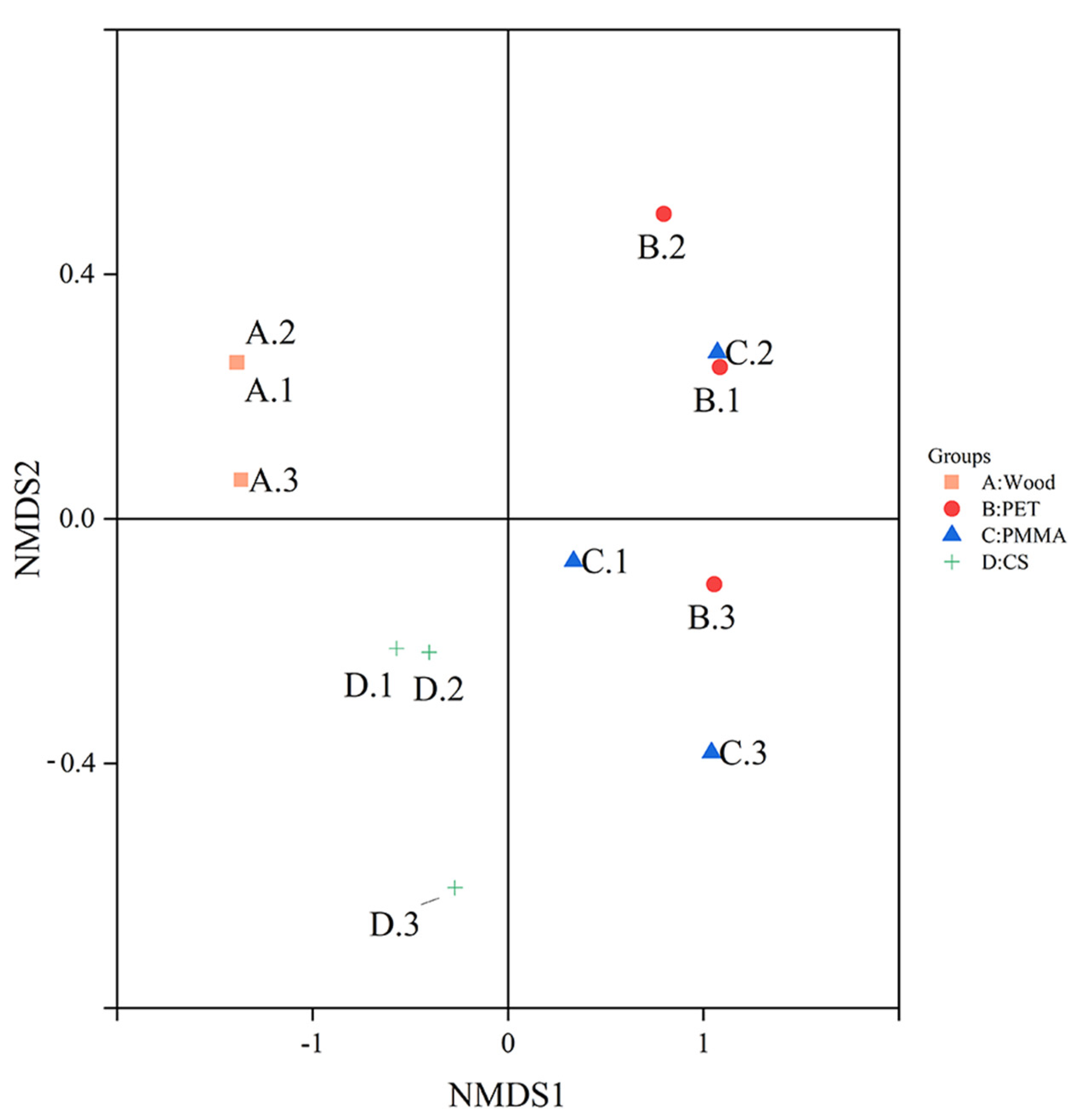
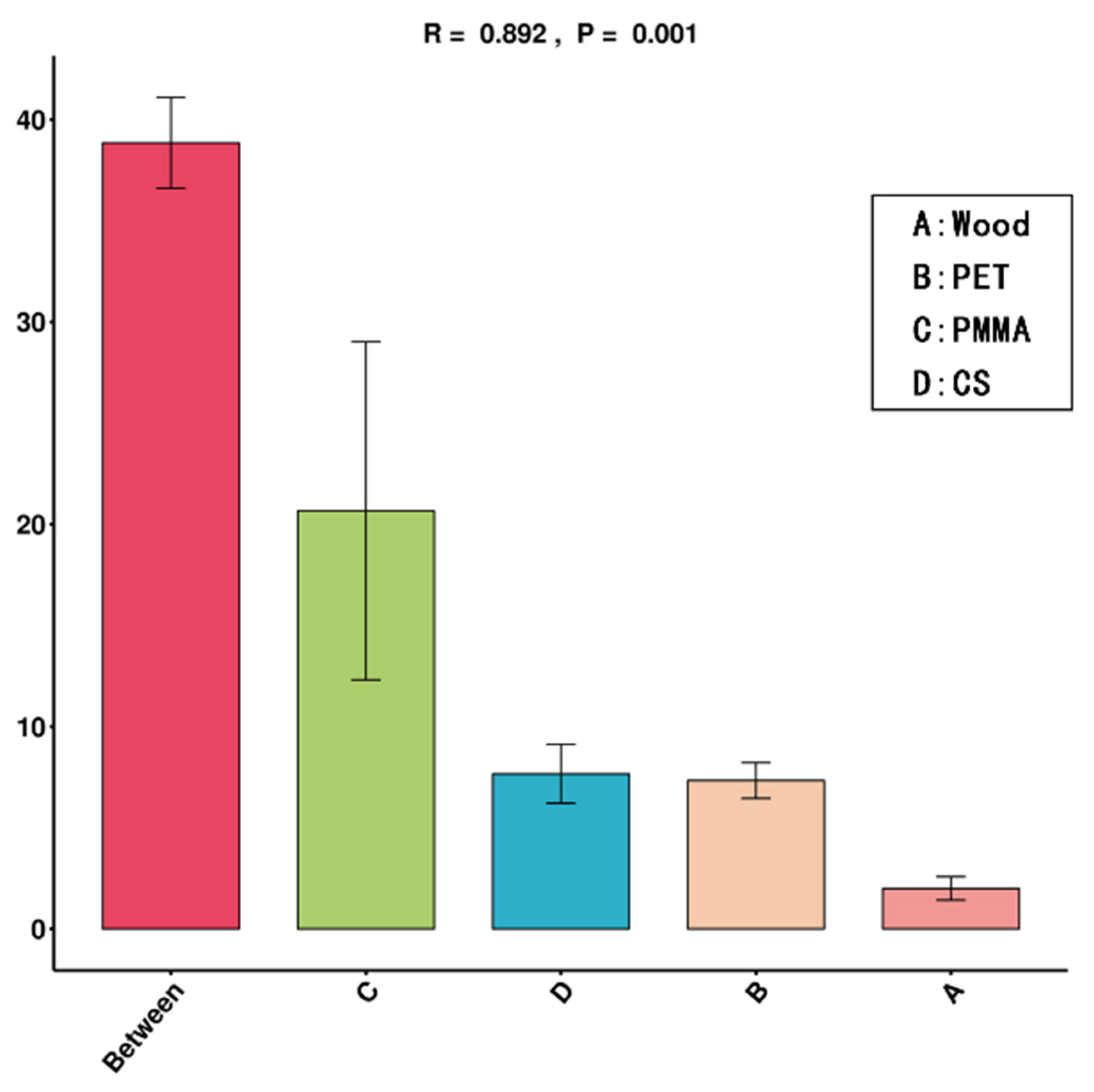
3.2. Community Composition and Structure of Biofilm
4. Discussion
4.1. Biofilm Community Diversity on Microplastics
4.2. Practical Implications of This Study
4.3. Limitation and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Plastics Production. Plastics-the Facts 2019; European plastics production: Brussels, Belgium, 2019. [Google Scholar]
- Thompson, R.C.; Olsen, Y.; Mitchell, R.P.; Davis, A.; Rowland, S.J.; John, A.W.G.; McGonigle, D.; Russell, A.E. Lost at sea: Where is all the plastic? Science 2004, 304, 838. [Google Scholar] [CrossRef] [PubMed]
- Waldschläger, K.; Lechthaler, S.; Stauch, G.; Schüttrumpf, H. The way of microplastic through the environment—Application of the source-pathway-receptor model (review). Sci. Total Environ. 2020, 713, 136584. [Google Scholar] [CrossRef]
- Wong, J.K.H.; Lee, K.K.; Tang, K.H.D.; Yap, P.-S. Microplastics in the freshwater and terrestrial environments: Prevalence, fates, impacts and sustainable solutions. Sci. Total Environ. 2020, 719, 137512. [Google Scholar] [CrossRef] [PubMed]
- Hirai, H.; Takada, H.; Ogata, Y.; Yamashita, R.; Mizukawa, K.; Saha, M.; Kwan, C.; Moore, C.; Gray, H.; Laursen, D.; et al. Organic micropollutants in marine plastics debris from the open ocean and remote and urban beaches. Mar. Pollut. Bull. 2011, 62, 1683–1692. [Google Scholar] [CrossRef]
- Ding, J.; Huang, Y.; Liu, S.; Zhang, S.; Zou, H.; Wang, Z.; Zhu, W.; Geng, J. Toxicological Effects of Nano- and Micro-Polystyrene Plastics On Red Tilapia: Are Larger Plastic Particles More Harmless? J. Hazard. Mater. 2020, 396, 122693. [Google Scholar] [CrossRef]
- Arias-Andres, M.; Klümper, U.; Rojas-Jimenez, K.; Grossart, H.-P. Microplastic pollution increases gene exchange in aquatic ecosystems. Environ. Pollut. 2018, 237, 253–261. [Google Scholar] [CrossRef] [Green Version]
- Miao, L.; Wang, P.; Hou, J.; Yao, Y.; Liu, Z.; Liu, S.; Li, T. Distinct community structure and microbial functions of biofilms colonizing microplastics. Sci. Total Environ. 2019, 650, 2395–2402. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Rajeev, M.; Sushmitha, T.J.; Toleti, S.R.; Pandian, S.K. Culture dependent and independent analysis and appraisal of early stage biofilm-forming bacterial community composition in the southern coastal seawater of india. Sci. Total Environ. 2019, 666, 308–320. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.-C. Biofouling in water systems—Cases, causes and countermeasures. Appl. Microbiol. Biotechnol. 2002, 59, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Flower, R.J. Effects of light and substrate on the benthic diatoms in an oligotrophic lake: A comparison between natural and artificial substrates. J. Phycol. 2012, 48, 1166–1177. [Google Scholar] [CrossRef]
- Mai, Y.Z.; Peng, S.Y.; Lai, Z.N. Structural and functional diversity of biofilm bacterial communities along the pearl river estuary, south china. Reg. Stud. Mar. Sci. 2020, 33, 100926. [Google Scholar] [CrossRef]
- Feng, L.; He, L.; Jiang, S.; Chen, J.; Zhou, C.; Qian, Z.-J.; Hong, P.; Sun, S.; Li, C. Investigating the composition and distribution of microplastics surface biofilms in coral areas. Chemosphere 2020, 252, 126565. [Google Scholar] [CrossRef] [PubMed]
- Misic, C.; Harriague, A.C. Development of marine biofilm on plastic: Ecological features in different seasons, temperatures, and light regimes. Hydrobiologia 2019, 835, 129–145. [Google Scholar] [CrossRef]
- Calapez, A.R.; Elias, C.L.; Alves, A.; Almeida, S.F.P.; Brito, A.G.; Feio, M.J. Shifts in biofilms’ composition induced by flow stagnation, sewage contamination and grazing. Ecol. Indic. 2020, 111, 106006. [Google Scholar] [CrossRef]
- Wang, C.Q.; Dong, D.M.; Zhang, L.W.; Song, Z.W.; Hua, X.Y.; Guo, Z.Y. Response of freshwater biofilms to antibiotic florfenicol and ofloxacin stress: Role of extracellular polymeric substances. Int. J. Environ. Res. Public Health 2019, 16, 715. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.T.; Hua, X.Y.; Zhang, L.W.; Song, N.; Dong, D.M.; Guo, Z.Y. Influence of organic carbon fractions of freshwater biofilms on the sorption for phenanthrene and ofloxacin: The important role of aliphatic carbons. Sci. Total Environ. 2019, 685, 818–826. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, F.; Wu, W.; Wan, J.; Yang, J.; Xiang, S.; Wu, Y. Biosorption of high-concentration cu (ii) by periphytic biofilms and the development of a fiber periphyton bioreactor (fpbr). Bioresour. Technol. 2018, 248, 127–134. [Google Scholar] [CrossRef]
- Shabbir, S.; Faheem, M.; Ali, N.; Kerr, P.G.; Wang, L.-F.; Kuppusamy, S.; Li, Y. Periphytic biofilm: An innovative approach for biodegradation of microplastics. Sci. Total Environ. 2020, 717, 137064. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, X.; Li, Y.; Li, J.; Wang, F.; Xia, S.; Zhao, J. Biofilm alters tetracycline and copper adsorption behaviors onto polyethylene microplastics. Chem. Eng. J. 2020, 392, 123808. [Google Scholar] [CrossRef]
- Chen, X.; Xiong, X.; Jiang, X.; Shi, H.; Wu, C. Sinking of floating plastic debris caused by biofilm development in a freshwater lake. Chemosphere 2019, 222, 856–864. [Google Scholar] [CrossRef]
- Tu, C.; Chen, T.; Zhou, Q.; Liu, Y.; Wei, J.; Waniek, J.J.; Luo, Y. Biofilm formation and its influences on the properties of microplastics as affected by exposure time and depth in the seawater. Sci. Total Environ. 2020, 734, 139237. [Google Scholar] [CrossRef] [PubMed]
- Zettler, E.R.; Mincer, T.J.; Amaral-Zettler, L.A. Life in the “plastisphere”: Microbial communities on plastic marine debris. Environ. Sci. Technol. 2013, 47, 7137–7146. [Google Scholar] [CrossRef] [PubMed]
- McCormick, A.R.; Hoellein, T.J.; London, M.G.; Hittie, J.; Scott, J.W.; Kelly, J.J. Microplastic in surface waters of urban rivers: Concentration, sources, and associated bacterial assemblages. ECOSPHERE 2016, 7, e01556. [Google Scholar] [CrossRef]
- Debroas, D.; Mone, A.; Ter Halle, A. Plastics in the North Atlantic garbage patch: A boat-microbe for hitchhikers and plastic degraders. Sci. Total Environ. 2017, 599–600, 1222. [Google Scholar] [CrossRef] [PubMed]
- Ogonowski, M.; Motiei, A.; Ininbergs, K.; Hell, E.; Gerdes, Z.; Udekwu, K.I.; Bacsik, Z.; Gorokhova, E. Evidence for selective bacterial community structuring on microplastics. Environ. Microbiol. 2018, 20, 2796–2808. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Pan, J.; Li, M.; Li, Y.; Bartlam, M.; Wang, Y. Selective enrichment of bacterial pathogens by microplastic biofilm. Water Res. 2019, 165, 114979. [Google Scholar] [CrossRef] [PubMed]
- Amaral-Zettler, L.A.; Zettler, E.R.; Mincer, T.J. Ecology of the plastisphere. Nat. Rev. Microbiol. 2020, 18, 139–151. [Google Scholar] [CrossRef]
- Miao, L.; Yu, Y.; Adyel, T.M.; Wang, C.; Liu, Z.; Liu, S.; Huang, L.; You, G.; Meng, M.; Qu, H.; et al. Distinct microbial metabolic activities of biofilms colonizing microplastics in three freshwater ecosystems. J. Hazard. Mater. 2021, 403, 123577. [Google Scholar] [CrossRef]
- Kettner, M.T.; Rojas-Jimenez, K.; Oberbeckmann, S.; Labrenz, M.; Grossart, H.P. Microplastics alter composition of fungal communities in aquatic ecosystems. Environ. Microbiol. 2017, 19, 4447–4459. [Google Scholar] [CrossRef]
- Miao, L.; Wang, C.; Adyel, T.M.; Wu, J.; Liu, Z.; You, G.; Meng, M.; Qu, H.; Huang, L.; Yu, Y.; et al. Microbial carbon metabolic functions of biofilms on plastic debris influenced by the substrate types and environmental factors. Environ. Int. 2020, 143, 106007. [Google Scholar] [CrossRef]
- Gong, M.; Yang, G.; Zhuang, L.; Zeng, E.Y. Microbial biofilm formation and community structure on low-density polyethylene microparticles in lake water microcosms. Environ. Pollut. 2019, 252, 94–102. [Google Scholar] [CrossRef]
- Yang, H.; Flower, R.J.; Thompson, J.R. Sustaining china’s water resources. Science 2013, 339, 141. [Google Scholar] [CrossRef]
- Dantas, L.C.D.M.; Silva-Neto, J.P.D.; Dantas, T.S.; Naves, L.Z.; das Neves, F.D.; da Mota, A.S. Bacterial Adhesion and Surface Roughness for Different Clinical Techniques for Acrylic Polymethyl Methacrylate. Int. J. Dent. 2016, 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Zhang, B.; Liu, Y.; Suo, X.; Li, H. Influence of surface topography on bacterial adhesion: A review (Review). Biointerphases 2018, 13, 060801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Gong, M.; Mai, L.; Zhuang, L.; Zeng, E.Y. Diversity and Structure of Microbial Biofilms on Microplastics in Riverine Waters of the Pearl River Delta, China. Chemosphere 2021, 272, 129870. [Google Scholar] [CrossRef] [PubMed]
- Levantesi, C.; Beimfohr, C.; Geurkink, B.; Rossetti, S.; Thelen, K.; Krooneman, J.; Snaidr, J.; van der Waarde, J.; Tandoi, V. Filamentous alphaproteobacteria associated with bulking in industrial wastewater treatment plants. Syst. Appl. Microbiol. 2004, 27, 716–727. [Google Scholar] [CrossRef]
- Oberbeckmann, S.; Kreikemeyer, B.; Labrenz, M. Environmental factors support the formation of specific bacterial assemblages on microplastics. Front. Microbiol. 2018, 8, 2709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.J.; Wang, G.Q.; Yu, H.L.; Wang, J.; Wang, S.W.; Jia, Y.; Yu, Y.Y.; Xu, J.G. First report of human infection by rhodoplanes sp., alphaproteobacteria in china. Asian Pac. J. Trop. Med. 2011, 4, 248–250. [Google Scholar] [CrossRef] [Green Version]
- Hayder, G.A.S.I.M.; Mohd Sidek, L.; Aishah Mohiyaden, H.; Basri, H.I.D.A.Y.A.H.; Fauzan Mohd Sabri, A.; Noh, M.; Nasir, M. Evaluation of Different Biomedia Performance for River Purification: Preliminary Stage. Appl. Mech. Mater. 2015, 773–774, 1365–1369. [Google Scholar] [CrossRef] [Green Version]
- Hayder, G.; Sidek, L.M.; Mohiyaden, H.A.; Basri, H.; Ming Fai, C. Comparison of various types of biomedia in river water treatment using attached growth activated sludge process. Int. J. River Basin Manag. 2016, 14, 177–182. [Google Scholar] [CrossRef]
- Yang, H.; Ma, M.G.; Thompson, J.R.; Flower, R.J. Waste management, informal recycling, environmental pollution and public health. J. Epidemiol. Community Health 2018, 72, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Tong, H.; Jiang, Q.; Hu, X.; Zhong, X. Occurrence and identification of microplastics in tap water from china. Chemosphere 2020, 252, 126493. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Shi, H.; Peng, J.; Wang, Y.; Xiong, X.; Wu, C.; Lam, P.K.S. Microplastic pollution in china’s inland water systems: A review of findings, methods, characteristics, effects, and management. Sci. Total Environ. 2018, 630, 1641–1653. [Google Scholar] [CrossRef]
- Yang, H. China must continue the momentum of green law. Nature 2014, 509, 535. [Google Scholar] [CrossRef] [Green Version]
- Hahladakis, J.N.; Velis, C.A.; Weber, R.; Iacovidou, E.; Purnell, P. An overview of chemical additives present in plastics: Migration, release, fate and environmental impact during their use, disposal and recycling. J. Hazard. Mater. 2018, 344, 179–199. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vs_Group | Df | SumsofSqs | MeanSqs | F.Model | R2 | Pr (>F) |
|---|---|---|---|---|---|---|
| A_B_C_D | 3 | 1.584971838 | 0.528323946 | 11.18835851 | 0.807530724 | 0.001 |
| A_B | 1 | 0.849885112 | 0.849885112 | 32.26187121 | 0.889691296 | 0.1 |
| A_C | 1 | 0.708050946 | 0.708050946 | 13.15419172 | 0.766820841 | 0.1 |
| A_D | 1 | 0.275281725 | 0.275281725 | 10.38952391 | 0.72201999 | 0.1 |
| B_C | 1 | 0.15905402 | 0.15905402 | 2.340901059 | 0.369174828 | 0.1 |
| B_D | 1 | 0.64271035 | 0.64271035 | 15.82457228 | 0.798230199 | 0.1 |
| C_D | 1 | 0.534961524 | 0.534961524 | 7.855713948 | 0.662609943 | 0.1 |
| Plastics and Natural Substrates | Dominant Species | Biodiversity | Environment (Field or Laboratory) | References |
|---|---|---|---|---|
| PP, PE, cobblestone and wood | Proteobacteria was the dominant followed by Bacteroidetes in MPs (at phylum level) | The order of total species richness was the same as Shannon index, which was Wood > Cobblestone > PP > PE | Laboratory environment for 21 days | [7] |
| Nine kinds of plastic materials including PP, PC, ABS, LDPE and others | α-Proteobacteria, γ-Proteobacteria, Bacteroidia and Acidimicrobiia (at class level) | The minimum Simpson index is 0.9955, and all the Shannon curves tend to flatten with the increase of the number of sequencing lines | Natural environment for 2 weeks. (Artificial coral culture areas and wild coral areas) | [14] |
| PVC, rocks and leaves | Proteobacteria in MPs and rock biofilm, and Bacteroidetes in leaf biofilm (at phylum level) | The order of observed biomass was leaf biofilm > microplastic biofilm > rock biofilm, while the order of Shannon index was microplastic biofilm > rock biofilm > leaf biofilm. | Laboratory for 2 weeks (the water was collected in the Haihe River) | [28] |
| Selected two kinds of PMD (plastic marine debris), PE and PP, compared with sargassum and seawater | The major ciliates were the genus Ephelota (Gammaproteobacteria), and the alphaproteobacterial family Hyphomonadaceae were unique to PMD | The order of mean Simpson evenness is plastics (0.95) > sargassum (0.90) > seawater (0.85) | Natural environment (collected from the North Atlantic) | [24] |
| PS, PP, PE, glass beads, native fibrous cellulose | Proteobacteria was higher on the plastics than on glass and cellulose (at the genus level) | All the Shannon evenness and Shannon–Wiener diversity were similarly low except glass substrate | Natural environment for 2 weeks (around the Baltic Sea) | [27] |
| PET, PMMA, cobble stones and wood | Alphaproteobacterial, Betaproteobacteria and Synechococcophycideae played the dominating roles on the MPs substrate | The Shannon–Wiener diversity of biofilm bacterial communities on natural substrates were much higher than those on MPs substrates | Lake water for 40-day cultivation | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, C.; Huang, L.; Xie, G.; Wang, Y.; Ma, Z.; Yao, Y.; Yang, H. Effects of Plastic Debris on the Biofilm Bacterial Communities in Lake Water. Water 2021, 13, 1465. https://doi.org/10.3390/w13111465
Shen C, Huang L, Xie G, Wang Y, Ma Z, Yao Y, Yang H. Effects of Plastic Debris on the Biofilm Bacterial Communities in Lake Water. Water. 2021; 13(11):1465. https://doi.org/10.3390/w13111465
Chicago/Turabian StyleShen, Chao, Liuyan Huang, Guangwu Xie, Yulai Wang, Zongkai Ma, Yu Yao, and Hong Yang. 2021. "Effects of Plastic Debris on the Biofilm Bacterial Communities in Lake Water" Water 13, no. 11: 1465. https://doi.org/10.3390/w13111465
APA StyleShen, C., Huang, L., Xie, G., Wang, Y., Ma, Z., Yao, Y., & Yang, H. (2021). Effects of Plastic Debris on the Biofilm Bacterial Communities in Lake Water. Water, 13(11), 1465. https://doi.org/10.3390/w13111465