
Effects of Macrofaunal Recolonization on Biogeochemical Processes and Microbiota—A Mesocosm Study

 , , and
, , and 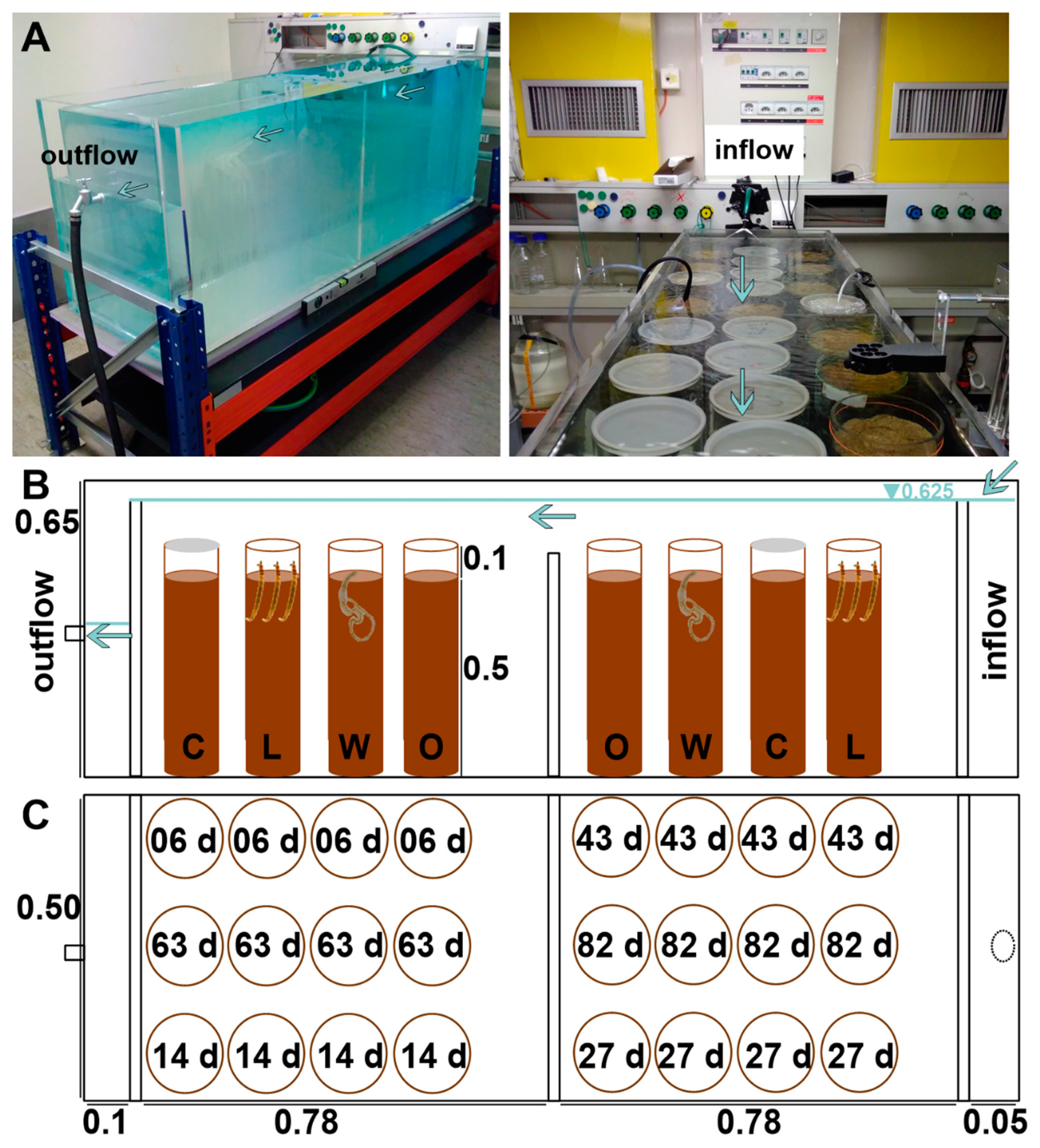{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Core and Macrofaunal Sampling
2.2. Experimental Design
2.3. Monitoring and Sampling
2.4. Analyses
2.4.1. Microsensor Analyses
2.4.2. Pore Water Analyses
2.4.3. Solid-Phase Analyses
2.4.4. Bioinformatic Analyses
2.4.5. Statistical Analyses
3. Results
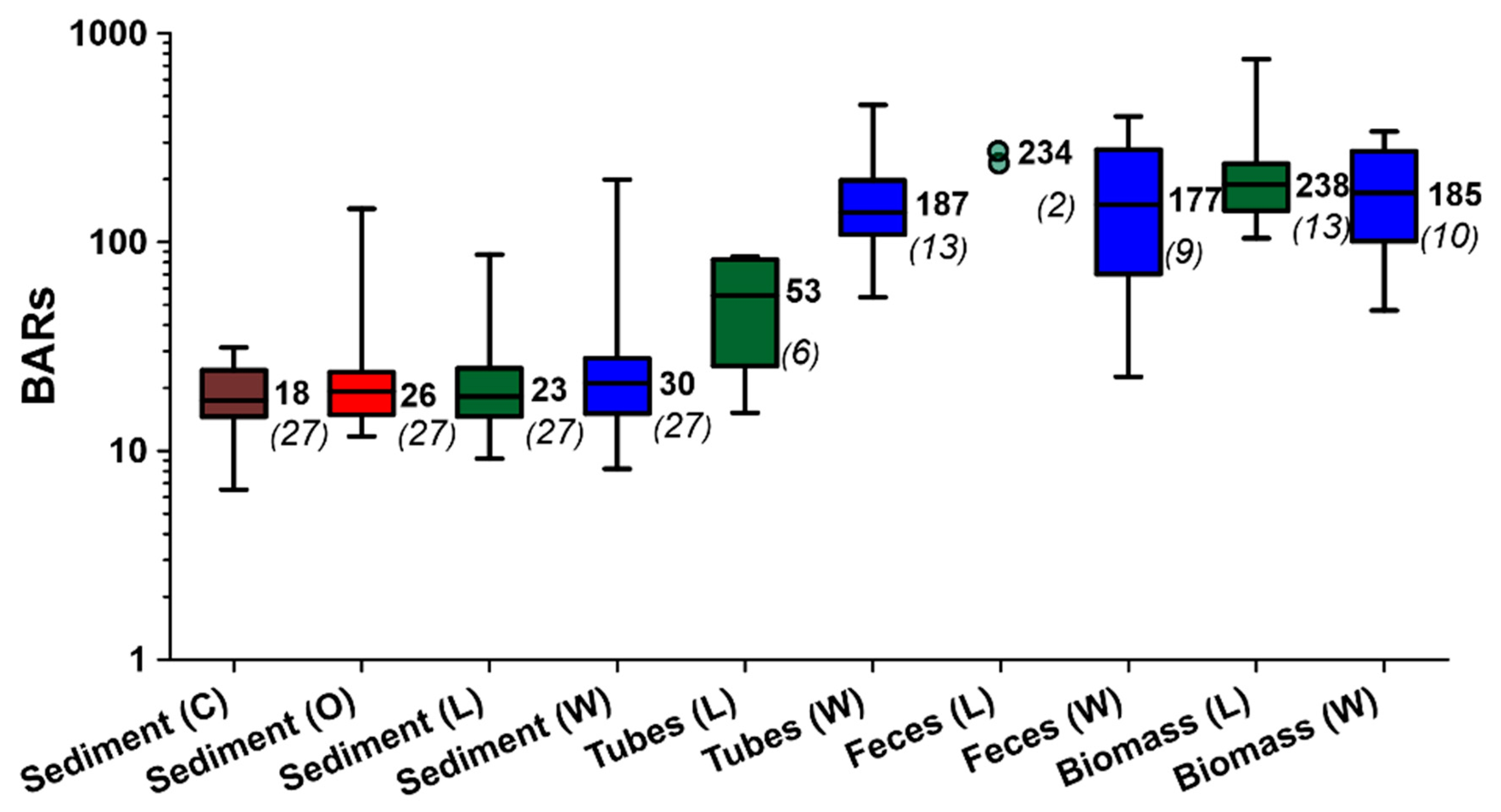
3.1. Visual Observations
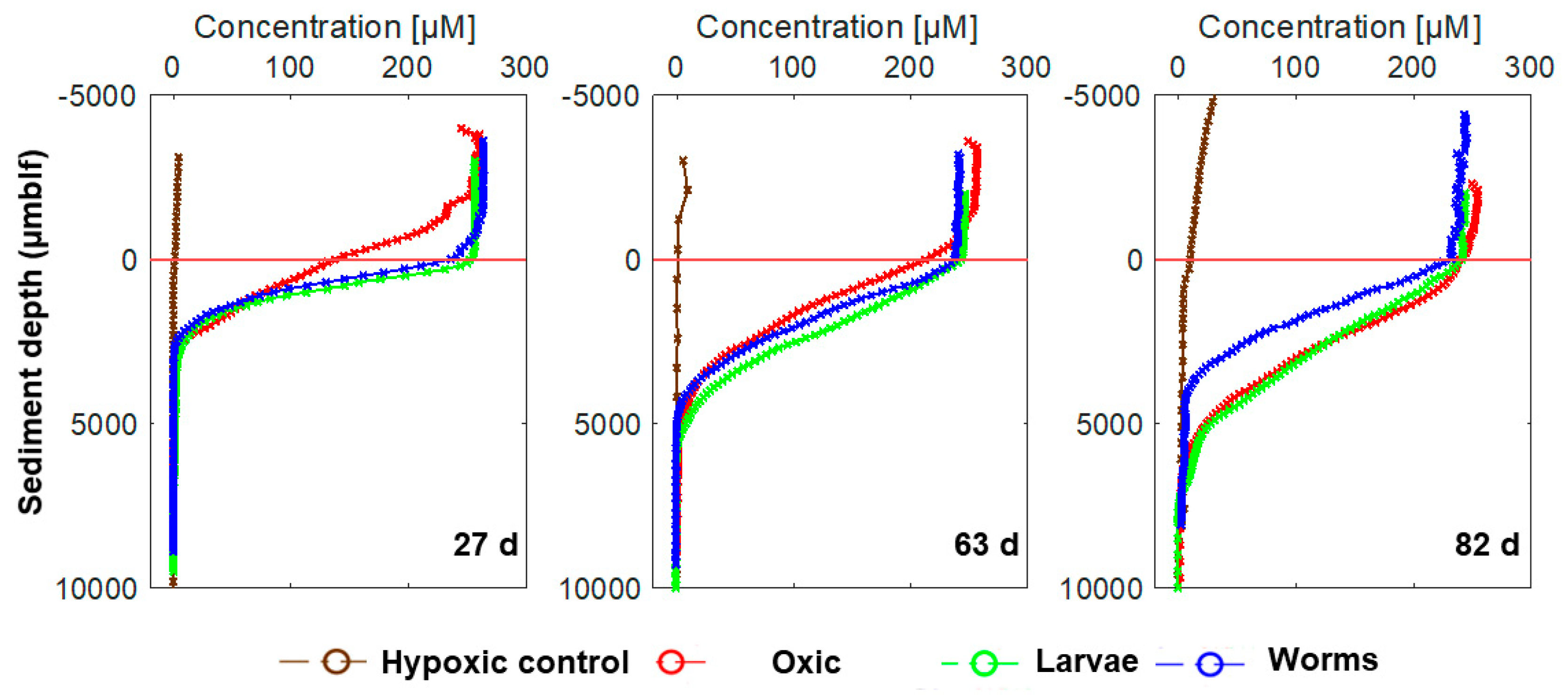
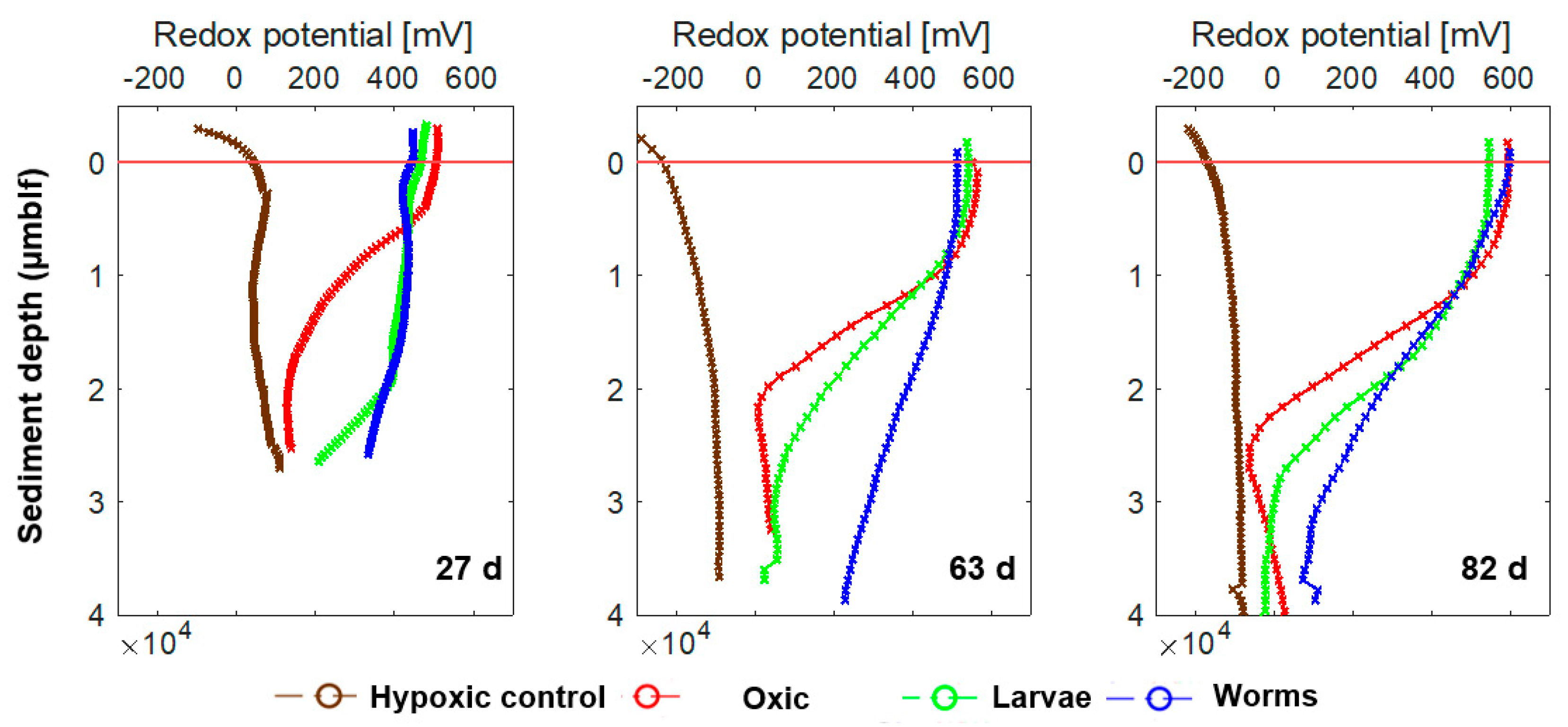
3.2. Microsensor Measurements of O2 Concentrations and Redox Potential
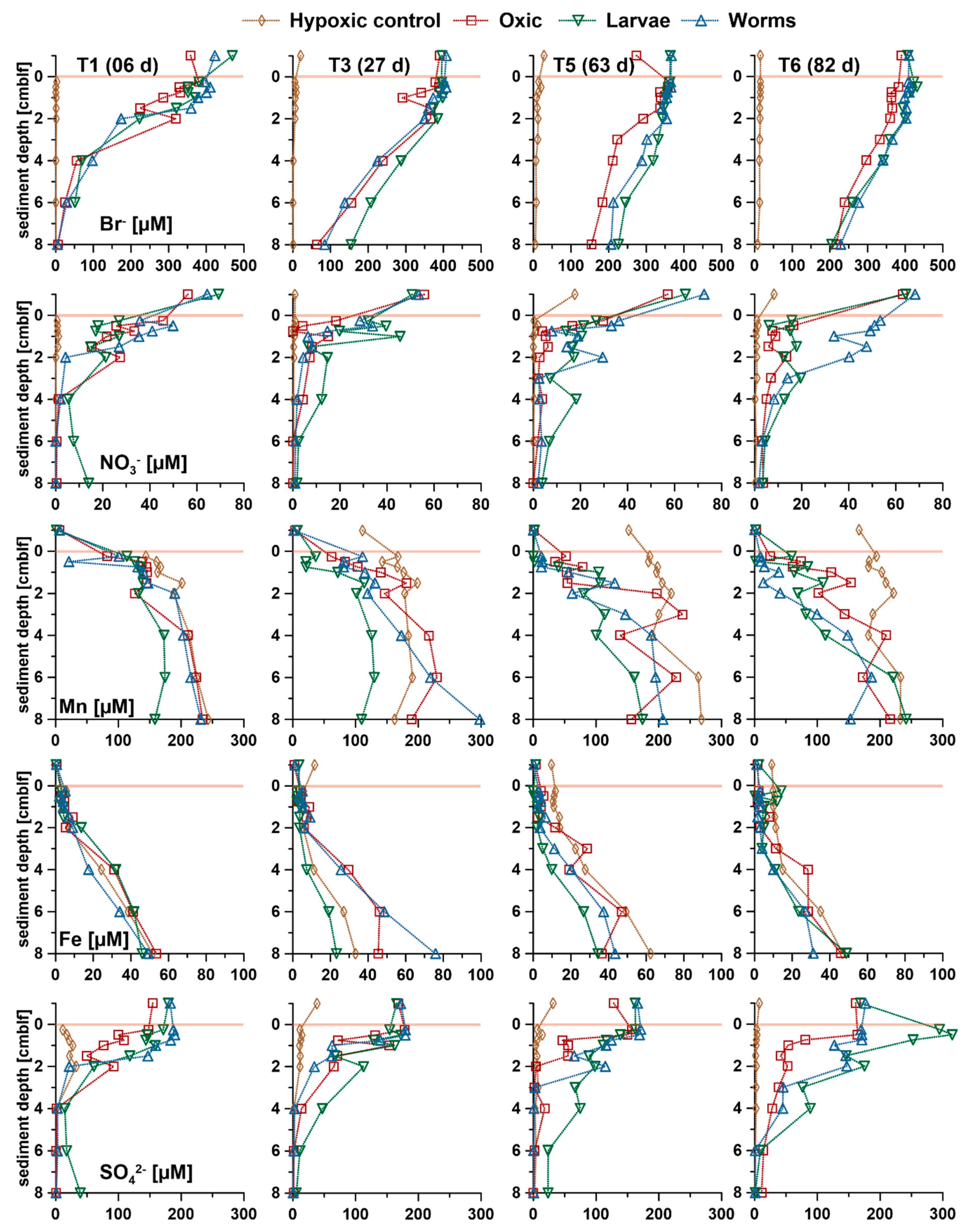
3.3. Pore Water Concentrations of Anions and Cations
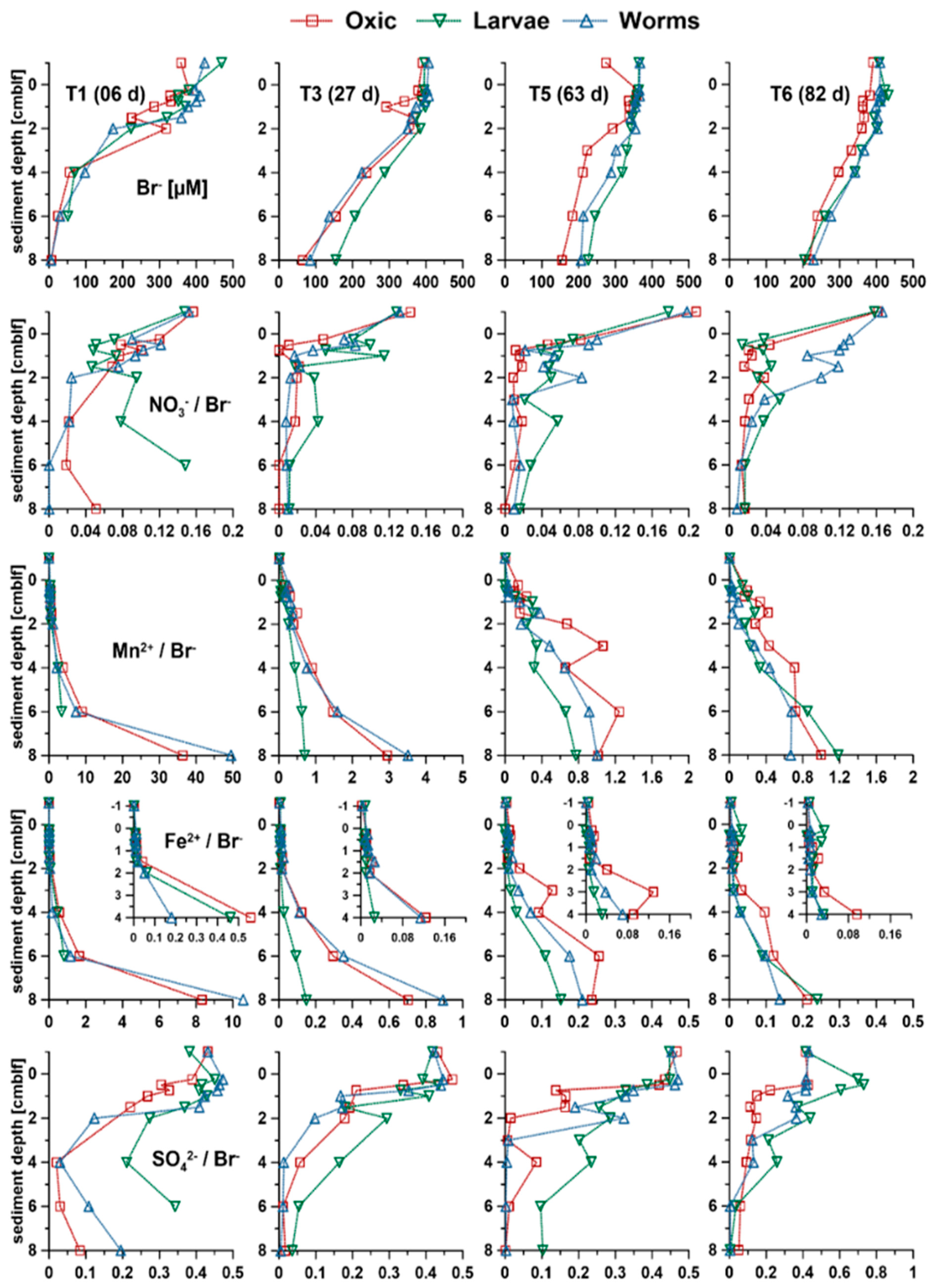
3.3.1. Bromide (Br−)
3.3.2. NO3− and SO42−
3.3.3. Mn2+ and Fe2+
3.3.4. NH4+
3.4. Solid Phase Analyses
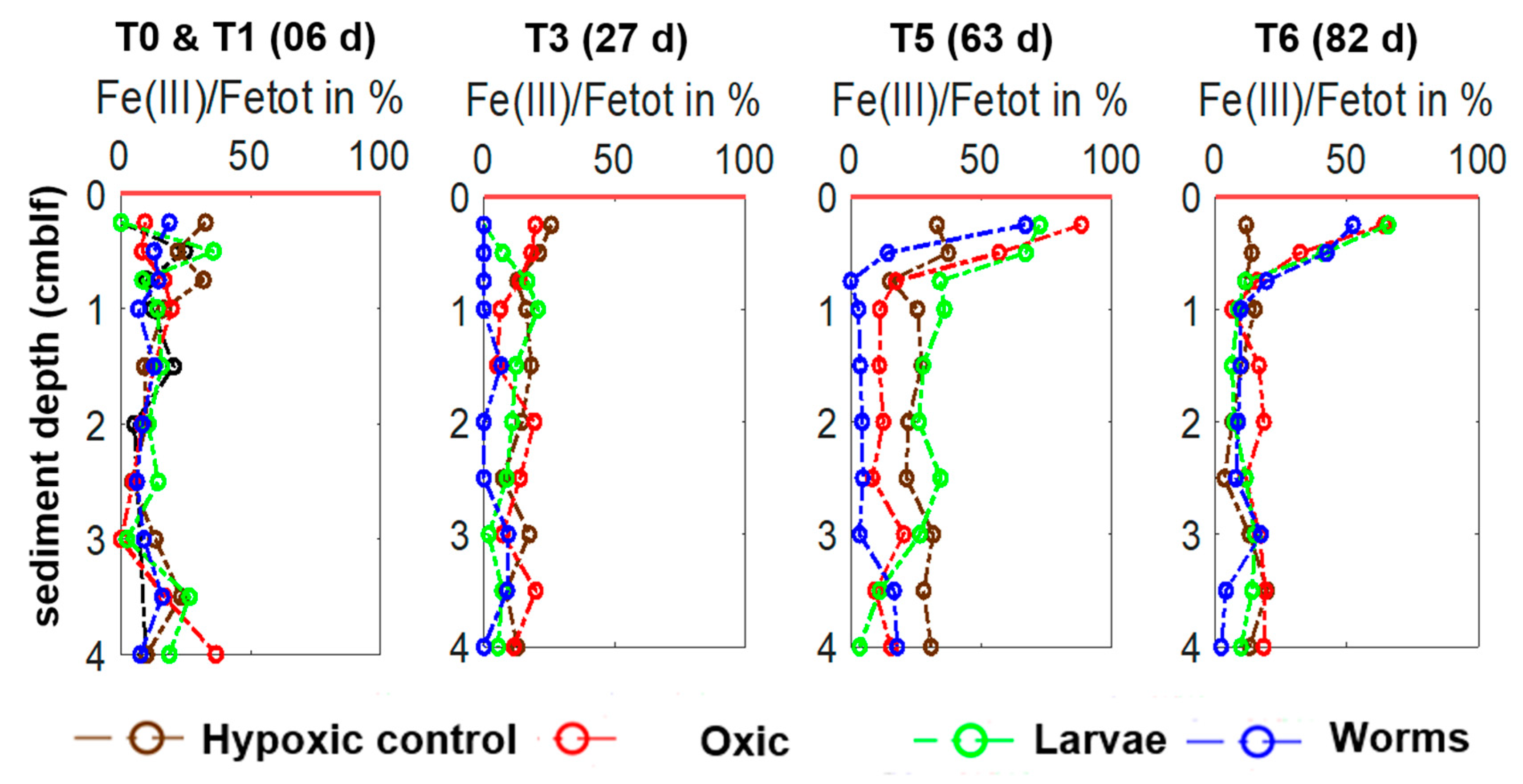
3.4.1. Iron and EAC/EDC
3.4.2. Luminophores and Chlorophyll a
3.5. Microbial Community Composition
3.5.1. qPCR Values of Bacterial and Archaeal 16S rRNA Genes
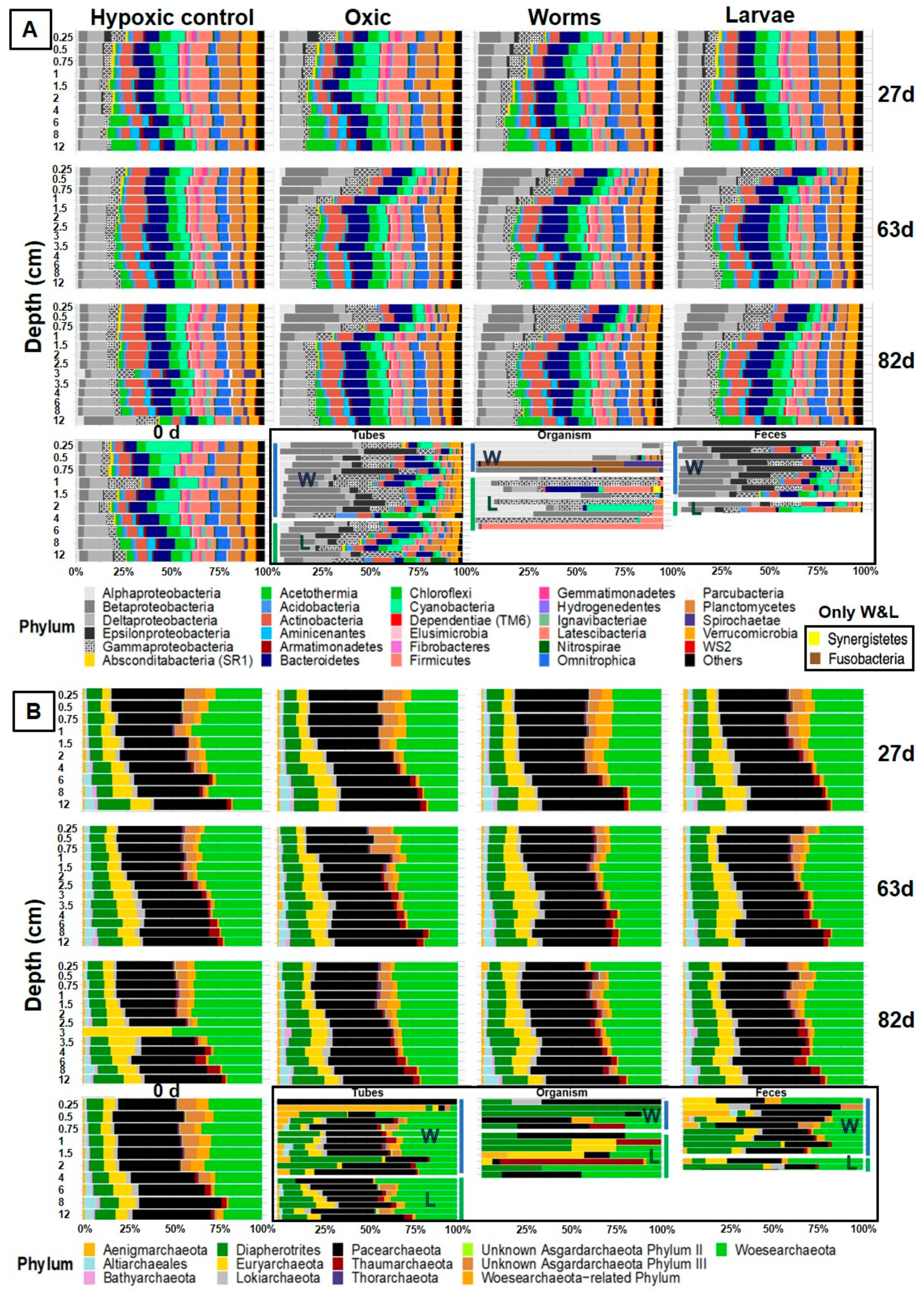
3.5.2. Amplicon Sequencing of 16S rRNA
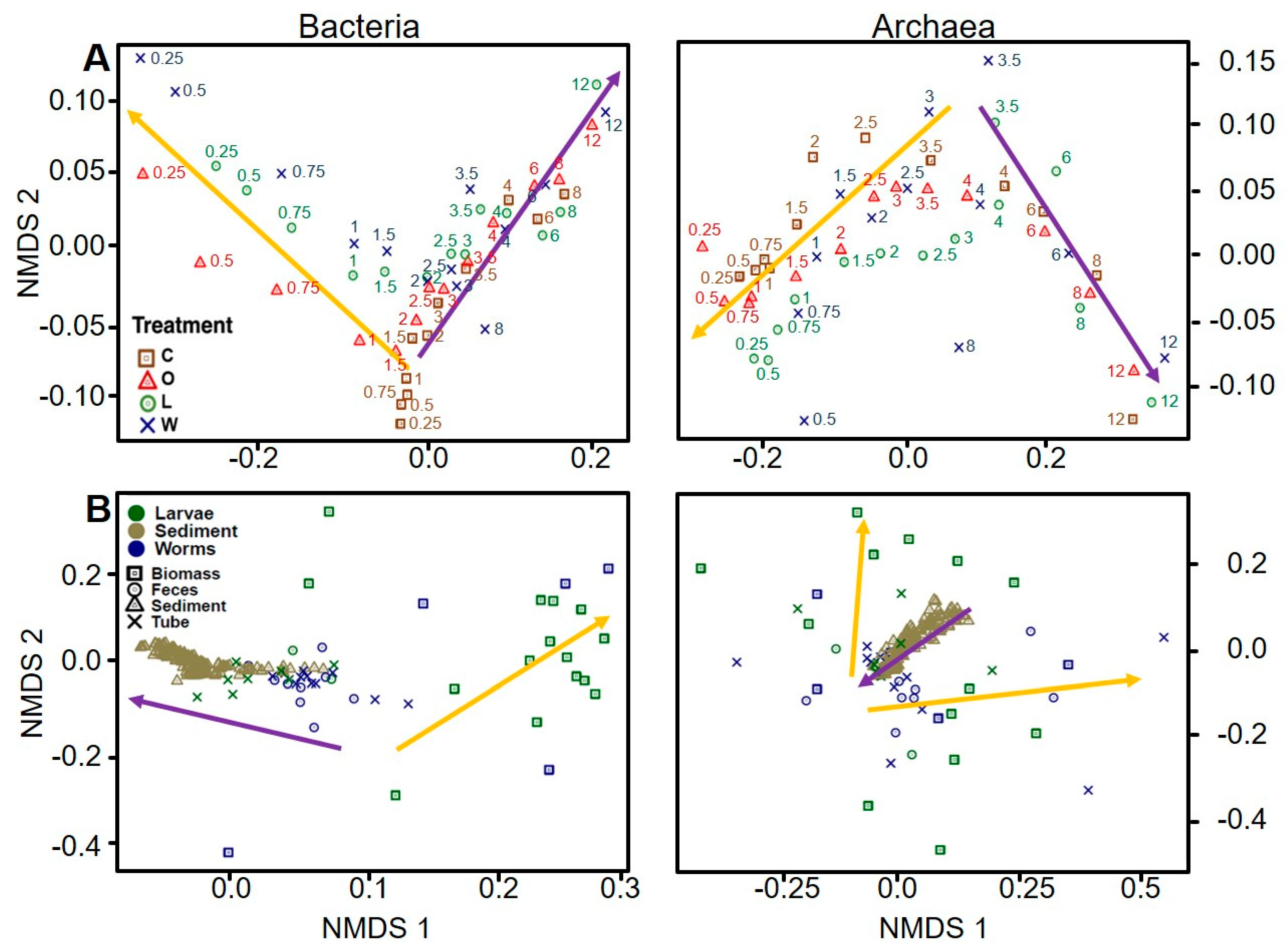
3.5.3. Sediment Microbial Community Zonation Patterns across Experimental Treatments
3.5.4. Bacteria
3.5.5. Archaea
3.5.6. Comparison of Microbial Communities in Sediment, Faunal Specimens, Burrow Structures, and Feces
4. Discussion
4.1. The Effect of Bottom Water O2 and Macrofaunal Activity on Sediment Biogeochemistry
4.2. Effect of Bottom Water O2 on Sediment Microbial Communities
4.3. Effect of Chironomid Larval and Worm Bioturbation on Sediment Microbial Communities
4.4. Possible Reasons for Weak Bioturbation Effects
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Kristensen, E.; Penha-Lopes, G.; Delefosse, M.; Valdemarsen, T.; Quintana, C.O.; Banta, G.T. What is bioturbation? The need for a precise definition for fauna in aquatic sciences. Mar. Ecol. Prog. Ser. 2012, 446, 285–302. [Google Scholar] [CrossRef] [Green Version]
- Meysman, F.J.R.; Middelburg, J.J.; Heip, C.H.R. Bioturbation: A fresh look at darwin’s last idea. Trends Ecol. Evol. 2006, 21, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Bertics, V.J.; Ziebis, W. Biodiversity of benthic microbial communities in bioturbated coastal sediments is controlled by geochemical microniches. ISME J. 2009, 3, 1269–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristensen, E.; Holmer, M. Decomposition of plant materials in marine sediment exposed to different electron accepters (o-2, no3-, and so42-), with emphasis on substrate origin, degradation kinetics, and the role of bioturbation. Geochim. Cosmochim. Acta 2001, 65, 419–433. [Google Scholar] [CrossRef]
- Canfield, D.E.; Kristensen, E.; Thamdrup, B. Aquatic geomicrobiology. Adv. Mar. Biol. 2005, 48, 1–640. [Google Scholar] [PubMed]
- Christensen, B.; Vedel, A.; Kristensen, E. Carbon and nitrogen fluxes in sediment inhabited by suspension-feeding (nereis diversicolor) and non-suspension-feeding (n-virens) polychaetes. Mar. Ecol. Prog. Ser. 2000, 192, 203–217. [Google Scholar] [CrossRef]
- Thamdrup, B.; Canfield, D.E. Pathways of carbon oxidation in continental margin sediments off central chile. Limnol. Oceanogr. 1996, 41, 1629–1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aller, R.C.; Aller, J.Y. The effect of biogenic irrigation intensity and solute exchange on diagenetic reaction rates in marine sediments. J. Mar. Res. 1998, 56, 905–936. [Google Scholar] [CrossRef]
- Kajan, R.; Frenzel, P. The effect of chironomid larvae on production, oxidation and fluxes of methane in a flooded rice soil. FEMS Microbiol. Ecol. 1999, 28, 121–129. [Google Scholar] [CrossRef]
- Verdonschot, P.F. Macrofaunal community types in ponds and small lakes (overijssel, the netherlands). Hydrobiologia 1992, 232, 111–132. [Google Scholar] [CrossRef]
- Mermillod-Blondin, F.; Rosenberg, R. Ecosystem engineering: The impact of bioturbation on biogeochemical processes in marine and freshwater benthic habitats. Aquat. Sci. 2006, 68, 434–442. [Google Scholar] [CrossRef]
- McCall, P.L.; Tevesz, M.J.S. The effects of benthos on physical properties of freshwater sediments. In Animal-Sediment Relations: The Biogenic Alteration of Sediments; McCall, P.L., Tevesz, M.J.S., Eds.; Springer: Boston, MA, USA, 1982; pp. 105–176. [Google Scholar]
- Pinder, L. The habitats of chironomid larvae. In The Chironomidae; Springer: Berlin/Heidelberg, Germany, 1995; pp. 107–135. [Google Scholar]
- Armitage, P.D.; Pardo, I.; Brown, A. Temporal constancy of faunal assemblages in mesohabitats—Application to management. Arch. Hydrobiol. 1995, 133, 367–387. [Google Scholar] [CrossRef]
- Holker, F.; Vanni, M.J.; Kuiper, J.J.; Meile, C.; Grossart, H.P.; Stief, P.; Adrian, R.; Lorke, A.; Dellwig, O.; Brand, A.; et al. Tube-dwelling invertebrates: Tiny ecosystem engineers have large effects in lake ecosystems. Ecol. Monogr. 2015, 85, 333–351. [Google Scholar] [CrossRef] [Green Version]
- Stief, P.; Holker, F. Trait-mediated indirect effects of predatory fish on microbial mineralization in aquatic sediments. Ecology 2006, 87, 3152–3159. [Google Scholar] [CrossRef]
- Lewandowski, J.; Laskov, C.; Hupfer, M. The relationship between chironomus plumosus burrows and the spatial distribution of pore-water phosphate, iron and ammonium in lake sediments. Freshw. Biol. 2007, 52, 331–343. [Google Scholar] [CrossRef]
- Hupfer, M.; Jordan, S.; Herzog, C.; Ebeling, C.; Ladwig, R.; Rothe, M.; Lewandowski, J. Chironomid larvae enhance phosphorus burial in lake sediments: Insights from long-term and short-term experiments. Sci. Total Environ. 2019, 663, 254–264. [Google Scholar] [CrossRef]
- Pelegri, S.P.; Nielsen, L.P.; Blackburn, T.H. Denitrification in estuarine sediment stimulated by the irrigation activity of the amphipod corophium-volutator. Mar. Ecol. Prog. Ser. 1994, 105, 285–290. [Google Scholar] [CrossRef]
- Svensson, J.M.; Enrich-Prast, A.; Leonardson, L. Nitrification and denitrification in a eutrophic lake sediment bioturbated by oligochaetes. Aquat. Microb. Ecol. 2001, 23, 177–186. [Google Scholar] [CrossRef]
- Timm, T.; Martin, P.J. Clitellata: Oligochaeta. In Ecology and General Biology, Vol I: Thorp and Covich’s Freshwater Invertebrates, 4th ed.; Academic Press: Cambridge, MA, USA, 2015; pp. 529–549. [Google Scholar]
- Davis, R.B. Tubificids alter profiles of redox potential and ph in profundal lake sediment 1. Limnol. Oceanogr. 1974, 19, 342–346. [Google Scholar] [CrossRef]
- Gautreau, E.; Volatier, L.; Nogaro, G.; Gouze, E.; Mermillod-Blondin, F. The influence of bioturbation and water column oxygenation on nutrient recycling in reservoir sediments. Hydrobiologia 2020, 847, 1027–1040. [Google Scholar] [CrossRef]
- Mermillod-Blondin, F.; Nogaro, G.; Datry, T.; Malard, F.; Gibert, J. Do tubificid worms influence the fate of organic matter and pollutants in stormwater sediments? Environ. Pollut. 2005, 134, 57–69. [Google Scholar] [CrossRef]
- Anderson, N.J.; Dietz, R.D.; Engstrom, D.R. Land-use change, not climate, controls organic carbon burial in lakes. Proc. R. Soc. B Biol. Sci. 2013, 280, 20131278. [Google Scholar] [CrossRef] [Green Version]
- Anderson, N.J.; Bennion, H.; Lotter, A.F. Lake eutrophication and its implications for organic carbon sequestration in europe. Glob. Chang. Biol. 2014, 20, 2741–2751. [Google Scholar] [CrossRef] [Green Version]
- Heathcote, A.J.; Downing, J.A. Impacts of eutrophication on carbon burial in freshwater lakes in an intensively agricultural landscape. Ecosystems 2012, 15, 60–70. [Google Scholar] [CrossRef]
- Chapman, P.M.; Farrell, M.A.; Brinkhurst, R.O. Relative tolerances of selected aquatic oligochaetes to individual pollutants and environmental-factors. Aquat. Toxicol. 1982, 2, 47–67. [Google Scholar] [CrossRef]
- Chapman, P.M.; Farrell, M.A.; Brinkhurst, R.O. Effects of species interactions on the survival and respiration of limnodrilus-hoffmeisteri and tubifex-tubifex (oligochaeta, tubificidae) exposed to various pollutants and environmental-factors. Water Res. 1982, 16, 1405–1408. [Google Scholar] [CrossRef]
- Chapman, P.M.; Farrell, M.A.; Brinkhurst, R.O. Relative tolerances of selected aquatic oligochaetes to combinations of pollutants and environmental-factors. Aquat. Toxicol. 1982, 2, 69–78. [Google Scholar] [CrossRef]
- Nicacio, G.; Juen, L. Chironomids as indicators in freshwater ecosystems: An assessment of the literature. Insect Conserv. Divers. 2015, 8, 393–403. [Google Scholar] [CrossRef] [Green Version]
- van de Bund, W.J.; Goedkoop, W.; Johnson, R.K. Effects of deposit-feeder activity on bacterial production and abundance in profundal lake sediment. J. N. Am. Benthol. Soc. 1994, 13, 532–539. [Google Scholar] [CrossRef]
- Wieltschnig, C.; Fischer, U.R.; Velimirov, B.; Kirschner, A.K. Effects of deposit-feeding macrofauna on benthic bacteria, viruses, and protozoa in a silty freshwater sediment. Microb. Ecol. 2008, 56, 1–12. [Google Scholar] [CrossRef]
- Zeng, J.; Zhao, D.-Y.; Liu, P.; Yu, Z.-B.; Huang, R.; Wu, Q.L. Effects of benthic macrofauna bioturbation on the bacterial community composition in lake sediments. Can. J. Microbiol. 2014, 60, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Hunting, E.R.; Whatley, M.H.; van der Geest, H.G.; Mulder, C.; Kraak, M.H.S.; Breure, A.M.; Admiraal, W. Invertebrate footprints on detritus processing, bacterial community structure, and spatiotemporal redox profiles. Freshw. Sci. 2012, 31, 724–732. [Google Scholar] [CrossRef] [Green Version]
- Fiskal, A.; Deng, L.; Michel, A.; Eickenbusch, P.; Han, X.; Lagostina, L.; Zhu, R.; Sander, M.; Schroth, M.; Bernasconi, S.; et al. Effects of eutrophication on sedimentary organic carbon cycling in five temperate lakes. Biogeosciences 2019, 16, 3725–3746. [Google Scholar] [CrossRef] [Green Version]
- Fiskal, A.; Anthamatten, E.; Deng, L.; Han, X.; Lagostina, L.; Michel, A.; Zhu, R.; Dubois, N.; Schubert, C.J.; Bernasconi, S.M.; et al. Carbon sources of benthic fauna in temperate lakes across multiple trophic states. Biogeosci. Discuss. 2020, 2020, 1–26. [Google Scholar]
- Mahaut, M.-L.; Graf, G. A luminophore tracer technique for bioturbation studies. Oceanol. Acta Paris 1987, 10, 323–328. [Google Scholar]
- Kempers, A.; Kok, C. Re-examination of the determination of ammonium as the indophenol blue complex using salicylate. Anal. Chim. Acta 1989, 221, 147–155. [Google Scholar] [CrossRef]
- Lovley, D.R.; Phillips, E.J.P. Rapid assay for microbially reducible ferric iron in aquatic sediments. Appl. Environ. Microb. 1987, 53, 1536–1540. [Google Scholar] [CrossRef] [Green Version]
- Kluepfel, L.; Keiluweit, M.; Kleber, M.; Sander, M. Redox properties of plant biomass-derived black carbon (biochar). Environ. Sci. Technol. 2014, 48, 5601–5611. [Google Scholar] [CrossRef] [PubMed]
- Sander, M.; Hofstetter, T.B.; Gorski, C.A. Electrochemical analyses of redox-active iron minerals: A review of nonmediated and mediated approaches. Environ. Sci. Technol. 2015, 49, 5862–5878. [Google Scholar] [CrossRef] [Green Version]
- Lever, M.A.; Valiela, I. Response of microphytobenthic biomass to experimental nutrient enrichment and grazer exclusion at different land-derived nitrogen loads. Mar. Ecol. Prog. Ser. 2005, 294, 117–129. [Google Scholar] [CrossRef] [Green Version]
- Lorenzen, C.J. Determination of chlorophyll and pheo-pigments—spectrophotometric equations. Limnol. Oceanogr. 1967, 12, 343–346. [Google Scholar] [CrossRef]
- Lever, M.A.; Torti, A.; Eickenbusch, P.; Michaud, A.B.; Santl-Temkiv, T.; Jorgensen, B.B. A modular method for the extraction of DNA and rna, and the separation of DNA pools from diverse environmental sample types. Front. Microbiol. 2015, 6, 476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starnawski, P.; Bataillon, T.; Ettema, T.J.G.; Jochum, L.M.; Schreiber, L.; Chen, X.; Lever, M.A.; Polz, M.F.; Jorgensen, B.B.; Schramm, A.; et al. Microbial community assembly and evolution in subseafloor sediment. Proc. Natl. Acad. Sci. USA 2017, 114, 2940–2945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claesson, M.J.; O’Sullivan, O.; Wang, Q.; Nikkila, J.; Marchesi, J.R.; Smidt, H.; de Vos, W.M.; Ross, R.P.; O’Toole, P.W. Comparative analysis of pyrosequencing and a phylogenetic microarray for exploring microbial community structures in the human distal intestine. PLoS ONE 2009, 4, e6669. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012; Available online: http://www.R-project.org (accessed on 20 February 2018).
- Han, X.; Schubert, C.J.; Fiskal, A.; Dubois, N.; Lever, M.A. Eutrophication as a driver of microbial community structure in lake sediments. Environ. Microbiol. 2020, 22, 3446–3462. [Google Scholar] [CrossRef] [PubMed]
- Baranov, V.; Lewandowski, J.; Krause, S. Bioturbation enhances the aerobic respiration of lake sediments in warming lakes. Biol. Lett. 2016, 12, 269–281. [Google Scholar] [CrossRef]
- Lagauzere, S.; Moreira, S.; Koschorreck, M. Influence of bioturbation on the biogeochemistry of littoral sediments of an acidic post-mining pit lake. Biogeosciences 2011, 8, 339–352. [Google Scholar] [CrossRef]
- Klupfel, L.; Piepenbrock, A.; Kappler, A.; Sander, M. Humic substances as fully regenerable electron acceptors in recurrently anoxic environments. Nat. Geosci. 2014, 7, 195–200. [Google Scholar] [CrossRef]
- Walpen, N.; Getzinger, G.J.; Schroth, M.H.; Sander, M. Electron-donating phenolic and electron-accepting quinone moieties in peat dissolved organic matter: Quantities and redox transformations in the context of peat biogeochemistry. Environ. Sci. Technol. 2018, 52, 5236–5245. [Google Scholar] [CrossRef]
- Stove Poindexter, J.L.; Cohen-Bazire, G. The fine structure of stalked bacteria belonging to the family caulobacteraceae. J. Cell Biol. 1964, 23, 587–607. [Google Scholar] [CrossRef] [PubMed]
- Garrity, G.M.; Bell, J.A.; Lilburn, T. Caulobacteraceae. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2015; p. 1. [Google Scholar] [CrossRef]
- Spring, S.; Scheuner, C.; Göker, M.; Klenk, H.-P. A taxonomic framework for emerging groups of ecologically important marine gammaproteobacteria based on the reconstruction of evolutionary relationships using genome-scale data. Front. Microbiol. 2015, 6, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kämpfer, P. Sphingobacteriia class. Nov. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2015; p. 1. [Google Scholar] [CrossRef]
- Orata, F.D.; Meier-Kolthoff, J.P.; Sauvageau, D.; Stein, L.Y. Phylogenomic analysis of the gammaproteobacterial methanotrophs (order methylococcales) calls for the reclassification of members at the genus and species levels. Front. Microbiol. 2018, 9, 3162. [Google Scholar] [CrossRef] [Green Version]
- Bowman, J.P. Methylococcales. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2018; pp. 1–4. [Google Scholar] [CrossRef]
- Kowalchuk, G.A.; Stephen, J.R. Ammonia-oxidizing bacteria: A model for molecular microbial ecology. Annu. Rev. Microbiol. 2001, 55, 485–529. [Google Scholar] [CrossRef] [Green Version]
- Spieck, E.; Bock, E. Nitrospira. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2015; pp. 1–4. [Google Scholar] [CrossRef]
- Willems, A. The family comamonadaceae. In The Prokaryotes: Alphaproteobacteria and Betaproteobacteria; Springer: New York, NY, USA, 2013; pp. 777–851. [Google Scholar]
- Chen, Q.; Ni, J. Heterotrophic nitrification–aerobic denitrification by novel isolated bacteria. J. Ind. Microbiol. Biot. 2011, 38, 1305–1310. [Google Scholar] [CrossRef]
- Liou, J.S.-C.; DeRito, C.M.; Madsen, E.L. Field-based and laboratory stable isotope probing surveys of the identities of both aerobic and anaerobic benzene-metabolizing microorganisms in freshwater sediment. Environ. Microbiol. 2008, 10, 1964–1977. [Google Scholar] [CrossRef] [PubMed]
- Garrity, G.M.; Bell, J.A.; Lilburn, T. Thiotrichaceae fam. Nov. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2015; p. 1. [Google Scholar] [CrossRef]
- Krieg, N.R. Bacteroidales ord. Nov. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2015; p. 1. [Google Scholar] [CrossRef]
- Glöckner, J.; Kube, M.; Shrestha, P.M.; Weber, M.; Glöckner, F.O.; Reinhardt, R.; Liesack, W. Phylogenetic diversity and metagenomics of candidate division op3. Environ. Microbiol. 2010, 12, 1218–1229. [Google Scholar] [CrossRef]
- Yamada, T.; Sekiguchi, Y. Anaerolineae. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2018; pp. 1–2. [Google Scholar] [CrossRef]
- Krzmarzick, M.J.; McNamara, P.J.; Crary, B.B.; Novak, P.J. Abundance and diversity of organohalide-respiring bacteria in lake sediments across a geographical sulfur gradient. FEMS Microbiol. Ecol. 2013, 84, 248–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.Y.; Park, J.H.; Jang, S.H.; Nielsen, L.K.; Kim, J.; Jung, K.S. Fermentative butanol production by clostridia. Biotechnol. Bioeng. 2008, 101, 209–228. [Google Scholar] [CrossRef] [PubMed]
- Farag, I.F.; Youssef, N.H.; Elshahed, M.S. Global distribution patterns and pangenomic diversity of the candidate phylum “latescibacteria” (ws3). Appl. Environ. Microb. 2017, 83, e00521-17. [Google Scholar] [CrossRef] [Green Version]
- Farag, I.F. Exploring the Habitat Distribution, Metabolic Diversities and Potential Ecological Roles of Candidate Phyla “Aminicenantes” (op8) and “Latescibacteria”(ws3). Master’s Thesis, Ain Shams University, Cairo, Egypt, 2017. [Google Scholar]
- Kuever, J. The family syntrophaceae. In The Prokaryotes: Deltaproteobacteria and Epsilonproteobacteria; Springer: Berlin/Heidelberg, Germany, 2014; pp. 281–288. [Google Scholar]
- Kuever, J.; Rainey, F.A.; Widdel, F. Desulfarculaceae fam. Nov. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2015; pp. 41–44. [Google Scholar] [CrossRef]
- Hallbeck, L.; Pedersen, K. The family gallionellaceae. Prokaryotes 2014, 853–858. [Google Scholar] [CrossRef]
- Norris, P.R. Acidimicrobiales. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2015; pp. 1–2. [Google Scholar] [CrossRef]
- Lovley, D.R. Microbial fe (iii) reduction in subsurface environments. FEMS Microbiol. Rev. 1997, 20, 305–313. [Google Scholar] [CrossRef]
- Chen, X.H.; Andersen, T.J.; Morono, Y.; Inagaki, F.; Jorgensen, B.B.; Lever, M.A. Bioturbation as a key driver behind the dominance of bacteria over archaea in near-surface sediment. Sci. Rep. UK 2017, 7, 2400. [Google Scholar] [CrossRef]
- Hunting, E.; Kampfraath, A. Contribution of bacteria to redox potential (e h) measurements in sediments. Int. J. Environ. Sci. Technol. 2013, 10, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Hunting, E.; Van der Geest, H. Predictability of bacterial activity and denitrification in aquatic sediments with continuous measurements of redox potential. Int. J. Environ. Sci. Technol. 2011, 8, 553–560. [Google Scholar] [CrossRef] [Green Version]
- Svensson, J.M. Influence of chironomus plumosus larvae on ammonium flux and denitrification (measured by the acetylene blockage- and the isotope pairing-technique) in eutrophic lake sediment. Hydrobiologia 1997, 346, 157–168. [Google Scholar] [CrossRef]
- Samuiloviene, A.; Bartoli, M.; Bonaglia, S.; Cardini, U.; Vybernaite-Lubiene, I.; Marzocchi, U.; Petkuviene, J.; Politi, T.; Zaiko, A.; Zilius, M. The effect of chironomid larvae on nitrogen cycling and microbial communities in soft sediments. Water 2019, 11, 1931. [Google Scholar] [CrossRef] [Green Version]
- Carpintero Moraes, P.; Zilius, M.; Benelli, S.; Bartoli, M. Nitrification and denitrification in estuarine sediments with tube-dwelling benthic animals. Hydrobiologia 2018, 819, 217–230. [Google Scholar] [CrossRef]
- Pelegrí, S.P.; Blackburn, T.H. Effects of tubifex tubifex (oligochaeta: Tubificidae) on n-mineralization in freshwater sediments, measured with 15n isotopes. Aquat. Microb. Ecol. 1995, 9, 289–294. [Google Scholar] [CrossRef]
- Jones, R.I.; Carter, C.E.; Kelly, A.; Ward, S.; Kelly, D.J.; Grey, J. Widespread contribution of methane-cycle bacteria to the diets of lake profundal chironomid larvae. Ecology 2008, 89, 857–864. [Google Scholar] [CrossRef]
- Moog, O. Fauna Aquatica Austriaca, Edition 2002, Wassserwirtschaftskataster, Bundesministerium für Land und Forstwirtschaft; Umwelt und Wasserwirtschaft Vienna: Vienna, Austria, 2002.
- Lagauzère, S.; Boyer, P.; Stora, G.; Bonzom, J.-M. Effects of uranium-contaminated sediments on the bioturbation activity of chironomus riparius larvae (insecta, diptera) and tubifex tubifex worms (annelida, tubificidae). Chemosphere 2009, 76, 324–334. [Google Scholar] [CrossRef]
- Morad, M.R.; Khalili, A.; Roskosch, A.; Lewandowski, J. Quantification of pumping rate of chironomus plumosus larvae in natural burrows. Aquat. Ecol. 2010, 44, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Baranov, V.; Lewandowski, J.; Romeijn, P.; Singer, G.; Krause, S. Effects of bioirrigation of non-biting midges (diptera: Chironomidae) on lake sediment respiration. Sci. Rep. UK 2016, 6, 27329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matisoff, G.; Wang, X. Solute transport in sediments by freshwater infaunal bioirrigators. Limnol. Oceanogr. 1998, 43, 1487–1499. [Google Scholar] [CrossRef] [Green Version]
- Matisoff, G.; Wang, X.S.; McCall, P.L. Biological redistribution of lake sediments by tubificid oligochaetes: Branchiura sowerbyi and limnodrilus hoffmeisteri/tubifex tubifex. J. Great Lakes Res. 1999, 25, 205–219. [Google Scholar] [CrossRef]
- Fisher, J.; Lick, W.; McCall, P.; Robbins, J. Vertical mixing of lake sediments by tubificid oligochaetes. J. Geophys. Res. Ocean. 1980, 85, 3997–4006. [Google Scholar] [CrossRef]
- Chen, M.; Ding, S.; Liu, L.; Wang, Y.; Xing, X.; Wang, D.; Gong, M.; Zhang, C. Fine-scale bioturbation effects of tubificid worm (limnodrilus hoffmeisteri) on the lability of phosphorus in sediments. Environ. Pollut. 2016, 219, 604–611. [Google Scholar] [CrossRef]
- Franzmann, P.D.; Liu, Y.; Balkwill, D.L.; Aldrich, H.C.; Conway De Macario, E.; Boone, D.R. Methanogenium frigidum sp. Nov., a psychrophilic, h2-using methanogen from ace lake, antarctica. Int. J. Syst. Evol. Microbiol. 1997, 47, 1068–1072. [Google Scholar] [CrossRef] [Green Version]
- Mailloux, B.J.; Fuller, M.E. Determination of in situ bacterial growth rates in aquifers and aquifer sediments. Appl. Environ. Microb. 2003, 69, 3798–3808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nauhaus, K.; Albrecht, M.; Elvert, M.; Boetius, A.; Widdel, F. In vitro cell growth of marine archaeal-bacterial consortia during anaerobic oxidation of methane with sulfate. Environ. Microbiol. 2007, 9, 187–196. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fiskal, A.; Gaillard, A.; Giroud, S.; Malcic, D.; Joshi, P.; Sander, M.; Schubert, C.J.; Lever, M.A. Effects of Macrofaunal Recolonization on Biogeochemical Processes and Microbiota—A Mesocosm Study. Water 2021, 13, 1599. https://doi.org/10.3390/w13111599
Fiskal A, Gaillard A, Giroud S, Malcic D, Joshi P, Sander M, Schubert CJ, Lever MA. Effects of Macrofaunal Recolonization on Biogeochemical Processes and Microbiota—A Mesocosm Study. Water. 2021; 13(11):1599. https://doi.org/10.3390/w13111599
Chicago/Turabian StyleFiskal, Annika, Aixala Gaillard, Sebastien Giroud, Dejan Malcic, Prachi Joshi, Michael Sander, Carsten J. Schubert, and Mark Alexander Lever. 2021. "Effects of Macrofaunal Recolonization on Biogeochemical Processes and Microbiota—A Mesocosm Study" Water 13, no. 11: 1599. https://doi.org/10.3390/w13111599
APA StyleFiskal, A., Gaillard, A., Giroud, S., Malcic, D., Joshi, P., Sander, M., Schubert, C. J., & Lever, M. A. (2021). Effects of Macrofaunal Recolonization on Biogeochemical Processes and Microbiota—A Mesocosm Study. Water, 13(11), 1599. https://doi.org/10.3390/w13111599