
Decay Rate of Escherichia coli in a Mountainous Tropical Headwater Wetland

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
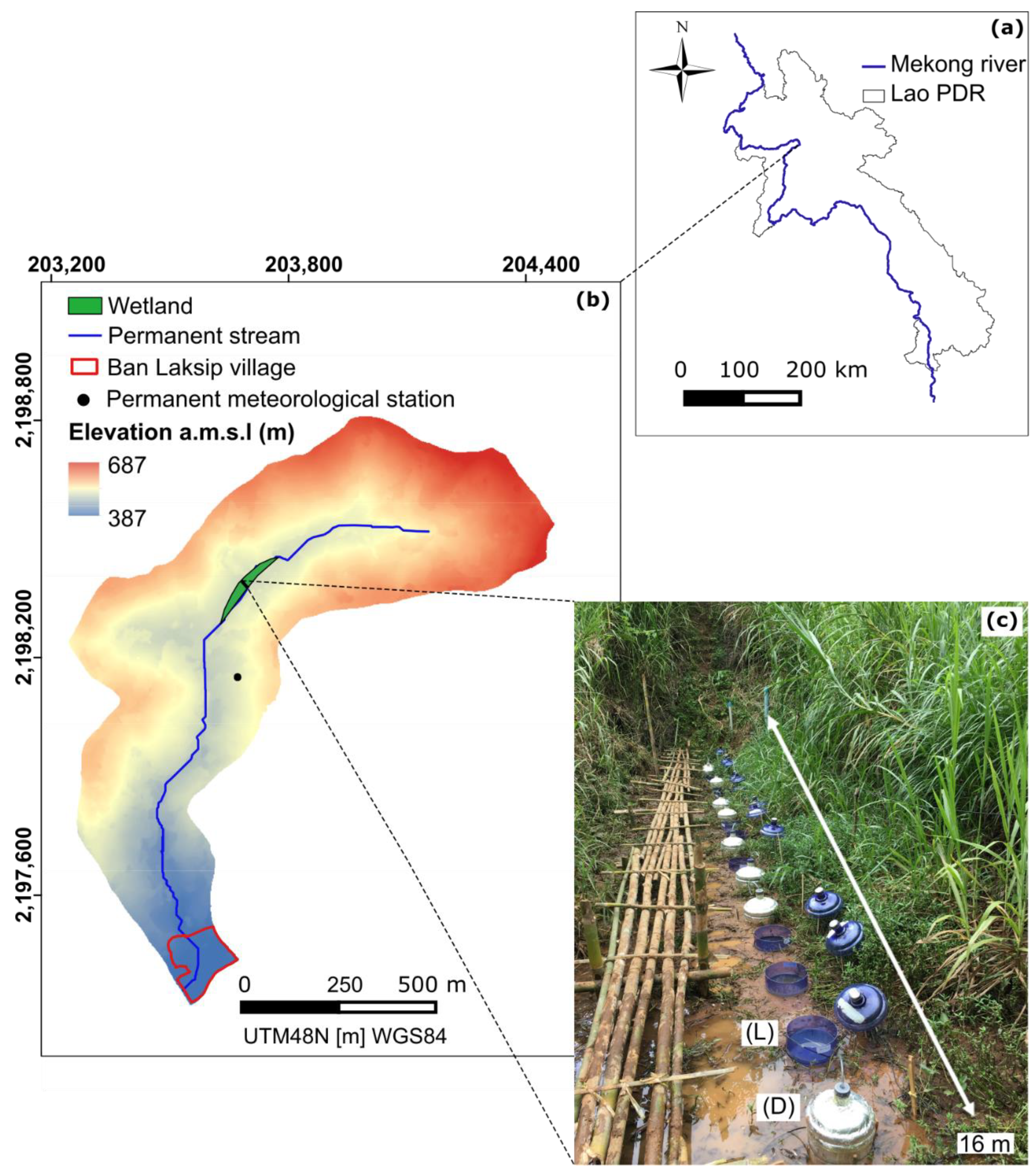
2.1. Study Area
2.2. Experimental Design
- Two of these treatments, i.e., suspended particles deposition in the light (DL) and suspended particles deposition in the dark (DD), were designed to quantify the decay rate of E. coli under wetland-like hydrodynamic conditions enabling the fall of suspended particles (i.e., absence or very low turbulence in the water column), with and without daylight (DL and DD respectively), to simulate the strong attenuation of the transmitted luminous flux to the water surface by dense vegetation cover (Figure 2).
- Two other treatments, i.e., sediment resuspension in the light (RL) and sediment resuspension in the dark (RD), were aimed to evaluate the possible survival or even growth of E. coli in the deposited sediment. To this end, bacteria that had been deposited at the bottom of the mesocosm with the solid particles were resuspended by a daily 1-min manual stirring of the water column using sterile gloves, preceding each sampling. Here the cases with and without daylight (RL and RD respectively) were also tested.
2.3. Mesocosms Preparation
2.4. Analytical Methods: Concentrations of E. coli, TSS, DOC, and Turbidity
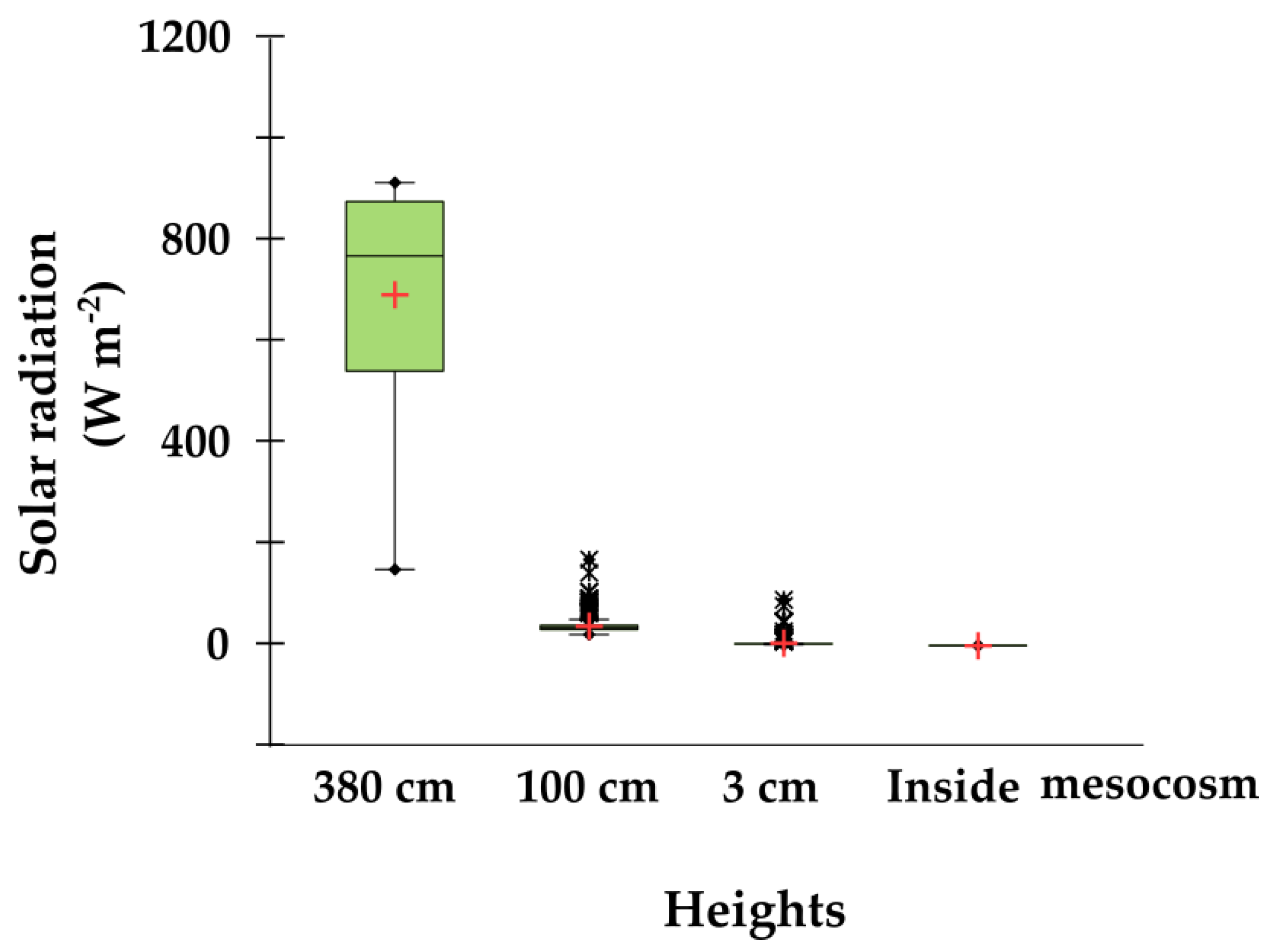
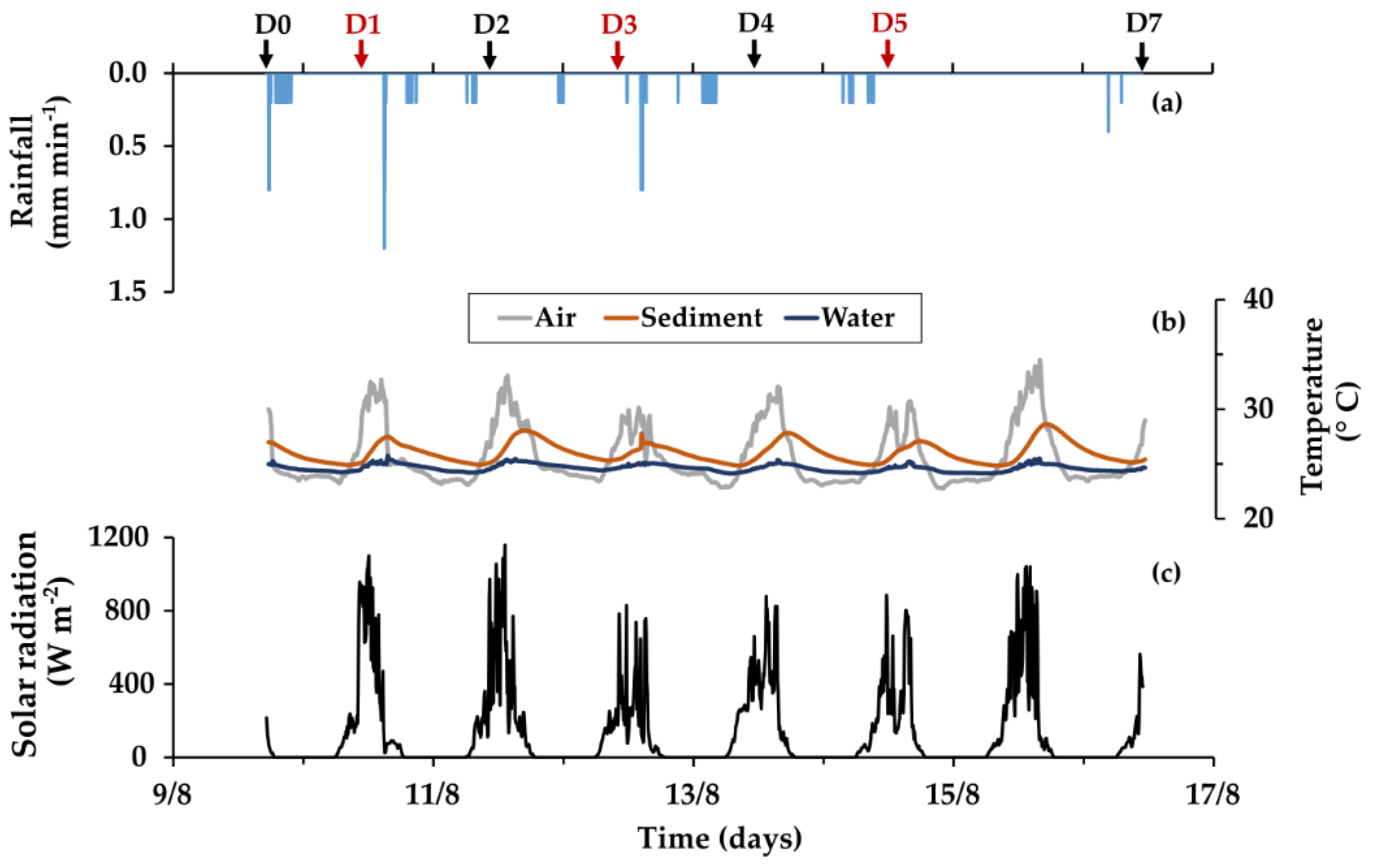
2.5. Environmental Variables: Rainfall, Temperatures and Global Solar Radiation
2.6. Apparent Decay Rates, T50 and T90 Values
2.7. E. coli Stock Variations
3. Results
3.1. Environmental Variables
3.2. Physico-Chemical and Microbiological Variables
3.3. Apparent Decay Rates and T50 and T90 Values
3.4. E. coli Stock Variations
4. Discussion
4.1. Particle Attachment Effect on E. coli Apparent Decay Rates
4.2. Deposition Effect on E. coli Apparent Decay Rates
4.3. Solar Radiation Effect on E. coli Apparent Decay Rates
4.4. Relative Effects of Solar Radiation and Deposition on E. coli Apparent Decay Rates
5. Conclusions
- -
- Particle-attached bacteria prevailed in all mesocosms over the incubation period: over 91% of total E. coli were attached to particles.
- -
- Apparent decay rates of free-living bacteria were higher than apparent decay rates of particle-attached bacteria in all mesocosms.
- -
- Apparent decay rates of total E. coli concentration in mesocosms left to deposit ranged from 1.43 ± 0.15 to 1.17 ± 0.13 day−1 when exposed to light and dark conditions, respectively. Apparent decay rates in mesocosms stirred prior to sampling ranged from 0.50 ± 0.15 to −0.14 ± 0.37 day−1 when exposed to light and dark conditions, respectively.
- -
- Deposition accounted for an average of 92% of the estimated E. coli stock reduction in the water column, while 2% of the estimated E. coli stock reduction was due to the exposure to solar radiation.
- -
- By resuspending the bottom sediment prior to sample collecting in RD mesocosms, we mobilized the deposited E. coli that showed stable concentration during the first 5 days of the experiment, suggesting a survival of bacterial population in bed sediments.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Inheriting a Sustainable World? Atlas on Children’s Health and the Environment; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- UNEPA. A Snapshot of the World’s Water Quality: Towards a Global Assessment; United Nations Environment Programme: Nairobi, Kenya, 2016. [Google Scholar]
- Prüss-Ustün, A.; Wolf, J.; Bartram, J.; Clasen, T.; Cumming, O.; Freeman, M.C.; Gordon, B.; Hunter, P.R.; Medlicott, K.; Johnston, R. Burden of Disease from Inadequate Water, Sanitation and Hygiene for Selected Adverse Health Outcomes: An Updated Analysis with a Focus on Low- and Middle-Income Countries. Int. J. Hyg. Environ. Health 2019, 222, 765–777. [Google Scholar] [CrossRef] [PubMed]
- WHO/UNICEF Joint Water Supply and Sanitation Monitoring Programme and World Health Organization. Progress on Sanitation and Drinking Water: 2015 Update and MDG Assessment; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Ribolzi, O.; Cuny, J.; Sengsoulichanh, P.; Mousquès, C.; Soulileuth, B.; Pierret, A.; Huon, S.; Sengtaheuanghoung, O. Land Use and Water Quality along a Mekong Tributary in Northern Lao P.D.R. Environ. Manag. 2011, 47, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Ribolzi, O.; Evrard, O.; Huon, S.; Rochelle-Newall, E.; Henri-des-Tureaux, T.; Silvera, N.; Thammahacksac, C.; Sengtaheuanghoung, O. Use of Fallout Radionuclides (7Be, 210Pb) to Estimate Resuspension of Escherichia coli from Streambed Sediments during Floods in a Tropical Montane Catchment. Environ. Sci. Pollut. Res. 2016, 23, 3427–3435. [Google Scholar] [CrossRef] [PubMed]
- Boithias, L.; Choisy, M.; Souliyaseng, N.; Jourdren, M.; Quet, F.; Buisson, Y.; Thammahacksa, C.; Silvera, N.; Latsachack, K.; Sengtaheuanghoung, O.; et al. Hydrological Regime and Water Shortage as Drivers of the Seasonal Incidence of Diarrheal Diseases in a Tropical Montane Environment. PLoS Negl. Trop. Dis. 2016, 10, e0005195. [Google Scholar] [CrossRef]
- Johnston, C.A. Sediment and Nutrient Retention by Freshwater Wetlands: Effects on Surface Water Quality. Crit. Rev. Environ. Control 1991, 21, 491–565. [Google Scholar] [CrossRef]
- Mander, Ü.; Mitsch, W.J. Pollution Control by Wetlands. Ecol. Eng. 2009, 35, 153–158. [Google Scholar] [CrossRef]
- Arias, M.E.; Holtgrieve, G.W.; Ngor, P.B.; Dang, T.D.; Piman, T. Maintaining Perspective of Ongoing Environmental Change in the Mekong Floodplains. Curr. Opin. Environ. Sustain. 2019, 37, 1–7. [Google Scholar] [CrossRef]
- MacAlister, C.; Mahaxay, M. Mapping Wetlands in the Lower Mekong Basin for Wetland Resource and Conservation Management Using Landsat ETM Images and Field Survey Data. J. Environ. Manag. 2009, 90, 2130–2137. [Google Scholar] [CrossRef]
- Berg, H.; Ekman Söderholm, A.; Söderström, A.S.; Tam, N.T. Recognizing Wetland Ecosystem Services for Sustainable Rice Farming in the Mekong Delta, Vietnam. Sustain. Sci. 2017, 12, 137–154. [Google Scholar] [CrossRef] [Green Version]
- Grundy-Warr, C.; Andrews, R.H.; Sithithaworn, P.; Petney, T.N.; Sripa, B.; Laithavewat, L.; Ziegler, A.D. Raw Attitudes, Wetland Cultures, Life-Cycles: Socio-Cultural Dynamics Relating to Opisthorchis Viverrini in the Mekong Basin. Parasitol. Int. 2012, 61, 65–70. [Google Scholar] [CrossRef]
- Zedler, J.B.; Kercher, S. WETLAND RESOURCES: Status, Trends, Ecosystem Services, and Restorability. Annu. Rev. Environ. Resour. 2005, 30, 39–74. [Google Scholar] [CrossRef] [Green Version]
- Millennium Ecosystem Assessment. Ecosystems and Human Well-Being: Synthesis; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- Brauman, K.A.; Daily, G.C.; Duarte, T.K.E.O.; Mooney, H.A. The Nature and Value of Ecosystem Services: An Overview Highlighting Hydrologic Services. Annu. Rev. Environ. Resour. 2007, 32, 67–98. [Google Scholar] [CrossRef]
- USEPA (U.S. Environmental Protection Agency). Connectivity of Streams and Wetlands to Downstream Waters: A Review and Synthesis of the Scientific Evidence; US Environmental Protection Agency: Washington, DC, USA, 2015. [Google Scholar]
- Lane, C.R.; Leibowitz, S.G.; Autrey, B.C.; LeDuc, S.D.; Alexander, L.C. Hydrological, Physical, and Chemical Functions and Connectivity of Non-Floodplain Wetlands to Downstream Waters: A Review. J. Am. Water Resour. Assoc. 2018, 54, 346–371. [Google Scholar] [CrossRef]
- Krecek, J.; Haigh, M. Environmental Role of Wetlands in Headwaters; Springer Science & Business Media: Dordrecht, The Netherlands, 2006; Volume 63. [Google Scholar]
- Alexander, R.B.; Boyer, E.W.; Smith, R.A.; Schwarz, G.E.; Moore, R.B. The Role of Headwater Streams in Downstream Water Quality1. JAWRA J. Am. Water Resour. Assoc. 2007, 43, 41–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colvin, S.A.R.; Sullivan, S.M.P.; Shirey, P.D.; Colvin, R.W.; Winemiller, K.O.; Hughes, R.M.; Fausch, K.D.; Infante, D.M.; Olden, J.D.; Bestgen, K.R.; et al. Headwater Streams and Wetlands Are Critical for Sustaining Fish, Fisheries, and Ecosystem Services. Fisheries 2019, 44, 73–91. [Google Scholar] [CrossRef]
- Herrera, D.; Ellis, A.; Fisher, B.; Golden, C.D.; Johnson, K.; Mulligan, M.; Pfaff, A.; Treuer, T.; Ricketts, T.H. Upstream Watershed Condition Predicts Rural Children’s Health across 35 Developing Countries. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, S.; Sadowsky, M.J. Escherichia Coli in the Environment: Implications for Water Quality and Human Health. Microbes Environ. 2008, 23, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quin, A.; Jaramillo, F.; Destouni, G. Dissecting the Ecosystem Service of Large-Scale Pollutant Retention: The Role of Wetlands and Other Landscape Features. AMBIO 2015, 44, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Knox, A.K.; Dahlgren, R.A.; Tate, K.W.; Atwill, E.R. Efficacy of Natural Wetlands to Retain Nutrient, Sediment and Microbial Pollutants. J. Environ. Qual. 2008, 37, 1837–1846. [Google Scholar] [CrossRef]
- Pachepsky, Y.A.; Shelton, D.R. Escherichia Coli and Fecal Coliforms in Freshwater and Estuarine Sediments. Crit. Rev. Environ. Sci. Technol. 2011, 41, 1067–1110. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for Drinking-Water Quality, 4th ed.; WHO Chronicle; WHO: Geneva, Switzerland, 2011. [Google Scholar]
- Rochelle-Newall, E.; Nguyen, T.M.H.; Le, T.P.Q.; Sengtaheuanghoung, O.; Ribolzi, O. A Short Review of Fecal Indicator Bacteria in Tropical Aquatic Ecosystems: Knowledge Gaps and Future Directions. Front. Microbiol. 2015, 6, 308. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.L.; Whitlock, J.E.; Harwood, V.J. Persistence and Differential Survival of Fecal Indicator Bacteria in Subtropical Waters and Sediments. Appl. Environ. Microbiol. 2005, 71, 3041–3048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naganandhini, S.; Kennedy, Z.J.; Uyttendaele, M.; Balachandar, D. Persistence of Pathogenic and Non-Pathogenic Escherichia Coli Strains in Various Tropical Agricultural Soils of India. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [Green Version]
- Isobe, K.O.; Tarao, M.; Chiem, N.H.; Minh, L.Y.; Takada, H. Effect of Environmental Factors on the Relationship between Concentrations of Coprostanol and Fecal Indicator Bacteria in Tropical (Mekong Delta) and Temperate (Tokyo) Freshwaters. Appl. Environ. Microbiol. 2004, 70, 814–821. [Google Scholar] [CrossRef] [Green Version]
- Petersen, F.; Hubbart, J.A. Physical Factors Impacting the Survival and Occurrence of Escherichia Coli in Secondary Habitats. Water 2020, 12, 1796. [Google Scholar] [CrossRef]
- Solo-Gabriele, H.M.; Wolfert, M.A.; Desmarais, T.R.; Palmer, C.J. Sources of Escherichia Coli in a Coastal Subtropical Environment. Appl. Environ. Microbiol. 2000, 66, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.M.; Thoe, W.; Lee, J.H.W. Field and Laboratory Studies of Escherichia Coli Decay Rate in Subtropical Coastal Water. J. Hydro-Environ. Res. 2015, 9, 1–14. [Google Scholar] [CrossRef]
- Fujiokal, R.S.; Narikawa, O.T. Effect of Sunlight on Enumeration of Indicator Bacteria Under Field Conditions. Appl. Environ. Microbiol. 1982, 44, 395–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinton, L.W.; Hall, C.H.; Lynch, P.A.; Davies-Colley, R.J. Sunlight Inactivation of Fecal Indicator Bacteria and Bacteriophages from Waste Stabilization Pond Effluent in Fresh and Saline Waters. Appl. Environ. Microbiol. 2002, 68, 1122–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, K.L.; Boehm, A.B.; Davies-Colley, R.J.; Dodd, M.C.; Kohn, T.; Linden, K.G.; Liu, Y.; Maraccini, P.A.; McNeill, K.; Mitch, W.A.; et al. Sunlight-Mediated Inactivation of Health-Relevant Microorganisms in Water: A Review of Mechanisms and Modeling Approaches. Environ. Sci. Process. Impacts 2018, 20, 1089–1122. [Google Scholar] [CrossRef]
- Jozić, S.; Morović, M.; Šolić, M.; Krstulović, N.; Ordulj, M. Effect of Solar Radiation, Temperature and Salinity on the Survival of Two Different Strains of Escherichia Coli. Fresenius Environ. Bull. 2014, 23, 1852–1859. [Google Scholar]
- Pienaar, J.A.; Singh, A.; Barnard, T.G. The Viable but Non-Culturable State in Pathogenic Escherichia Coli: A General Review. Afr. J. Lab. Med. 2016, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitman, R.L.; Nevers, M.B.; Korinek, G.C.; Byappanahalli, M.N. Solar and Temporal Effects on Escherichia Coli Concentration at a Lake Michigan Swimming Beach. Appl. Environ. Microbiol. 2004, 70, 4276–4285. [Google Scholar] [CrossRef] [Green Version]
- Maraccini, P.A.; Mattioli, M.C.M.; Sassoubre, L.M.; Cao, Y.; Griffith, J.F.; Ervin, J.S.; Van De Werfhorst, L.C.; Boehm, A.B. Solar Inactivation of Enterococci and Escherichia Coli in Natural Waters: Effects of Water Absorbance and Depth. Environ. Sci. Technol. 2016, 50, 5068–5076. [Google Scholar] [CrossRef] [PubMed]
- Walters, E.; Graml, M.; Behle, C.; Horn, H.; Müller, E. Influence of Particle Association and Suspended Solids on UV Inactivation of Fecal Indicator Bacteria in an Urban River. Water Air Soil Pollut. 2014, 225, 1822. [Google Scholar] [CrossRef]
- Perkins, T.L.; Perrow, K.; Rajko-Nenow, P.; Jago, C.F.; Jones, D.L.; Malham, S.K.; McDonald, J.E. Decay Rates of Faecal Indicator Bacteria from Sewage and Ovine Faeces in Brackish and Freshwater Microcosms with Contrasting Suspended Particulate Matter Concentrations. Sci. Total Environ. 2016, 572, 1645–1652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karim, M.R.; Manshadi, F.D.; Karpiscak, M.M.; Gerba, C.P. The Persistence and Removal of Enteric Pathogens in Constructed Wetlands. Water Res. 2004, 38, 1831–1837. [Google Scholar] [CrossRef] [PubMed]
- O’Geen, A.T.; Bianchi, M.L. Using Wetlands to Remove Microbial Pollutants from Farm Discharge Water. Agric. Nat. Resour. Publ. 2015, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Howell, J.M.; Coyne, M.S.; Cornelius, P.L. Effect of Sediment Particle Size and Temperature on Fecal Bacteria Mortality Rates and the Fecal Coliform/Fecal Streptococci Ratio. J. Environ. Qual. 1996, 25, 1216–1220. [Google Scholar] [CrossRef] [Green Version]
- Boutilier, L.; Jamieson, R.; Gordon, R.; Lake, C.; Hart, W. Adsorption, Sedimentation, and Inactivation of E. Coli within Wastewater Treatment Wetlands. Water Res. 2009, 43, 4370–4380. [Google Scholar] [CrossRef] [PubMed]
- Boithias, L.; Ribolzi, O.; Lacombe, G.; Thammahacksa, C.; Silvera, N.; Latsachack, K.; Soulileuth, B.; Viguier, M.; Auda, Y.; Robert, E.; et al. Quantifying the Effect of Overland Flow on Escherichia Coli Pulses during Floods: Use of a Tracer-Based Approach in an Erosion-Prone Tropical Catchment. J. Hydrol. 2021, 594, 125935. [Google Scholar] [CrossRef]
- Blaustein, R.A.; Pachepsky, Y.; Hill, R.L.; Shelton, D.R.; Whelan, G. Escherichia Coli Survival in Waters: Temperature Dependence. Water Res. 2013, 47, 569–578. [Google Scholar] [CrossRef]
- Craig, D.L.; Fallowfield, H.J.; Cromar, N.J. Use of Microcosms to Determine Persistence of Escherichia coli in Recreational Coastal Water and Sediment and Validation with in situ Measurements. J. Appl. Microbiol. 2004, 96, 922–930. [Google Scholar] [CrossRef]
- Shelton, D.R.; Pachepsky, Y.A.; Kiefer, L.A.; Blaustein, R.A.; McCarty, G.W.; Dao, T.H. Response of Coliform Populations in Streambed Sediment and Water Column to Changes in Nutrient Concentrations in Water. Water Res. 2014, 59, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Gregory, L.F.; Karthikeyan, R.; Aitkenhead-Peterson, J.A.; Gentry, T.J.; Wagner, K.L.; Harmel, R.D. Nutrient Loading Impacts on Culturable E. Coli and Other Heterotrophic Bacteria Fate in Simulated Stream Mesocosms. Water Res. 2017, 126, 442–449. [Google Scholar] [CrossRef]
- Bordalo, A.A.; Onrassami, R.; Dechsakulwatana, C. Survival of Faecal Indicator Bacteria in Tropical Estuarine Waters (Bangpakong River, Thailand). J. Appl. Microbiol. 2002, 93, 864–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korajkic, A.; Wanjugi, P.; Harwood, V.J. Indigenous Microbiota and Habitat Influence Escherichia Coli Survival More than Sunlight in Simulated Aquatic Environments. Appl. Environ. Microbiol. 2013, 79, 5329–5337. [Google Scholar] [CrossRef] [Green Version]
- Boithias, L.; Auda, Y.; Audry, S.; Bricquet, J.; Chanhphengxay, A.; Chaplot, V.; Rouw, A.; Tureaux, T.H.; Huon, S.; Janeau, J.; et al. The Multiscale TROPIcal CatchmentS Critical Zone Observatory M-TROPICS Dataset II: Land Use, Hydrology and Sediment Production Monitoring in Houay Pano, Northern Lao PDR. Hydrol. Process. 2021, 35, e14126. [Google Scholar] [CrossRef]
- Wang, Y.; Pandey, P.; Zheng, Y.; Atwill, E.R.; Pasternack, G. Particle Attached and Free Floating Pathogens Survival Kinetics under Typical Stream and Thermal Spring Temperature Conditions. AMB Express 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Haller, L.; Amedegnato, E.; Poté, J.; Wildi, W. Influence of Freshwater Sediment Characteristics on Persistence of Fecal Indicator Bacteria. Water Air Soil Pollut. 2009, 203, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Gaillardet, J.; Braud, I.; Hankard, F.; Anquetin, S.; Bour, O.; Dorfliger, N.; de Dreuzy, J.R.; Galle, S.; Galy, C.; Gogo, S.; et al. OZCAR: The French Network of Critical Zone Observatories. Vadose Zone J. 2018, 17, 180067. [Google Scholar] [CrossRef] [Green Version]
- Ribolzi, O.; Lacombe, G.; Pierret, A.; Robain, H.; Sounyafong, P.; de Rouw, A.; Soulileuth, B.; Mouche, E.; Huon, S.; Silvera, N.; et al. Interacting Land Use and Soil Surface Dynamics Control Groundwater Outflow in a Montane Catchment of the Lower Mekong Basin. Agric. Ecosyst. Environ. 2018, 268, 90–102. [Google Scholar] [CrossRef]
- Huon, S.; Evrard, O.; Gourdin, E.; Lefèvre, I.; Bariac, T.; Reyss, J.L.; Henry des Tureaux, T.; Sengtaheuanghoung, O.; Ayrault, S.; Ribolzi, O. Suspended Sediment Source and Propagation during Monsoon Events across Nested Sub-Catchments with Contrasted Land Uses in Laos. J. Hydrol. Reg. Stud. 2017, 9, 69–84. [Google Scholar] [CrossRef]
- Huon, S.; de Rouw, A.; Bonté, P.; Robain, H.; Valentin, C.; Lefèvre, I.; Girardin, C.; Le Troquer, Y.; Podwojewski, P.; Sengtaheuanghoung, O. Long-Term Soil Carbon Loss and Accumulation in a Catchment Following the Conversion of Forest to Arable Land in Northern Laos. Agric. Ecosyst. Environ. 2013, 169, 43–57. [Google Scholar] [CrossRef]
- Hjulstrom, F. Studies of the Morphological Activity of Rivers as Illustrated by the River Fyris, Bulletin. Geol. Inst. Upsalsa 1935, 25, 221–527. [Google Scholar]
- Huon, S.; Ribolzi, O.; Aubry, E.; Soulileuth, B.; Longchamp, M.; Angeli, O.; Sengtaheuanghoung, N. Iron and Manganese Concentration Levels in Watercress Cultivated within the Main Stream of the Houay Pano Catchment, Northern Lao PDR. Lao J. Agric. For. 2008, 17, 113–128. [Google Scholar]
- Nguyen, H.T.M.; Le, Q.T.P.; Garnier, J.; Janeau, J.L.; Rochelle-Newall, E. Seasonal Variability of Faecal Indicator Bacteria Numbers and Die-off Rates in the Red River Basin, North Viet Nam. Sci. Rep. 2016, 6, 21644. [Google Scholar] [CrossRef]
- Lebaron, P.; Henry, A.; Lepeuple, A.S.; Pena, G.; Servais, P. An Operational Method for the Real-Time Monitoring of E. Coli Numbers in Bathing Waters. Mar. Pollut. Bull. 2005, 50, 652–659. [Google Scholar] [CrossRef]
- Chick, H. An Investigation of the Laws of Disinfection. J. Hyg. 1908, 8, 92–158. [Google Scholar] [CrossRef] [Green Version]
- Ribolzi, O.; Andrieux, P.; Valles, V.; Bouzigues, R.; Bariac, T.; Voltz, M. Contribution of Groundwater and Overland Flows to Storm Flow Generation in a Cultivated Mediterranean Catchment. Quantification by Natural Chemical Tracing. J. Hydrol. 2000, 233, 241–257. [Google Scholar] [CrossRef]
- Nakhle, P.; Boithias, L.; Pando-Bahuon, A.; Thammahacksa, C.; Gallion, N.; Sounyafong, P.; Silvera, N.; Latsachack, K.; Soulileuth, B.; Rochelle-Newall, E.; et al. Concentrations of Escherichia Coli, Total Suspended Sediment, and Dissolved Organic Carbon, Turbidity, and Environmental Variables Measured in 2019 in a Headwater Wetland in The Houay Pano Catchment, Northern Lao PDR. Available online: https://dataverse.ird.fr/dataset.xhtml?persistentId=doi:10.23708/E52ZTX (accessed on 29 June 2021).
- Oliver, D.M.; Clegg, C.D.; Heathwaite, A.L.; Haygarth, P.M. Preferential Attachment of Escherichia Coli to Different Particle Size Fractions of an Agricultural Grassland Soil. Water Air Soil Pollut. 2007, 185, 369–375. [Google Scholar] [CrossRef] [Green Version]
- Malham, S.K.; Rajko-Nenow, P.; Howlett, E.; Tuson, K.E.; Perkins, T.L.; Pallett, D.W.; Wang, H.; Jago, C.F.; Jones, D.L.; McDonald, J.E. The Interaction of Human Microbial Pathogens, Particulate Material and Nutrients in Estuarine Environments and Their Impacts on Recreational and Shellfish Waters. Environ. Sci. Process. Impacts 2014, 16, 2145–2155. [Google Scholar] [CrossRef] [Green Version]
- Petersen, F.; Hubbart, J.A. Quantifying Escherichia Coli and Suspended Particulate Matter Concentrations in a Mixed-Land Use Appalachian Watershed. Water 2020, 12, 532. [Google Scholar] [CrossRef] [Green Version]
- Nakhle, P.; Ribolzi, O.; Boithias, L.; Rattanavong, S.; Auda, Y.; Sayavong, S.; Zimmermann, R.; Soulileuth, B.; Pando, A.; Thammahacksa, C.; et al. Effects of Hydrological Regime and Land Use on In-Stream Escherichia Coli Concentration in the Mekong Basin, Lao PDR. Sci. Rep. 2021, 11, 3460. [Google Scholar] [CrossRef] [PubMed]
- Byamukama, D.; Mach, R.L.; Kansiime, F.; Manafi, M.; Farnleitner, A.H. Discrimination Efficacy of Fecal Pollution Detection in Different Aquatic Habitats of a High-Altitude Tropical Country, Using PresumptivE. coliforms, Escherichia Coli, and Clostridium Perfringens Spores. Appl. Environ. Microbiol. 2005, 71, 65–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanjugi, P.; Harwood, V.J. The Influence of Predation and Competition on the Survival of Commensal and Pathogenic Fecal Bacteria in Aquatic Habitats. Environ. Microbiol. 2013, 15, 517–526. [Google Scholar] [CrossRef]
- Davies, C.M.; Bavor, H.J. The Fate of Stormwater-Associated Bacteria in Constructed Wetland and Water Pollution Control Pond Systems. J. Appl. Microbiol. 2000, 89, 349–360. [Google Scholar] [CrossRef] [Green Version]
- Brettar, I.; Hoflet, M.G. Influence of Ecosystematic Factors on Survival of Escherichia Coli after Large-Scale Release into Lake Water Mesocosms. Appl. Environ. Microbiol. 1992, 58, 2201–2210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, C.M.; Long, J.A.H.; Donald, M.; Ashbolt, N.J. Survival of Fecal Microorganisms in Marine and Freshwater Sediments. Appl. Environ. Microbiol. 1995, 61, 1888–1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujioka, R.S.; Hashimoto, H.H.; Siwak, E.B.; Young’, R.H.F. Effect of Sunlight on Survival of Indicator Bacteria in Seawater. Appl. Environ. Microbiol. 1981, 41, 690–696. [Google Scholar] [CrossRef] [Green Version]
- Kapuscinski, R.B.; Mitchell, R. Sunlight-Induced Mortality of Viruses and Escherichia Coli in Coastal Seawater. Environ. Sci. Technol. 1983, 17, 1–6. [Google Scholar] [CrossRef]
- Davies-Colley, R.J.; Donnison, A.M.; Speed, D.J.; Ross, C.M.; Nagels, J.W. Inactivation of Faecal Indicator Micro-Organisms in Waste Stabilisation Ponds: Interactions of Environmental Factors with Sunlight. Water Res. 1999, 33, 1220–1230. [Google Scholar] [CrossRef]
- Farrell, C.; Hassard, F.; Jefferson, B.; Leziart, T.; Nocker, A.; Jarvis, P. Turbidity Composition and the Relationship with Microbial Attachment and UV Inactivation Efficacy. Sci. Total Environ. 2018, 624, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Madge, B.A.; Jensen, J.N. Ultraviolet Disinfection of Fecal Coliform in Municipal Wastewater: Effects of Particle Size. Water Environ. Res. 2006, 78, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Droppo, I.G.; Liss, S.N.; Williams, D.; Nelson, T.; Jaskot, C.; Trapp, B. Dynamic Existence of Waterborne Pathogens within River Sediment Compartments. Implications for Water Quality Regulatory Affairs. Environ. Sci. Technol. 2009, 43, 1737–1743. [Google Scholar] [CrossRef]
- Thornton, C.I.; Abt, S.R.; Clary, W.P. Vegetation influence on small stream siltation. J. Am. Water Resour. Assoc. 2007, 33, 1279–1288. [Google Scholar] [CrossRef]
- Buckley, R.; Clough, E.; Warnken, W.; Wild, C. Coliform Bacteria in Streambed Sediments in a Subtropical Rainforest Conservation Reserve. Water Res. 1998, 32, 1852–1856. [Google Scholar] [CrossRef]
- Crabill, C.; Donald, R.; Snelling, J.; Foust, R.; Southam, G. The Impact of Sediment Fecal Coliform Reservoirs on Seasonal Waterquality in Oak Creek, Arizona. Water Res. 1999, 33, 2163–2171. [Google Scholar] [CrossRef]
- Smith, J.; Edwards, J.; Hilger, H.; Steck, T.R. Sediment Can Be a Reservoir for Coliform Bacteria Released into Streams. J. Gen. Appl. Microbiol. 2008, 54, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Jamieson, R.C.; Joy, D.M.; Lee, H.; Kostaschuk, R.; Gordon, R.J. Resuspension of Sediment-Associated Escherichia Coli in a Natural Stream. J. Environ. Qual. 2005, 34, 581–589. [Google Scholar] [CrossRef]
- Fries, J.S.; Characklis, G.W.; Noble, R.T. Sediment-Water Exchange of Vibrio Sp. and Fecal Indicator Bacteria: Implications for Persistence and Transport in the Neuse River Estuary, North Carolina, USA. Water Res. 2008, 42, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.E.; Stocker, M.D.; Hill, R.L.; Pachepsky, Y.A. The Effect of Temperature Oscillations and Sediment Texture on Fecal Indicator Bacteria Survival in Sediments. Water Air Soil Pollut. 2019, 230, 1–11. [Google Scholar] [CrossRef]
- Nowicki, S.; DeLaurent, Z.R.; de Villiers, E.P.; Githinji, G.; Charles, K.J. The Utility of Escherichia Coli as a Contamination Indicator for Rural Drinking Water: Evidence from Whole Genome Sequencing. PLoS ONE 2021, 16, e0245910. [Google Scholar] [CrossRef]
- Noble, R.T.; Lee, I.M.; Schiff, K.C. Inactivation of Indicator Micro-Organisms from Various Sources of Faecal Contamination in Seawater and Freshwater. J. Appl. Microbiol. 2004, 96, 464–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenstark, A. Mutagenic and Lethal Effects of Visible and Near-Ultraviolet Light on Bacterial Cells. Adv. Genet. 1971, 16, 167–198. [Google Scholar] [CrossRef] [PubMed]
- Ziegelhoffer, E.C.; Donohue, T.J. Bacterial Responses to Photo-Oxidative Stress. Nat. Rev. Microbiol. 2009, 7, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Kay, D.; Stapleton, C.M.; Wyer, M.D.; McDonald, A.T.; Crowther, J.; Paul, N.; Jones, K.; Francis, C.; Watkins, J.; Wilkinson, J.; et al. Decay of Intestinal Enterococci Concentrations in High-Energy Estuarine and Coastal Waters: Towards Real-Time T90 Values for Modelling Faecal Indicators in Recreational Waters. Water Res. 2005, 39, 655–667. [Google Scholar] [CrossRef]
- Milliman, J.D. Sediment Discharge to the Ocean from Small Mountainous Rivers: The New Guinea Example. Geo-Marine Lett. 1995, 15, 127–133. [Google Scholar] [CrossRef]
- Tavares Carneiro, M.; Bandeira Vianna Cortes, M.; Cesar Wasserman, J. Critical Evaluation of the Factors Affecting Escherichia Coli Environmental Decay for Outfall Plume Models. Rev. Ambient. Água 2018, 13. [Google Scholar] [CrossRef]
- Le, H.A.; Gratiot, N.; Santini, W.; Ribolzi, O.; Tran, D.; Meriaux, X.; Deleersnijder, E.; Soares-Frazão, S. Suspended Sediment Properties in the Lower Mekong River, from Fluvial to Estuarine Environments. Estuar. Coast. Shelf Sci. 2020, 233, 106522. [Google Scholar] [CrossRef]
- Wang, C.; Zheng, S.-S.; Wang, P.-F.; Hou, J. Interactions between Vegetation, Water Flow and Sediment Transport: A Review. J. Hydrodyn. 2015, 27, 24–37. [Google Scholar] [CrossRef]
- Teeter, A.M.; Johnson, B.H.; Berger, C.; Stelling, G.; Scheffner, N.W.; Garcia, M.H.; Parchure, T.M. Hydrodynamic and Sediment Transport Modeling with Emphasis on Shallow-Water, Vegetated Areas (Lakes, Reservoirs, Estuaries and Lagoons). Hydrobiologia 2001, 444, 1–23. [Google Scholar] [CrossRef]
- Morgan, J.A.; Hoet, A.E.; Wittum, T.E.; Monahan, C.M.; Martin, J.F. Reduction of Pathogen Indicator Organisms in Dairy Wastewater Using an Ecological Treatment System. J. Environ. Qual. 2008, 37, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Scholz, M. Wetland Systems: Storm Water Management Control; Springer Science & Business Media: Berlin, Germany, 2010. [Google Scholar]
- Sani, A.; Scholz, M.; Bouillon, L. Seasonal Assessment of Experimental Vertical-Flow Constructed Wetlands Treating Domestic Wastewater. Bioresour. Technol. 2013, 147, 585–596. [Google Scholar] [CrossRef]
- Paing, J.; Guilbert, A.; Gagnon, V.; Chazarenc, F. Effect of Climate, Wastewater Composition, Loading Rates, System Age and Design on Performances of French Vertical Flow Constructed Wetlands: A Survey Based on 169 Full Scale Systems. Ecol. Eng. 2015, 80, 46–52. [Google Scholar] [CrossRef]
- Almuktar, S.A.A.A.N.; Abed, S.N.; Scholz, M. Wetlands for Wastewater Treatment and Subsequent Recycling of Treated Effluent: A Review. Environ. Sci. Pollut. Res. 2018, 25, 23595–23623. [Google Scholar] [CrossRef] [Green Version]
- Mthembu, M.; Odinga, C.; Swalaha, F.M.; Bux, F. Constructed Wetlands: A Future Alternative Wastewater Treatment Technology. Afr. J. Biotechnol. 2013, 12, 4542–4553. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | RL | DL | RD | DD |
|---|---|---|---|---|
| Total | 0.50 ± 0.15 | 1.43 ± 0.15 | −0.14 ± 0.37 | 1.17 ± 0.13 |
| Attached | 0.49 ± 0.15 | 1.41 ± 0.15 | −0.15 ± 0.38 | 1.18 ± 0.13 |
| Free | 1.20 ± 0.13 | 1.53 ± 0.19 | 0.28 ± 0.35 | 1.36 ± 0.32 |
| Treatments | RL | DL | RD | DD | ||||
|---|---|---|---|---|---|---|---|---|
| T50 | T90 | T50 | T90 | T50 | T90 | T50 | T90 | |
| Total | 33.4 | 111 | 11.6 | 38.6 | 117.8 | 391.2 | 14.2 | 47.2 |
| Attached | 33.9 | 112.8 | 11.8 | 39.2 | 110.9 | 368.4 | 14.1 | 46.8 |
| Free | 13.9 | 46 | 10.9 | 36.1 | 59.4 | 197.4 | 12.2 | 40.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakhle, P.; Boithias, L.; Pando-Bahuon, A.; Thammahacksa, C.; Gallion, N.; Sounyafong, P.; Silvera, N.; Latsachack, K.; Soulileuth, B.; Rochelle-Newall, E.J.; et al. Decay Rate of Escherichia coli in a Mountainous Tropical Headwater Wetland. Water 2021, 13, 2068. https://doi.org/10.3390/w13152068
Nakhle P, Boithias L, Pando-Bahuon A, Thammahacksa C, Gallion N, Sounyafong P, Silvera N, Latsachack K, Soulileuth B, Rochelle-Newall EJ, et al. Decay Rate of Escherichia coli in a Mountainous Tropical Headwater Wetland. Water. 2021; 13(15):2068. https://doi.org/10.3390/w13152068
Chicago/Turabian StyleNakhle, Paty, Laurie Boithias, Anne Pando-Bahuon, Chanthamousone Thammahacksa, Nicolas Gallion, Phabvilay Sounyafong, Norbert Silvera, Keooudone Latsachack, Bounsamay Soulileuth, Emma J. Rochelle-Newall, and et al. 2021. "Decay Rate of Escherichia coli in a Mountainous Tropical Headwater Wetland" Water 13, no. 15: 2068. https://doi.org/10.3390/w13152068
APA StyleNakhle, P., Boithias, L., Pando-Bahuon, A., Thammahacksa, C., Gallion, N., Sounyafong, P., Silvera, N., Latsachack, K., Soulileuth, B., Rochelle-Newall, E. J., Marcangeli, Y., Pierret, A., & Ribolzi, O. (2021). Decay Rate of Escherichia coli in a Mountainous Tropical Headwater Wetland. Water, 13(15), 2068. https://doi.org/10.3390/w13152068