
Picocyanobacterial Contribution to the Total Primary Production in the Northwestern Pacific Ocean

 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
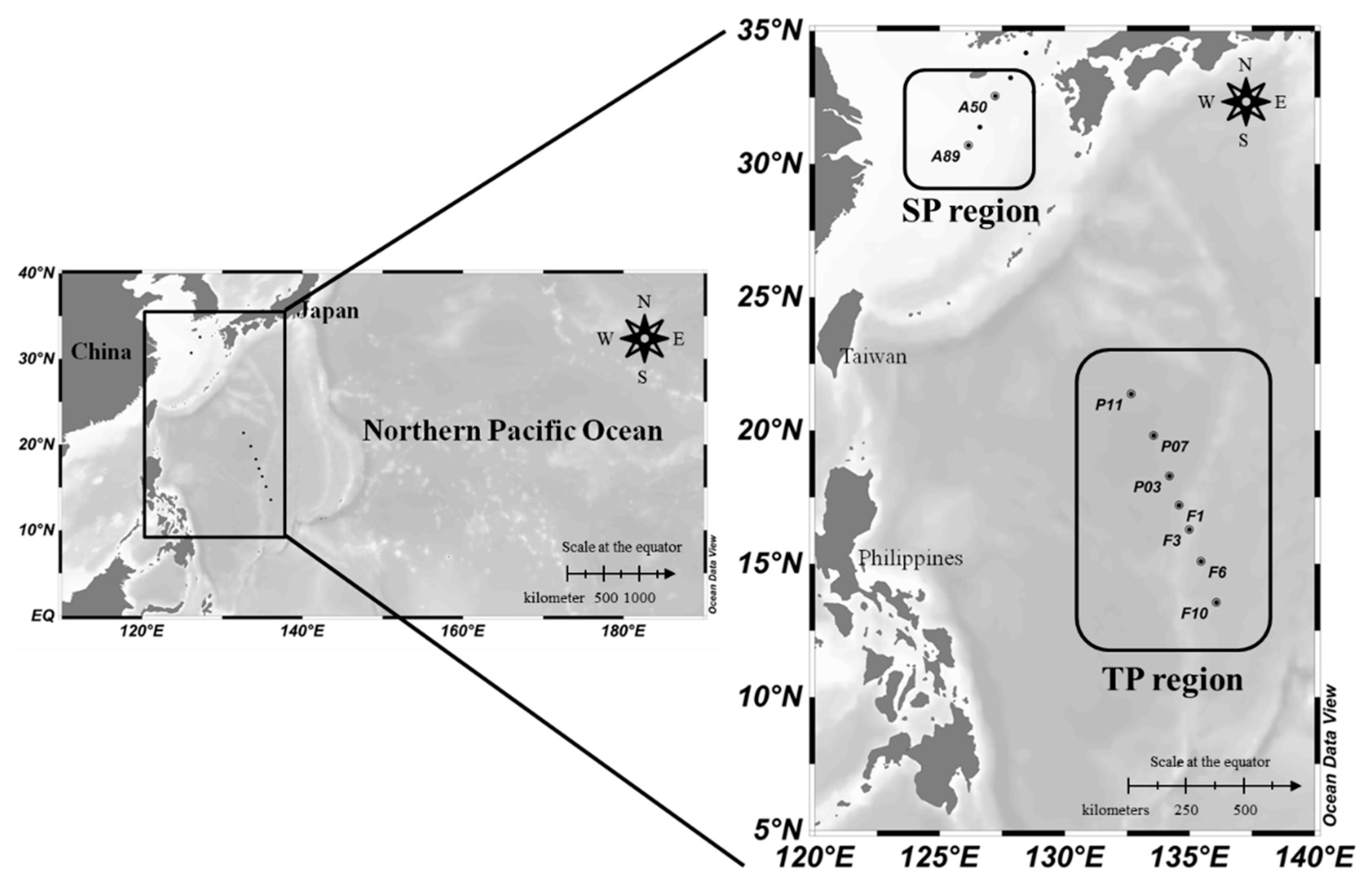
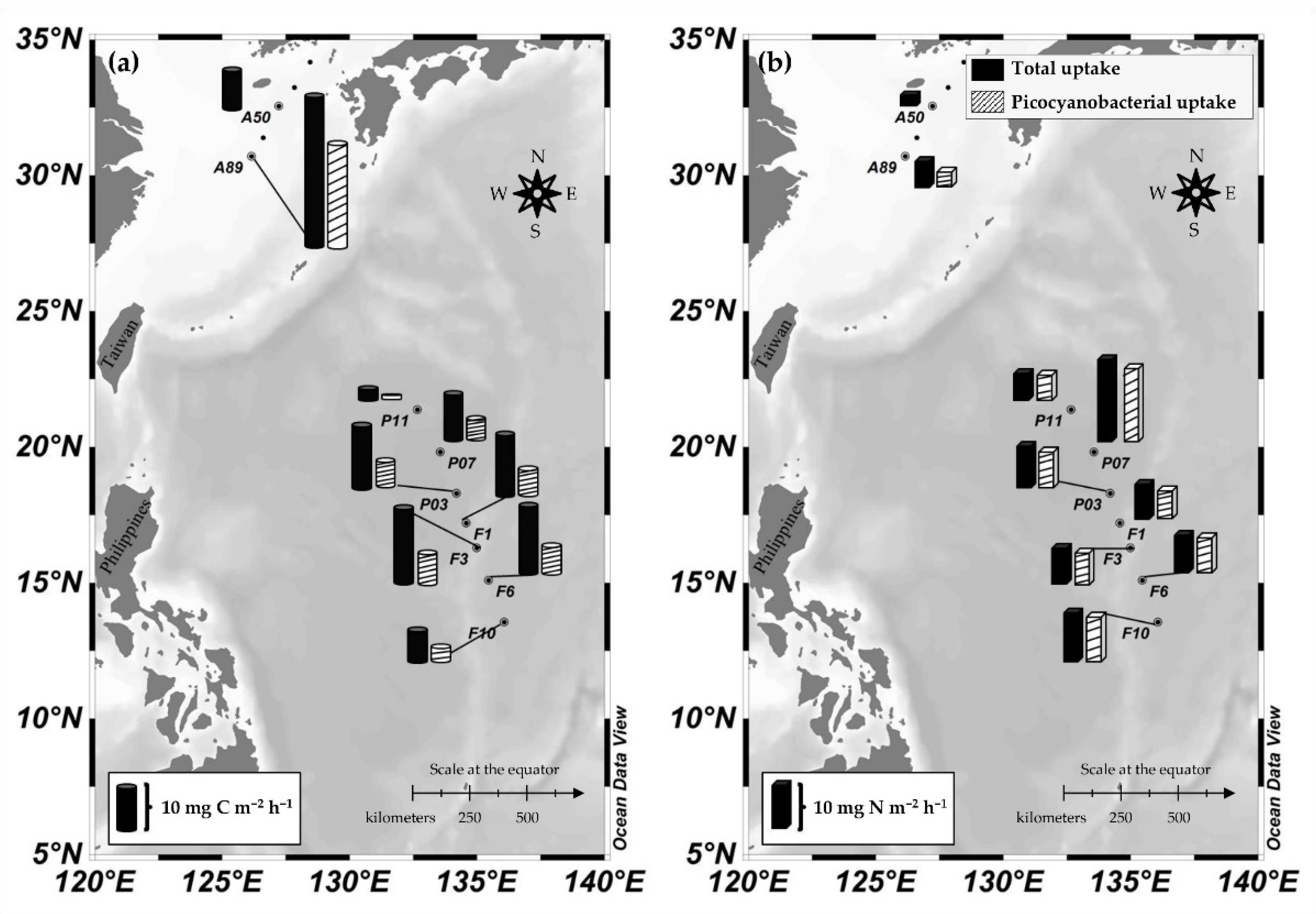
2.1. Study Area and Sample Collection
2.2. Measurements for Biomass and Abundance of Phytoplankton and Nutrient Concentrations
2.3. Carbon and Nitrogen Uptake Rate Measurements
3. Results
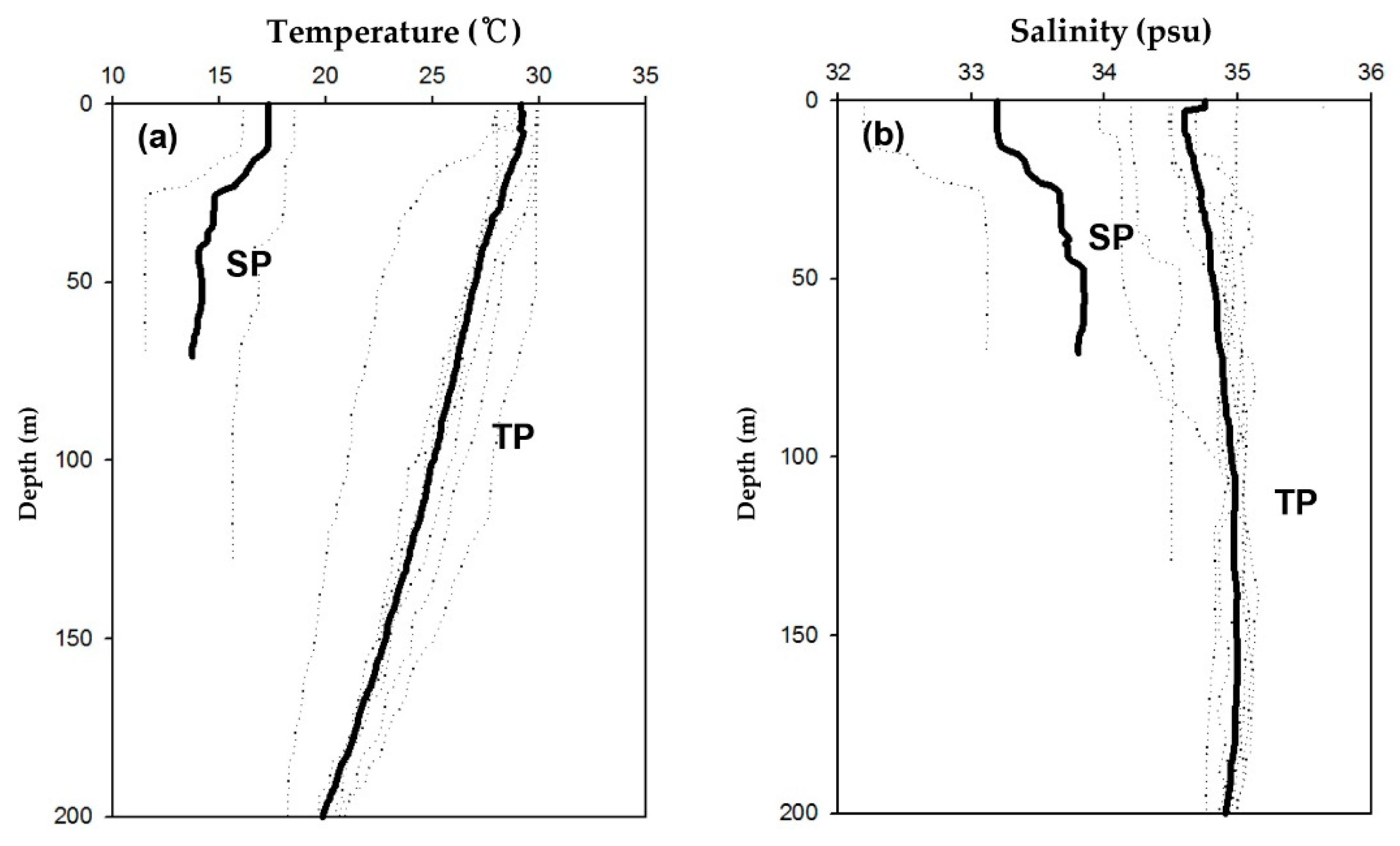
3.1. Physiochemical Structures in Water Column
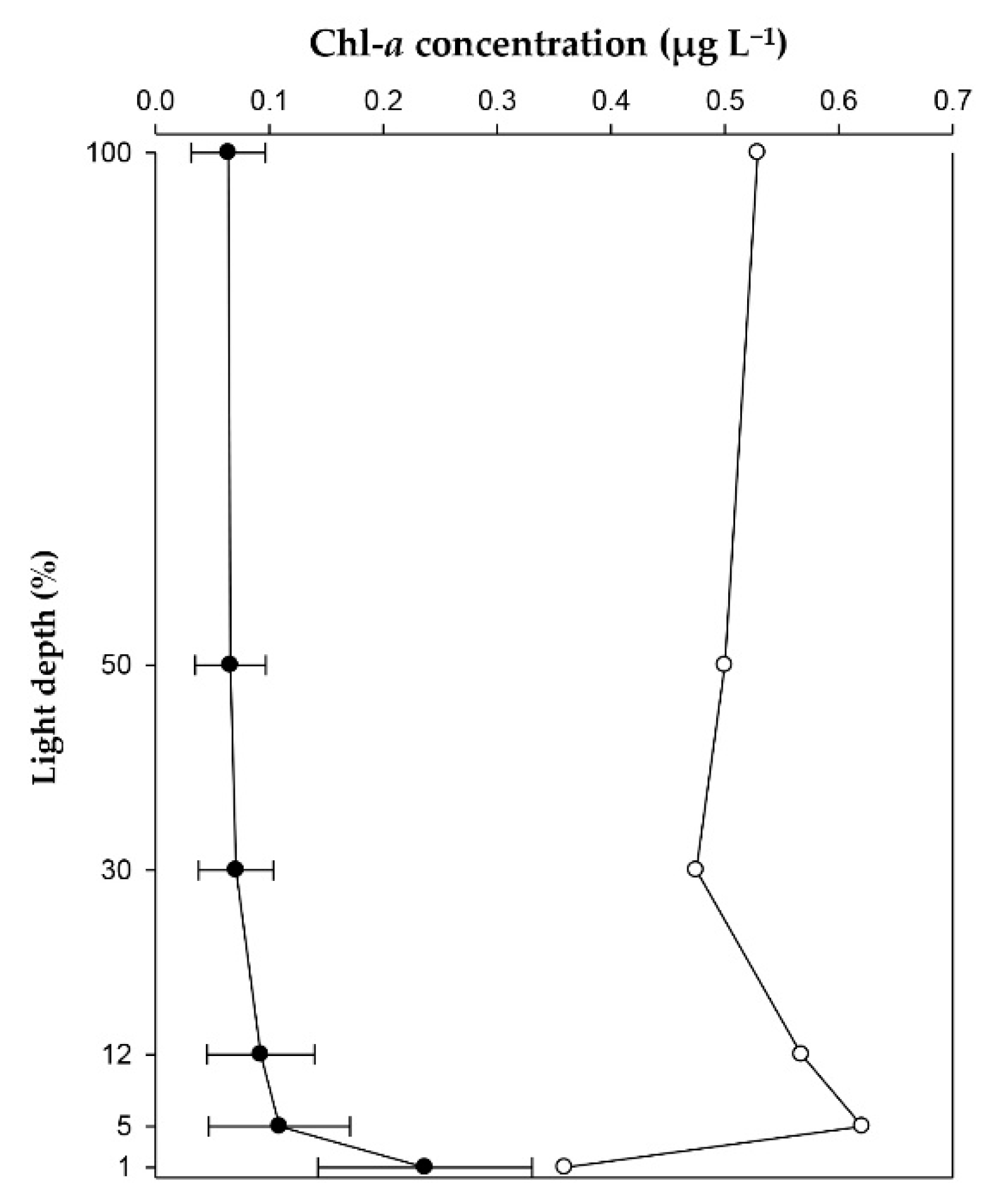
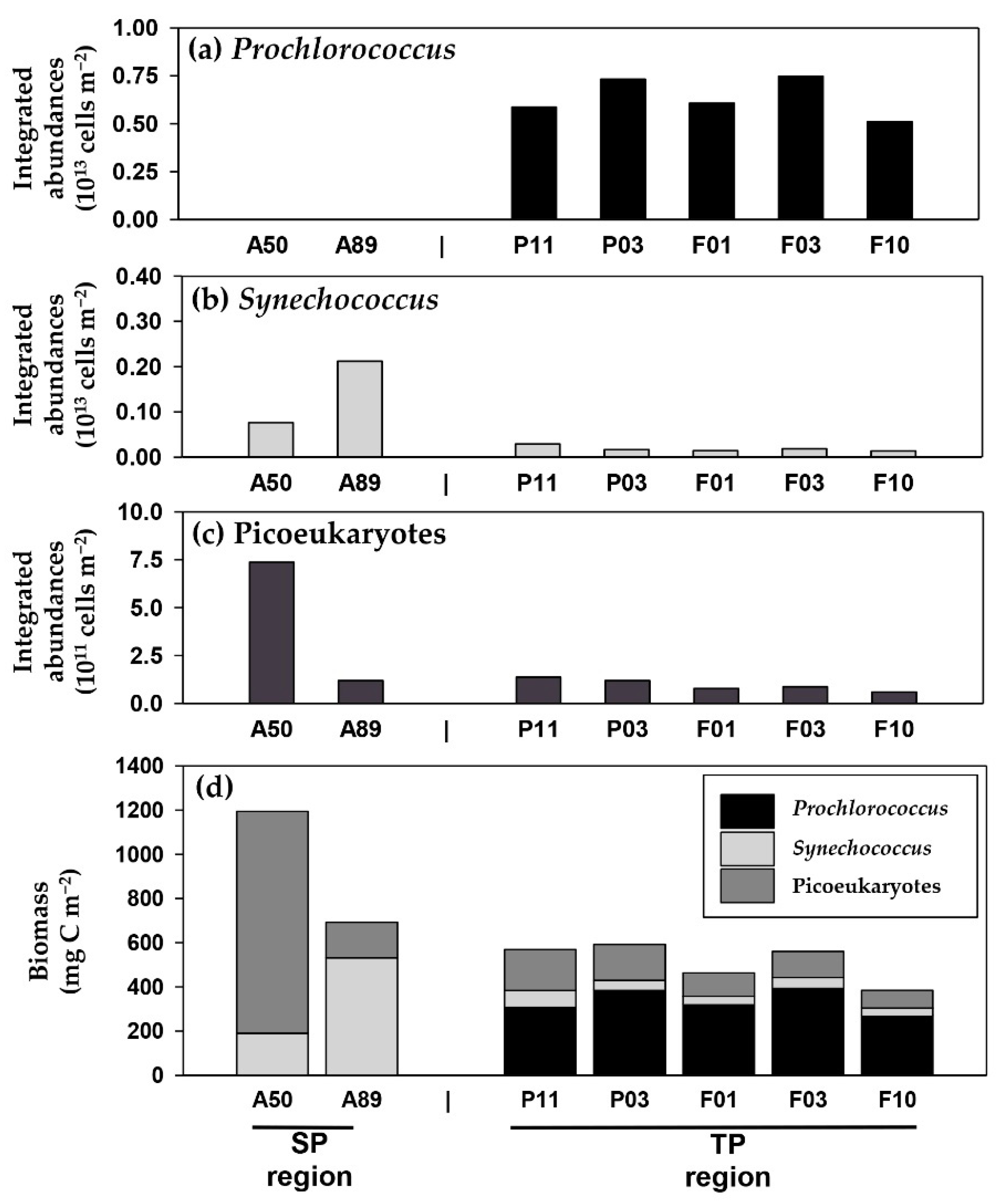
3.2. Distribution of Phytoplankton in Water Column
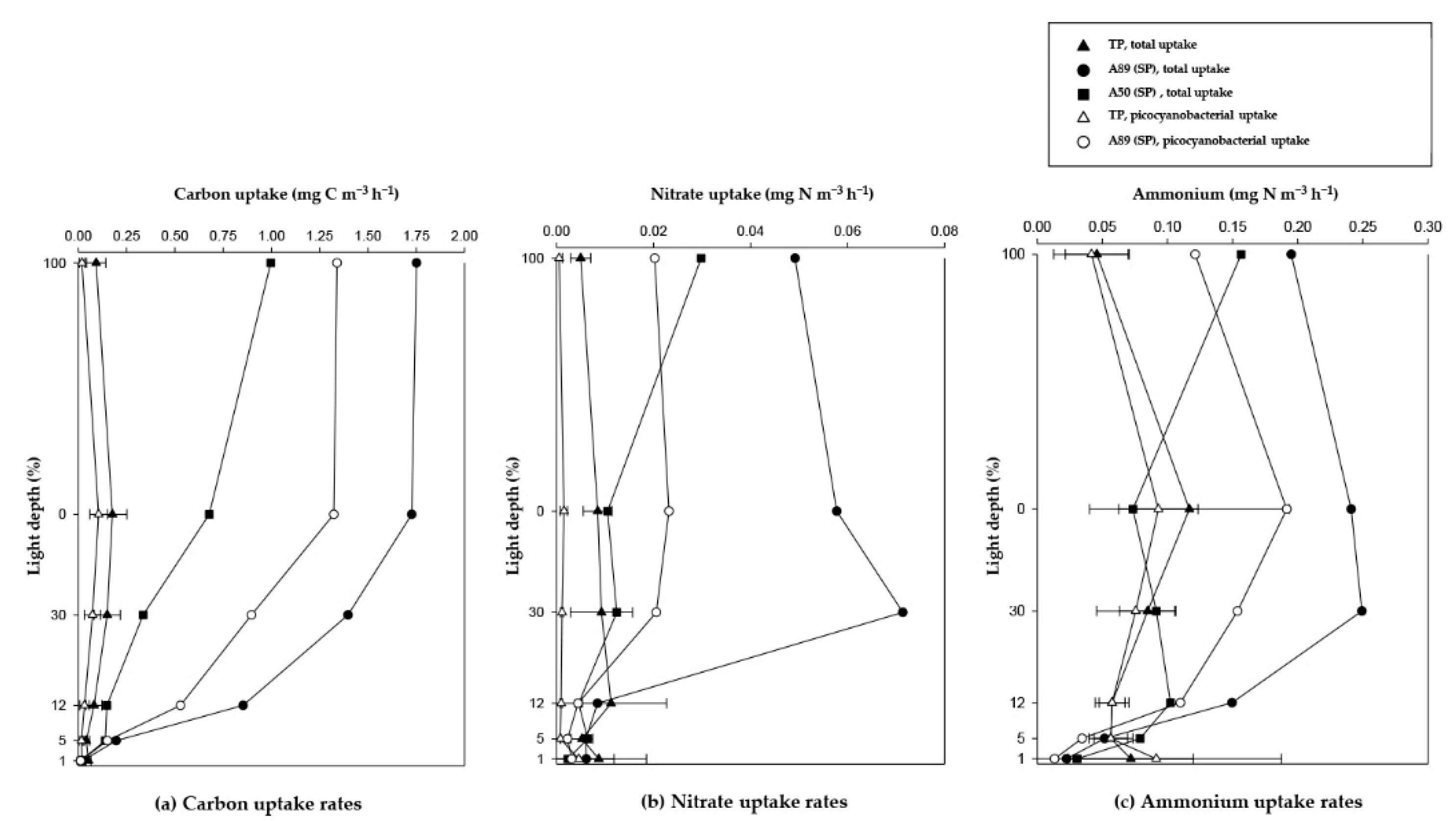
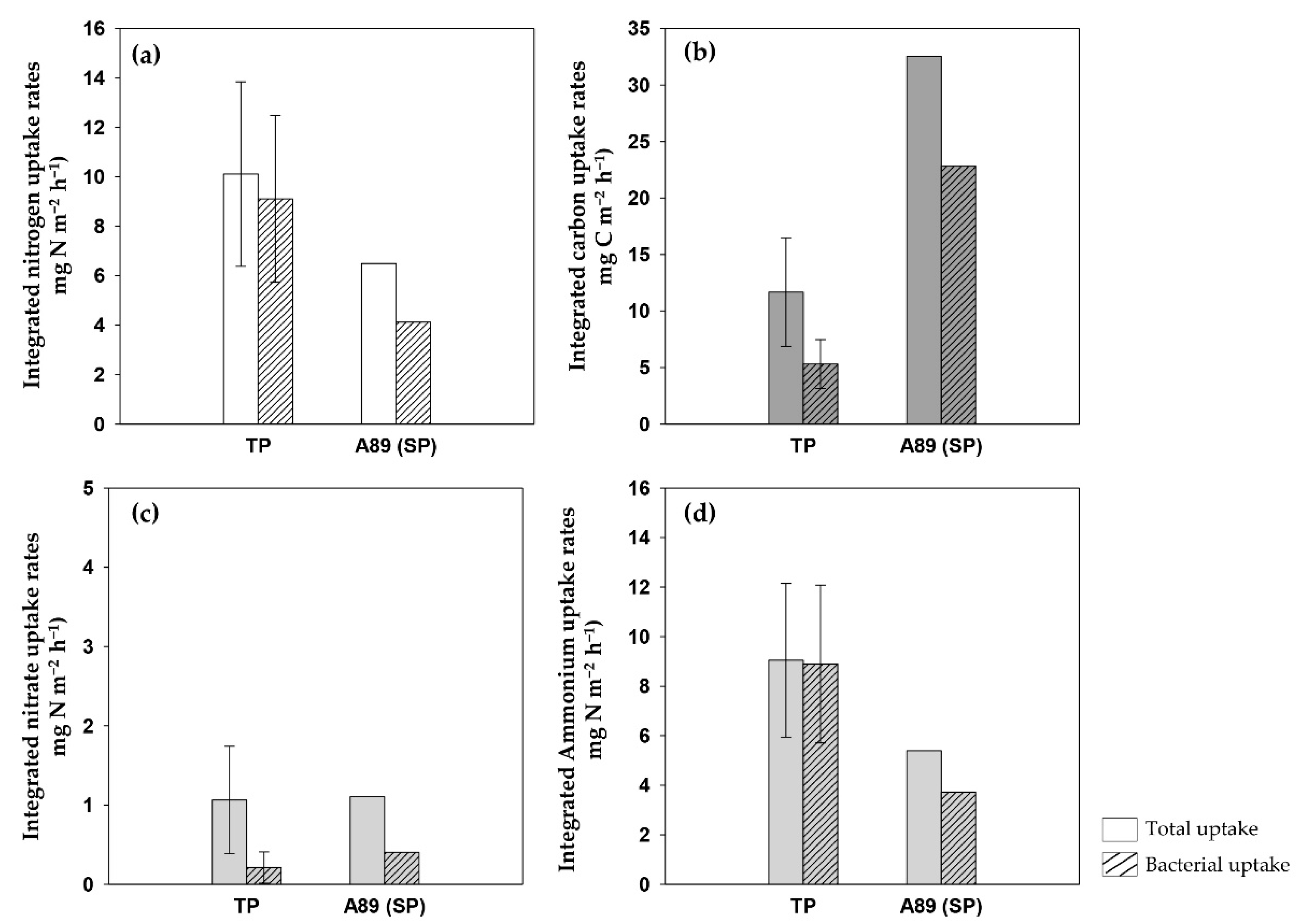
3.3. Total Carbon and Nitrogen Uptake Rates in the NPO
3.4. Picocyanobacterial Carbon and Nitrogen Uptakes in the NPO
4. Discussion
5. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Behrenfeld, M.J.; Randerson, J.T.; McClain, C.R.; Feldman, G.C.; Los, S.O.; Tucker, C.J.; Falkowski, P.G.; Field, C.B.; Frouin, R.; Esaias, W.E.; et al. Biospheric primary production during an ENSO transition. Science 2001, 291, 2594–2597. [Google Scholar] [CrossRef] [Green Version]
- Howarth, R.W. Nutrient Limitation of Net Primary Production in Marine Ecosystems. Annu. Rev. Ecol. Syst. 1988, 19, 89–110. [Google Scholar] [CrossRef]
- Morán, X.A.G.; López-Urrutia, Á.; Calvo-Díaz, A.; LI, W.K.W. Increasing importance of small phytoplankton in a warmer ocean. J. Glob. Chang. Biol. 2010, 16, 1137–1144. [Google Scholar] [CrossRef]
- Taniguchi, A. Geographical variation of primary production in the Western Pacific Ocean and adjacent seas with reference to the inter-relations between various parameters of primary production. Mem. Fac. Fish. Hokkaido Univ. 1972, 19, 1–33. [Google Scholar]
- Kwak, J.H.; Hwang, J.; Choy, E.J.; Park, H.J.; Kang, D.J.; Lee, T.; Chang, K.I.; Kim, K.R.; Kang, C.K. High primary productivity and f-ratio in summer in the Ulleung basin of the East/Japan Sea. Deep-Sea Res. I 2013, 79, 74–785. [Google Scholar] [CrossRef]
- Lee, S.H.; Yun, M.S.; Kim, B.K.; Saitoh, S.; Kang, C.K.; Kang, S.H.; Whitledge, T. Latitudinal carbon productivity in the Bering and Chukchi Seas during the summer in 2007. Cont. Shelf Res. 2013, 59, 28–36. [Google Scholar]
- Lee, S.H.; Yun, M.S.; Kim, B.K.; Joo, H.T.; Kang, S.H.; Kang, C.K.; Whitledge, T.E. Contribution of small phytoplankton to total primary production in the Chukchi Sea. Cont. Shelf Res. 2013, 68, 43–50. [Google Scholar] [CrossRef]
- Kim, B.K.; Joo, H.T.; Song, H.J.; Yang, E.J.; Lee, S.H.; Hahm, D.; Rhee, T.S.; Lee, S.H. Large seasonal variation in phytoplankton production in the Amundsen Sea. Polar Biol. 2015, 38, 319–331. [Google Scholar] [CrossRef]
- Fuhrman, J.A.; Hagström, Å. Bacterial and Archaeal Community Structure and its Patterns. In Microbial Ecology of the Oceans, 2nd ed.; Kirchman, D.L., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2008; pp. 45–90. [Google Scholar]
- Lee, C.R.; Choi, K.H.; Kang, H.K.; Yang, E.J.; Noh, J.H.; Choi, D.H. Biomass and trophic structure of the plankton community in subtropical and temperate waters of the northwestern Pacific Ocean. J. Oceanogr. 2012, 68, 473–482. [Google Scholar] [CrossRef]
- Li, W.K.W.; Rao, D.V.S.; Harrison, W.G.; Smith, J.C.; Cullen, J.J.; Irwin, B.; Platt, T. Autotrophic picoplankton in the tropical ocean. Science 1983, 219, 292–295. [Google Scholar] [CrossRef]
- Glover, H.E.; Campbell, L.; Prezelin, B.B. Contribution of Synechococcus spp. to size-fractioned primary productivity in three water masses in the Northwest Atlantic Ocean. Mar. Biol. 1986, 91, 193–203. [Google Scholar] [CrossRef]
- Vaulot, D. The Cell Cycle of Phytoplankton: Coupling Cell Growth to Population Growth. In Molecular Ecology of Aquatic Microbes; NATO ASI Series (Series G: Ecological Sciences); Joint, I., Ed.; Springer: Berlin/Heidelberg, Germany, 1995. [Google Scholar] [CrossRef]
- Chavez, F.P.; Buck, K.R.; Service, S.K.; Newton, J.; Barber, R.T. Phytoplankton variability in the central and eastern tropical pacific. Deep Sea Res. II 1996, 43, 835–870. [Google Scholar] [CrossRef]
- Liu, H.; Nolla, H.A.; Campbell, L. Prochlorococcus growth rate and contribution to primary production in the equatorial and subtropical North Pacific Ocean. Aquat. Microb. Ecol. 1997, 12, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Choi, D.H.; Selph, K.E.; Noh, J.H. Niche partitioning of picocyanobacterial lineages in the oligotrophic northwestern Pacific Ocean. Algae 2015, 30, 223–232. [Google Scholar] [CrossRef]
- Bhavya, P.S.; Min, J.O.; Kim, M.S.; Jang, H.K.; Kim, K.; Kang, J.J.; Lee, J.H.; Lee, D.; Jo, N.; Kim, M.J.; et al. A Review on Marine N2 Fixation: Mechanism, Evolution of Methodologies, Rates, and Future Concerns. Ocean Sci. J. 2019, 54, 515–528. [Google Scholar] [CrossRef]
- Vaulot, D.; Partensky, F.; Neveux, J.; Mantoura, R.F.C.; Llewellyn, C.A. Winter presence of prochlorophytes in surface waters of the northwestern Mediterranean Sea. Limnol. Oceanogr. 1990, 35, 1156–1164. [Google Scholar] [CrossRef]
- Chisholm, S.W.; Frankel, S.L.; Goericke, R.; Olson, R.J.; Palenik, B.; Waterbury, J.B.; West-Johnsrud, L.; Zettler, E.R. Prochlorococcus marinus nov. gen. nov. sp.: An oxyphototrophic marine prokaryote containing divinyl chlorophyll a and b. Arch. Microbiol. 1992, 157, 297–300. [Google Scholar] [CrossRef]
- Goericke, R.; Welschmeyer, N.A. The marine prochlorophyte Prochlorococcus contributes significantly to phytoplankton biomass and primary production in the Sargasso Sea. Deep-Sea Res. I 1993, 40, 2283–2294. [Google Scholar] [CrossRef]
- Landry, M.R.; Kirshtein, J.; Constantinou, J. Abundances and distributions of picoplankton populations in the central equatorial pacific from 12° N to 12° S, 140° W. Deep-Sea Res. II 1996, 43, 871–890. [Google Scholar] [CrossRef]
- Partensky, F.; Blanchot, J.; Vaulot, D. Differential distribution of Prochlorococcus and Synechococcus in oceanic waters: A review. Ocean. Monaco-Numero Spec. 1999, 19, 457–476. [Google Scholar]
- Zinser, E.R.; Johnson, Z.I.; Coe, A.; Karaca, E.; Veneziano, D.; Chisholm, S.W. Influence of light and temperature on Prochlorococcus ecotype distributions in the Atlantic Ocean. Limnol. Oceanogr. 2007, 52, 2205–2220. [Google Scholar] [CrossRef]
- Choi, D.H.; Noh, J.H.; An, S.M.; Choi, Y.R.; Lee, H.; Ra, K.; Kim, D.; Rho, T.; Lee, S.H.; Kim, K.T.; et al. Spatial distribution of cold-adapted Synechococcus during spring in seas adjacent to Korea. Algae 2016, 31, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Barber, R.T. Picoplankton do some heavy lifting. Science 2007, 315, 777–778. [Google Scholar] [CrossRef]
- Flombaum, P.; Gallegos, J.L.; Gordillo, R.A.; Rincón, J.; Zabala, L.L.; Jiao, N.; Karl, D.M.; Li, W.K.W.; Lomas, M.W.; Veneziano, D.; et al. Present and future global distributions of the marine Cyanobacteria Prochlorococcus and Synechococcus. Proc. Natl. Acad. Sci. USA 2013, 110, 9824–9829. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Joo, H.T.; Lee, J.H.; Lee, J.H.; Kang, J.J.; Lee, H.W.; Lee, D.; Kang, C.K. Seasonal carbon uptake rates of phytoplankton in the northern East/Japan Sea. Deep-Sea Res. II 2017, 143, 45–53. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, B.K.; Lim, Y.J.; Joo, H.T.; Kang, J.J.; Lee, D.; Park, J.; Ha, S.Y.; Lee, S.H. Small phytoplankton contribution to the standing stocks and the total primary production in the Amundsen Sea. Biogeosciences 2017, 14, 3705–3713. [Google Scholar] [CrossRef] [Green Version]
- Jang, H.K.; Kang, J.J.; Lee, J.H.; Kim, M.; Ahn, S.H.; Jeong, J.Y.; Yun, M.S.; Han, I.S.; Lee, S.H. Recent Primary Production and Small Phytoplankton Contribution in the Yellow Sea during the Summer in 2016. Ocean Sci. J. 2018, 53, 509–519. [Google Scholar] [CrossRef]
- Kang, J.J.; Jang, H.K.; Lim, J.H.; Lee, D.; Lee, J.H.; Bae, H.; Lee, C.H.; Kang, C.K.; Lee, S.H. Characteristics of Different Size Phytoplankton for Primary Production and Biochemical Compositions in the Western East/Japan Sea. Front. Microbiol. 2020, 11, 1–16. [Google Scholar] [CrossRef]
- Sarma, V.V.S.S.; Chopra, M.; Rao, D.N.; Priya, M.M.R.; Rajula, G.R.; Lakshmi, D.S.R.; Rao, V.D. Role of eddies on controlling total and size-fractionated primary production in the Bay of Bengal. Cont. Shelf Res. 2020, 204, 104186. [Google Scholar] [CrossRef]
- Fouilland, E.; Gosselin, M.; Rivkin, R.B.; Vasseur, C.; Mostajir, B. Nitrogen uptake by heterotrophic bacteria and phytoplankton in Arctic surface waters. J. Plankton Res. 2007, 29, 369–376. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, P.A.; Kirchman, D.L. Utilization of inorganic and organic nitrogen by bacteria in marine systems. Limnol. Oceanogr. 1986, 31, 998–1009. [Google Scholar] [CrossRef] [Green Version]
- Middelburg, J.J.; Nieuwenhuize, J. Nitrogen uptake by heterotrophic bacteria and phytoplankton in the nitrate-rich Thames estuary. Mar. Ecol. Prog. Ser. 2000, 203, 13–21. [Google Scholar] [CrossRef]
- Middelburg, J.J.; Nieuwenhuize, J. Uptake of dissolved inorganic nitrogen in turbid, tidal estuaries. Mar. Ecol. Prog. Ser. 2000, 192, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Vazquez, D. Inhibitors of protein synthesis. FEBS Lett. 1974, 40, S63–S84. [Google Scholar] [CrossRef] [Green Version]
- Jacoby, G.A.; Gorini, L. The Effect of Streptomycin and Other Aminoglycoside Antibiotics on Protein Synthesis. In Antibiotics; Gottlieb, D., Shaw, P.D., Eds.; Springer: Berlin/Heidelberg, Germany, 1967. [Google Scholar] [CrossRef]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A Manual of Chemical and Biological Methods for Seawater Analysis; Pergamon Press: New York, NY, USA, 1984. [Google Scholar]
- Olson, R.J.; Shalapyonok, A.; Sosik, H.M. An automated submersible flow cytometer for analyzing pico- and nanophytoplankton: FlowCytobot. Deep-Sea Res. I 2003, 50, 301–315. [Google Scholar] [CrossRef]
- Lee, S.H.; Whitledge, T.E.; Kang, S.H. Recent carbon and nitrogen uptake rates of phytoplankton in Bering Strait and the Chukchi Sea. Cont. Shelf Res. 2007, 27, 2231–2249. [Google Scholar] [CrossRef]
- Song, H.J.; Kim, K.; Lee, J.H.; Ahn, S.H.; Joo, H.M.; Jeong, J.Y.; Yang, E.J.; Kang, S.H.; Yun, M.S.; Lee, S.H. In-situ Measured Carbon and Nitrogen Uptake Rates of Melt Pond Algae in the Western Arctic Ocean, 2014. Ocean Sci. J. 2018, 53, 107–117. [Google Scholar] [CrossRef]
- Lim, Y.J.; Kim, T.W.; Lee, S.H.; Lee, D.; Park, J.; Kim, B.K.; Kim, K.; Jang, H.K.; Bhavya, P.S.; Lee, S.H. Seasonal Variations in the Small Phytoplankton Contribution to the Total Primary Production in the Amundsen Sea, Antarctica. J. Geophys. Res. Oceans. 2019, 124, 8324–8341. [Google Scholar] [CrossRef]
- Yun, M.S.; Kim, Y.; Jeong, Y.; Joo, H.T.; Jo, Y.H.; Lee, C.H.; Bae, H.; Lee, D.; Bhavya, P.S.; Kim, D.; et al. Weak Response of Biological Productivity and Community Structure of Phytoplankton to Mesoscale Eddies in the Oligotrophic Philippine Sea. J. Geophys. Res. Oceans 2020, 125, e2020JC016436. [Google Scholar] [CrossRef]
- Hama, T.; Miyazaki, T.; Ogawa, Y.; Iwakuma, T.; Takahashi, M.; Otsuki, A.; Ichimura, S. Measurement of photosynthetic production of a marine phytoplankton population using a stable 13C isotope. Mar. Biol. 1983, 73, 31–36. [Google Scholar] [CrossRef]
- Dugdale, R.C.; Goering, J.J. Uptake of New and Regenerated Forms of Nitrogen in Primary Productivity. Limnol. Oceanogr. 1967, 12, 196–206. [Google Scholar] [CrossRef] [Green Version]
- Gosselin, M.; Levasseur, M.; Wheeler, P.A.; Horner, R.A.; Booth, B.C. New measurements of phytoplankton and ice algal production in the Arctic Ocean. Deep Sea Res. Part II Top. Stud. Ocean. 1997, 44, 1623–1644. [Google Scholar] [CrossRef]
- Furuya, K.; Kurita, K.; Odate, T. Distribution of phytoplankton in the East China sea in the winter of 1993. J. Oceanogr. 1996, 52, 323–333. [Google Scholar] [CrossRef]
- Gibb, S.W.; Barlow, R.G.; Cummings, D.G.; Rees, N.W.; Trees, C.C.; Holligan, P.; Suggett, D. Surface phytoplankton pigment distributions in the Atlantic Ocean: An assessment of basin scale variability between 50° N and 50° S. Prog. Oceanogr. 2000, 45, 339–368. [Google Scholar] [CrossRef]
- Platt, T.; Bouman, H.; Devred, E.; Fuentes-Yaco, C.; Sathyendranath, S. Physical forcing and phytoplankton distributions. Sci. Mar. 2005, 69, 55–73. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Huang, B.; Huang, Q.; Wang, L.; Ni, X.; Tang, Q.; Sun, S.; Wei, H.; Liu, S.; Li, C.; et al. Seasonal phytoplankton response to physical processes in the southern Yellow Sea. J. Sea Res. 2015, 95, 45–55. [Google Scholar] [CrossRef]
- Gong, G.C.; Wen, Y.H.; Wang, B.W.; Liu, G.J. Seasonal variation of chlorophyll a concentration, primary production and environmental conditions in the subtropical East China Sea. Deep-Sea Res. II 2003, 50, 1219–1236. [Google Scholar] [CrossRef]
- Moore, L.R.; Chisholm, S.W. Photophysiology of the marine cyanobacterium Prochlorococcus: Ecotypic differences among cultured isolates. Limnol. Oceanogr. 1999, 44, 628–638. [Google Scholar] [CrossRef] [Green Version]
- Moore, L.R.; Post, A.F.; Rocap, G.; Chisholm, S.W. Utilization of different nitrogen sources by the marine cyanobacteria Prochlorococcus and Synechococcus. Limnol. Oceanogr. 2002, 47, 989–996. [Google Scholar] [CrossRef]
- Zhao, S.; Wei, J.; Yue, H.; Xiao, T. Picophytoplankton abundance and community structure in the Philippine Sea, western Pacific. Chin. J. Oceanol. Limnol. 2010, 28, 88–95. [Google Scholar] [CrossRef]
- Choi, D.H.; Noh, J.H.; Hahm, M.S.; Lee, C.M. Picocyanobacterial abundances and diversity in surface water of the northwestern Pacific Ocean. Ocean Sci. J. 2011, 46, 265–271. [Google Scholar] [CrossRef]
- Charpy, L.; Blanchot, J. Photosynthetic picoplankton in French Polynesian atoll lagoons: Estimation of taxa contribution to biomass and production by flow cytometry. Mar. Ecol. Prog. Ser. 1998, 162, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Blanchot, J.; André, J.M.; Navarette, C.; Neveux, J.; Radenac, M.H. Picophytoplankton in the equatorial pacific: Vertical distributions in the warm pool and in the high nutrient low chlorophyll conditions. Deep-Sea Res. I 2001, 48, 297–314. [Google Scholar] [CrossRef]
- McCarthy, J.J.; Garside, C.; Nevins, J.L.; Barber, R.T. New production along 140 degrees W in the equatorial Pacific during and following the 1992 El Niño event. Deep-Sea Res. II 1996, 43, 1065–1093. [Google Scholar] [CrossRef]
- Eppley, R.W.; Peterson, B.J. Particulate organic matter flux and planktonic new production in the deep ocean. Nature 1979, 282, 677–680. [Google Scholar] [CrossRef]
- Kirchman, D.L. The uptake of inorganic nutrients by heterotrophic bacteria. Microb. Ecol. 1994, 28, 225–271. [Google Scholar] [CrossRef]
- Rodrigues, R.M.N.V.; Williams, P.J.L.B. Inorganic nitrogen assimilation by picoplankton and whole plankton in a coastal ecosystem. Limnol. Oceanogr. 2002, 47, 1608–1616. [Google Scholar] [CrossRef]
- Bhavya, P.S.; Kumar, S.; Gupta, G.V.M.; Sudheesh, V.; Sudharma, K.V.; Varrier, D.S.; Dhanya, K.R.; Saravanane, N. Nitrogen Uptake Dynamics in a Tropical Eutrophic Estuary (Cochin, India) and Adjacent Coastal Waters. Estuaries Coasts 2016, 39, 54–67. [Google Scholar] [CrossRef]
- Waterbury, J.B.; Watson, S.W.; Guillard, R.R.; Brand, L.E. Widespread occurrence of a unicellular, marine, planktonic, cyanobacterium. Nature 1979, 227, 293–294. [Google Scholar] [CrossRef]
- Chisholm, S.W.; Olson, R.J.; Zettler, E.R.; Goericke, R.; Waterbury, J.B.; Welschmeyer, N.A. A novel free-living prochlorophyte occurs at high cell concentrations in the oceanic euphotic zone. Nature 1988, 334, 340–343. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TP | SP | |||
|---|---|---|---|---|
| Mean | S.D. | Mean | S.D. | |
| Temperature in the surface (°C) | 29.1 | 0.9 | 17.3 | - |
| Temperature in the euphotic depth (°C) | 26.7 | 2.3 | 15.3 | 2.3 |
| Salinity in the surface (psu) | 34.8 | 0.5 | 33.2 | - |
| Salinity in the euphotic zone (psu) | 34.8 | 0.6 | 33.9 | 0.6 |
| Nitrate in the euphotic zone (µM) | 0.13 | 0.35 | 0.84 | 1.8 |
| Ammonium in the euphotic depth (µM) | 0.14 | 0.07 | 0.18 | 0.03 |
| Euphotic depth (m) | 127.4 | 16.5 | 35 | - |
| TP | SP | |||||
|---|---|---|---|---|---|---|
| Mean | S.D. | n | Mean | S.D. | n | |
| Integrated total Chlorophyll-a (mg Chl-a m−2) | 15 | 6.6 | 7 | 18.1 | - | 2 |
| C/N ratio (atom/atom) | 11 | 1.8 | 7 | 9.8 | - | 2 |
| Carbon specific uptake (h−1) | 0.001508 | 0.001034 | 42 | 0.004951 | 0.004069 | 12 |
| Carbon absolute uptake (mg C m−3·h−1) | 0.099 | 0.068 | 42 | 0.688 | 0.653 | 12 |
| Integrated carbon uptake (mg C m−2·h−1) | 11.66 | 4.8 | 7 | 20.85 | - | 2 |
| Nitrate specific uptake (h−1) | 0.000632 | 0.000435 | 42 | 0.001097 | 0.001096 | 12 |
| Nitrate absolute uptake (mg NO3− m−3·h−1) | 0.007987 | 0.006853 | 42 | 0.022084 | 0.024058 | 12 |
| Integrated nitrate uptake (mg NO3- m−2·h−1) | 1.06 | 0.68 | 7 | 0.69 | - | 2 |
| Ammonium specific uptake (h−1) | 0.006756 | 0.003664 | 42 | 0.006355 | 0.003179 | 12 |
| Ammonium absolute uptake (mg NH4+ m−3·h−1) | 0.072252 | 0.044304 | 42 | 0.120235 | 0.077651 | 12 |
| Integrated ammonium uptake (mg NH4+ m−2·h−1) | 9.05 | 3.1 | 7 | 4.05 | - | 2 |
| Nitrogen specific uptake (h−1) | 0.007388 | 0.004099 | 42 | 0.007452 | 0.004255 | 12 |
| Nitrogen absolute uptake (mg N m−2·h−1) | 0.08 | 0.047 | 42 | 0.142 | 0.1 | 12 |
| Integrated nitrogen uptake (mg N m−2·h−1) | 10.11 | 2.49 | 7 | 4.74 | - | 2 |
| f-ratio | 0.1 | 0.03 | 7 | 0.13 | - | 2 |
| TP | SP (A89) | |||||
|---|---|---|---|---|---|---|
| Mean | SD | n | Mean | SD | n | |
| Picocyanobacterial carbon specific uptake (h−1) | 0.000695 | 0.000548 | 42 | 0.004404 | 0.003065 | 6 |
| Picocyanobacterial carbon absolute uptake (mg C m−3·h−1) | 0.044 | 0.043 | 42 | 0.708 | 0.573 | 6 |
| Integrated picocyanobacterial carbon uptake (mg C m−2·h−1) | 5.31 | 2.16 | 7 | 22.83 | - | 1 |
| Picocyanobacterial nitrate specific uptake (h−1) | 0.000104 | 0.000174 | 42 | 0.00047 | 0.000348 | 6 |
| Picocyanobacterial nitrate absolute uptake (mg NO3− m−3·h−1) | 0.001577 | 0.003138 | 42 | 0.012296 | 0.00997 | 6 |
| Integrated picocyanobacterial nitrate uptake (mg NO3− m−2·h−1) | 0.21 | 0.2 | 7 | 0.4 | - | 1 |
| Picocyanobacterial ammonium specific uptake (h−1) | 0.005073 | 0.002693 | 42 | 0.004497 | 0.002518 | 6 |
| Picocyanobacterial ammonium absolute uptake (mg NH4+ m−3·h−1) | 0.070531 | 0.046278 | 42 | 0.104054 | 0.068611 | 6 |
| Integrated picocyanobacterial ammonium uptake (mg NH4+ m−2·h−1) | 8.89 | 3.18 | 7 | 3.72 | - | 1 |
| Integrated picocyanobacterial nitrogen uptake (mg N m−2·h−1) | 9.1 | 3.37 | 7 | 4.12 | - | 1 |
| Picocyanobacterial f-ratio | 0.02 | 0.01 | 7 | 0.1 | - | 1 |
| Region | References | Carbon Uptake Rates | Nitrate Uptake Rates | Ammonium Uptake Rates | |
|---|---|---|---|---|---|
| Average ± SD (g C m−2·d−1) | Average ± SD (g N m−2·d−1) | Average ± SD (g N m−2·d−1) | Season | ||
| TP | Taniguchi (1972) | 0.09 | - | - | Winter |
| Kwak et al. (2013) | 0.2 | - | - | Summer | |
| In this study | 0.15 ± 0.06 | 0.01 ± 0.01 | 0.16 ± 0.01 | Late spring | |
| SP | Gong et al. (2003) | 0.31 ± 0.16 | - | - | Early spring |
| 0.52 ± 0.32 | - | - | Summer | ||
| Kwak et al. (2013) | 0.28 | - | - | Summer | |
| In this study | 0.45 (A89) | 0.02 (A89) | 0.10 (A89) | Late spring | |
| 0.13 (A50) | 0.01 (A50) | 0.05 (A50) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-W.; Noh, J.-H.; Choi, D.-H.; Yun, M.; Bhavya, P.S.; Kang, J.-J.; Lee, J.-H.; Kim, K.-W.; Jang, H.-K.; Lee, S.-H. Picocyanobacterial Contribution to the Total Primary Production in the Northwestern Pacific Ocean. Water 2021, 13, 1610. https://doi.org/10.3390/w13111610
Lee H-W, Noh J-H, Choi D-H, Yun M, Bhavya PS, Kang J-J, Lee J-H, Kim K-W, Jang H-K, Lee S-H. Picocyanobacterial Contribution to the Total Primary Production in the Northwestern Pacific Ocean. Water. 2021; 13(11):1610. https://doi.org/10.3390/w13111610
Chicago/Turabian StyleLee, Ho-Won, Jae-Hoon Noh, Dong-Han Choi, Misun Yun, P. S. Bhavya, Jae-Joong Kang, Jae-Hyung Lee, Kwan-Woo Kim, Hyo-Keun Jang, and Sang-Heon Lee. 2021. "Picocyanobacterial Contribution to the Total Primary Production in the Northwestern Pacific Ocean" Water 13, no. 11: 1610. https://doi.org/10.3390/w13111610
APA StyleLee, H. -W., Noh, J. -H., Choi, D. -H., Yun, M., Bhavya, P. S., Kang, J. -J., Lee, J. -H., Kim, K. -W., Jang, H. -K., & Lee, S. -H. (2021). Picocyanobacterial Contribution to the Total Primary Production in the Northwestern Pacific Ocean. Water, 13(11), 1610. https://doi.org/10.3390/w13111610