
Estimation of Populuseuphratica Forest Leaf Litterfall and Time Variation of Nutrient in Leaf Litter during Decomposition along the Main Channel of the Tarim River, China



Abstract
:1. Introduction
2. Materials and Methods
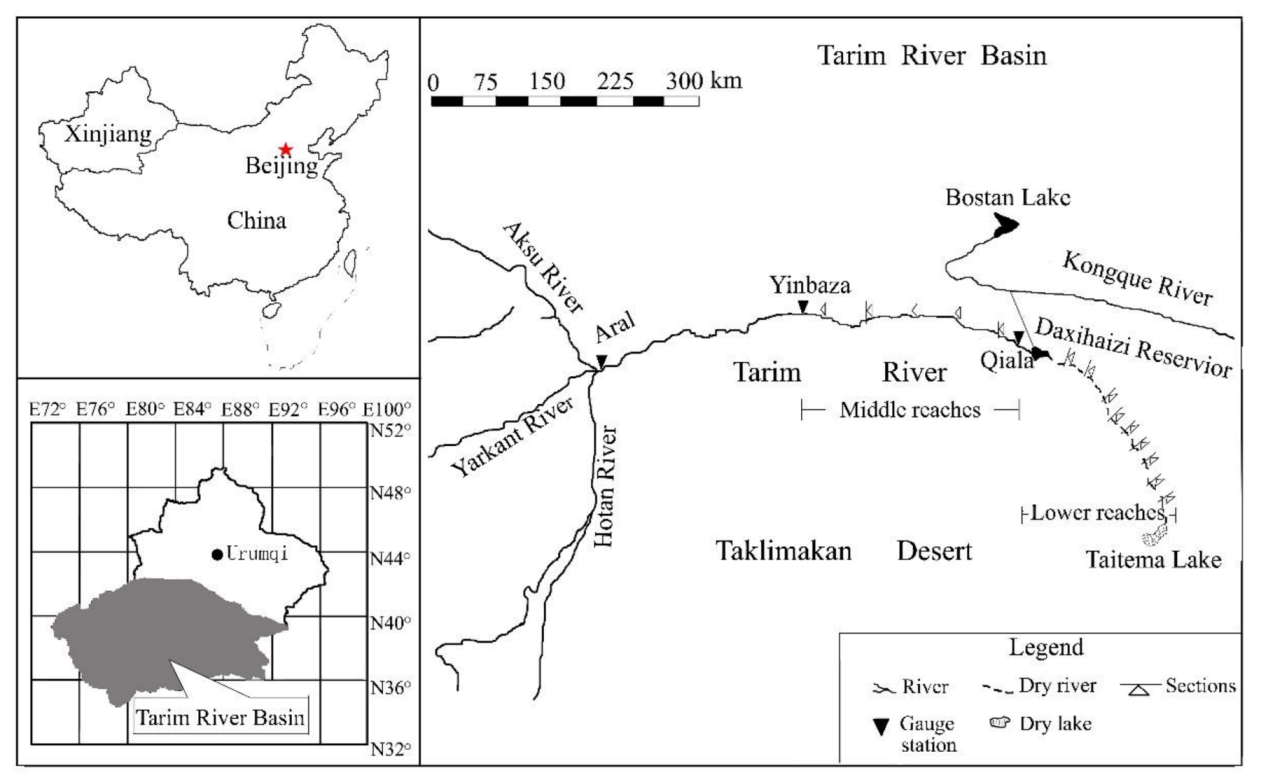
2.1. Study Area
2.2. Annual Leaf Litter Amount and Tree Characteristics of Individual P. euphratica Trees
2.3. The Population Density of P. euphratica and Area of P. euphratica Forest
2.4. Litterbag Experiment
2.5. Nutrient Chemical Analysis
2.6. Data Analysis
2.7. Statistical Analysis
3. Results
3.1. Equation between Leaf Litter Amount and Relevant Tree Characteristics on an Individual Scale
3.2. Leaf Litter Amount per Tree
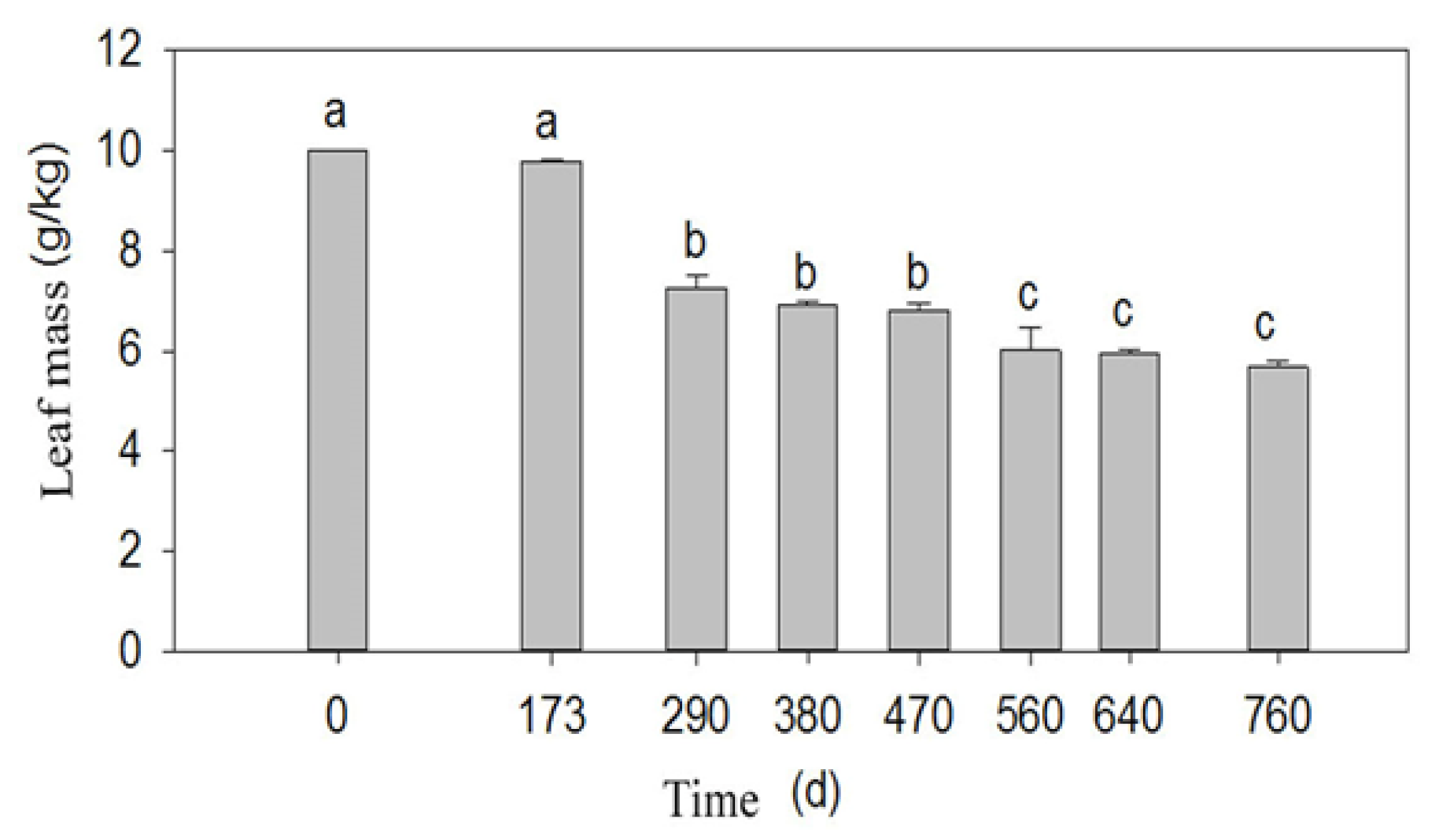
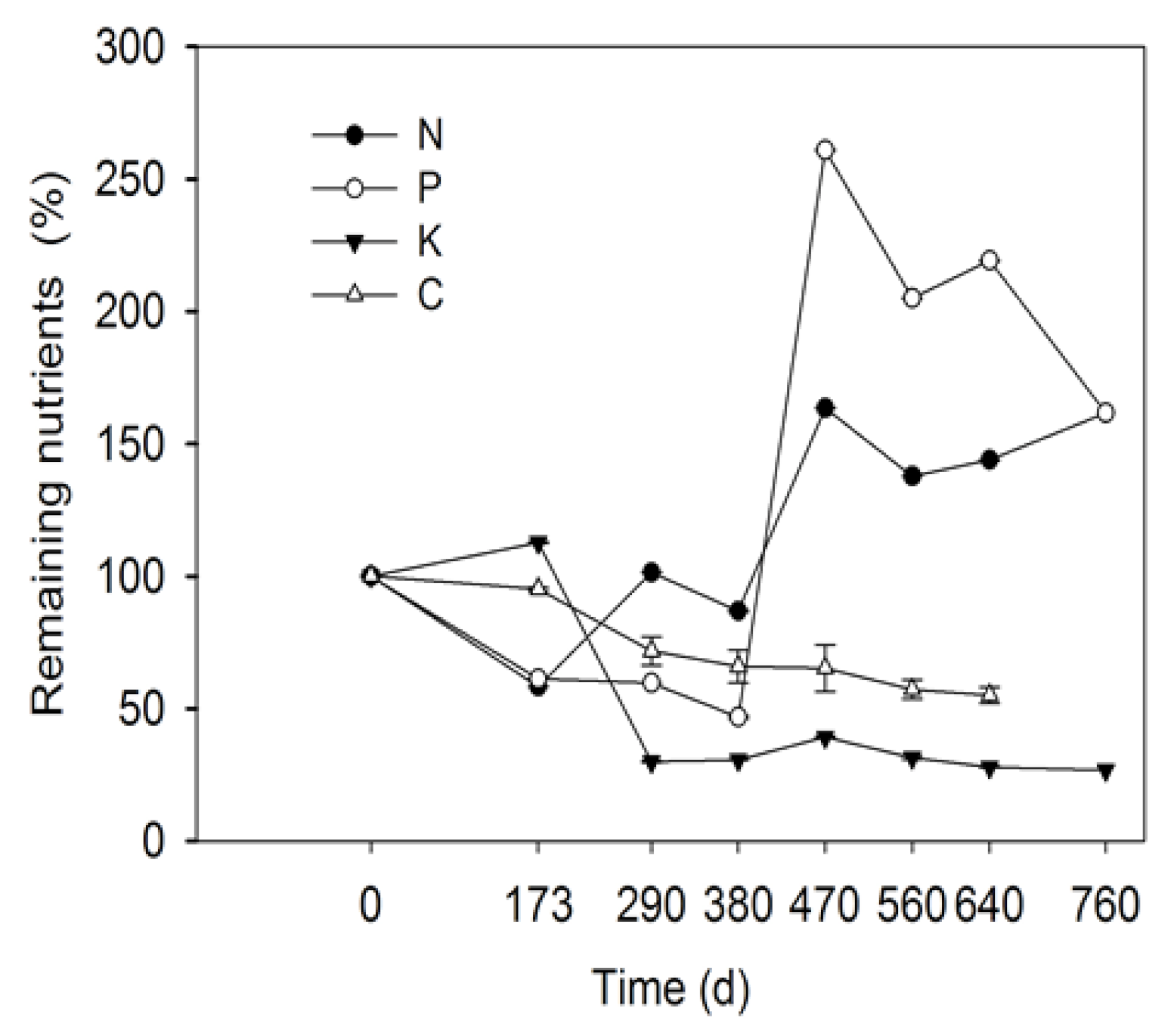
3.3. Change in Leaf Litter Mass and Nutrients over Time during Decay
4. Discussion
4.1. Leaf Litter Amount Estimation
4.2. Leaf Litter Decomposition
4.3. Ecological Effects of Leaf Litter Amount Change
5. Conclusions
- (1)
- The equation between canopy area (x) and annual leaf litter amount (y) for a P. euphratica tree was y = e(11.12 − 10.97/x) (R2 = 0.39, p < 0.05), which may be appropriate to estimate the average annual leaf litter amount for a tree. The mean annual leaf litter amount per tree was 10.2–14.83 kg/y. Loss of P. euphratica forest decreased the annual leaf litter amount.
- (2)
- Leaf litter mass changed with incubation time and exhibited three phases: an initial slow decomposition phase (0–173 d) with mass loss; a rapid mass loss phase (173–470 d); and a second rapid mass loss phase (470–560 d). Overall, C and K content decreased, while N and P content increased in a fluctuating manner within 760 d. Loss of the leaf litter would lead to a decrease in nutrient sources for plant and microorganisms in desert riparian forest.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, S.J. The Status, conservation and recovery of global resources of Populus euphradica. World For. Res. 1996, 6, 37–44. (In Chinese) [Google Scholar]
- Chen, Y.P.; Chen, Y.N.; Xu, C.C. Effects of groundwater depth on the gas exchange and chlorophy II fluorescence of Populus euphratica in the lower reaches of Tarim River. Acta Ecol. Sin. 2011, 31, 344–353. (In Chinese) [Google Scholar]
- Hao, X.M.; Li, W.H.; Huang, X.; Zhu, C.G.; Ma, J.X. Assessment of the groundwater threshold of desert riparian forest vegetation along the middle and lower reaches of the Tarim River, China. Hydrol. Process. 2010, 24, 178–186. [Google Scholar] [CrossRef]
- Ling, H.B.; Zhang, P.; Xu, H.L.; Zhao, X.F. How to regenerate and protect desert riparian Populus euphratica forest in arid areas. Sci. Rep. 2015, 5, 15418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.N.; Wumaierjiang, W.; Aikeremu, A.; Chen, Y.; Chen, Y.P.; Hao, X.M.; Zhu, C.G.; Wang, Y. Monitoring and analysis of ecological benefits of water conveyance in the lower reaches of Tarim River in recent 20 years. Arid. Land Geogr. 2021, 44, 605–611. (In Chinese) [Google Scholar]
- Liu, J.Z.; Chen, Y.N.; Li, W.H.; Chen, Y.P. Analysis on the distribution and degraded succession of plant communities in the lower reaches of Tarim River. Acta Ecol. Sin. 2004, 24, 379–383. (In Chinese) [Google Scholar]
- Shi, J.H.; Liu, M.X.; Wang, X.Y.; Ma, X.X. Biomass and stoichiometry of dead branches of standing trees in Populus euphratica forests with different ages. Acta Ecol. Sin. 2017, 37, 887–895. (In Chinese) [Google Scholar]
- Wang, R.Z.; Chen, Y.P.; Chen, Y.N.; Pan, Y.P.; He, G.Z. Effects of groundwater level on morphologica, anatomical structure and leaf hydraulic conductance of Populus euphratica. J. Desert Res. 2016, 36, 1303–1309. (In Chinese) [Google Scholar]
- Campos, C.A.; Cruz, H.L.; Rocha, O.S. Mass, nutrient pool, and mineralization of litter and fine roots in a tropical mountain cloud forest. Sci. Total Environ. 2017, 575, 876–886. [Google Scholar] [CrossRef]
- Alhamd, L.; Arakaki, S.; Hagihara, A. Decomposition of leaf litter of four tree species in a subtropical evergreen broad-leaved forest, Okinawa Island, Japan. For. Ecol. Manag. 2004, 202, 1–11. [Google Scholar] [CrossRef]
- Bray, J.R.; Gorham, E. Litter production in forests of the world. Adv. Ecol. Res. 1964, 2, 101–157. [Google Scholar]
- Zhang, H.C.; Yuan, W.P.; Dong, W.J.; Liu, S.G. Seasonal patterns of litterfall in forest ecosystem worldwide. Ecol. Complex 2014, 20, 240–247. [Google Scholar] [CrossRef]
- Peichl, M.; Leava, N.A.; Kiely, G. Above- and belowground ecosystem biomass, carbon and nitrogen allocation in recently afforested grassland and adjacent intensively managed grassland. Plant Soil 2012, 350, 281–296. [Google Scholar] [CrossRef]
- Preece, N.D.; Ceowley, G.M.; Lawes, M.J.; Van Oosterzee, P. Comparing above-ground biomass among forest types in the Wet Tropics: Small stems and plantation types matter in carbon accounting. For. Ecol. Manag. 2012, 264, 228–237. [Google Scholar] [CrossRef]
- Keller, M.; Palace, M.; Hurtt, G. Biomass estimation in the Tapajos National Forest, Brazil: Examination of sampling and allometric uncertainties. For. Ecol. Manag. 2001, 154, 371–382. [Google Scholar] [CrossRef]
- Ter-Mikaelian, M.T.; Korzukhin, M.D. Biomass equations for sixty-five North American tree species. For. Ecol. Manag. 1997, 97, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.Y.; Zhang, Z.W.; Li, H.K. Biomass equations of primary arbour tree species in China. J. Cent. South Univ. For. Technol. 2019, 39, 63–69. (In Chinese) [Google Scholar]
- Yang, Y.H.; Chen, Y.N.; Li, Z.; Chen, Y.P. Land–use/cover conversion affects soil organic-carbon stocks: A case study along the main channel of the Tarim River, China. PLoS ONE 2018, 13, e0206903. [Google Scholar] [CrossRef] [PubMed]
- Bai, G.Z. Study of Populus euphratic Community Pattern in the Upper, Middle and Lower Reaches of Tarim River. Master’s Thesis, Tarim University, Aral, Xinjiang, China, 2010. (In Chinese). [Google Scholar]
- Yang, Y.H.; Zhou, H.H.; Ye, Z.X.; Zhu, C.G.; Chen, Y.N. Effects of transient flooding on leaf litter decomposition: A case study of Populue euphratica leaf in an arid area. Int. J. Agric. Biol. 2019, 22, 1385–1392. [Google Scholar]
- Lu, R.K. Analytical Methods for Soil Agrochemistry; China Agricultural Science and Technology Press: Beijing, China, 1999. (In Chinese) [Google Scholar]
- Dong, D.R.; Li, X.; Wan, H.M.; Lin, H.J. Above biomass estimation of Populus euphratica in the lower reaches of Tarim River. J. Desert Res. 2013, 33, 724–730. (In Chinese) [Google Scholar]
- Zhang, H.; Li, J.W.; Li, J.Q.; Zhang, Y.B.; Sun, L. Branching patterns of Populus euphratica in the Ejina Oasis of Inner Mongolia. J. Northwest For. Univ. 2009, 24, 46–49. (In Chinese) [Google Scholar]
- Liu, M.X.; Shi, J.H.; Wang, X.Y.; Ma, X.X. Aboveground biomass and carbon storage of Populus euphratica plantation with different ages in Tahe River Basin. Bull. Soil Water Conserv. 2016, 36, 326–339. (In Chinese) [Google Scholar]
- Staelens, J.; Nachtergale, L.; An, D.S.; Vanhellemont, M.; Wuyts, K.; Verheyen, K. Spatio-temporal litterfall dynamics in a 60-year-old mixed deciduous forest. Ann. Sci. 2011, 68, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Fukuzawa, K.; Shibata, H. Temporal changes in litterfall, litter decomposition and their chemical composition in Sasa dwarf bamboo in a natural forest ecosystem of northern Japan. J. For. Res. 2013, 18, 129–138. [Google Scholar] [CrossRef]
- Zhou, J.; Lang, X.; Du, B.; Zhang, H.; Liu, H.; Zhang, Y.; Shang, L. Litterfall and nutrient return in moist evergreen broad-leaved primary forest and mixed subtropical secondary deciduous broad-leaved forest in China. Eur. J. For. Res. 2016, 135, 77–86. [Google Scholar] [CrossRef]
- Zhu, X.A.; Liu, W.J.; Chen, H.; Deng, Y.; Chen, C.F.; Zeng, H.H. Effects of forest transition on litterfall, standing litter and related nutrient returns: Implications for forest management in tropical China. Geoderma 2019, 333, 123–134. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, S.; Huang, Y. Comparisons of litterfall, litter decomposition and nutrient return in a monoculture Cunninghamia lanceolata and a mixed stand in southern China. For. Ecol. Manag. 2008, 255, 1210–1218. [Google Scholar] [CrossRef]
- Fan, C.N.; Guo, Z.L.; Zheng, J.P.; Li, B.; Yang, B.G.; Yue, L.; Yu, H.B. The amount and dynamics of litterfall in the natural secondary forest in Mopan Mountain. Acta Ecol. Sin. 2014, 34, 633–641. (In Chinese) [Google Scholar]
- Wanf, F.Y. Review on the study of forest litterfall. Adv. Ecol. 1989, 6, 82–89. [Google Scholar]
- IPCC 2014. Climate Change 2014: Mitigation of Climate Change; Edenhofer, O., Pichs-Madruga, R., Sokona, Y., Farahani, E., Kadner, S., Seyboth, K., Adler, A., Baum, I., Brunner, S., Eickemeier, P., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- Hansen, K.; Vesterdal, L.; Schmidt, I.K.; Gundersen, P.; Sevel, L.; Basturp-Birk, A.; Pedersen, L.B.; Bille-Hansen, J. Litterfall and nutrient return in five tree species in a common garden experiment. For. Ecol. Manag. 2009, 257, 2133–2144. [Google Scholar] [CrossRef]
- Pan, H.; Huang, S.D.; Hong, W.; Zhu, H.R.; Zhang, Z.H.; Lin, J. The carbon storage and distribution in three Acacia plantations. J. Fujian Coll. For. 2009, 29, 28–32. (In Chinese) [Google Scholar]
- Xie, H.H.; Ma, W.Y.; Zhao, C.Y.; Gao, Y.F.; Wang, Q.T.; Ge, H.Y.; Lin, M.; Huang, H.Y. Effects of litter and moss on soil respiration in a spruce forest in the Qilian Mountains, Qinhai. Acta Ecol. Sin. 2017, 37, 1379–1390. (In Chinese) [Google Scholar]
- Huang, W.J.; Spohn, M. Effects of long-term litter manipulation on soil carbon, nitrogen, and phosphorus in a temperate deciduous forest. Soil Biol. Biochem. 2015, 83, 12–18. [Google Scholar] [CrossRef]
- Xu, S.; Liu, L.L.; Sayer, E.J. Variability of above-ground litter inputs alters soil physicochemical and biological processes: A meta-analysis of litterfall manipulation experiments. Biogeosciences 2013, 10, 7423–7433. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.L.; Zeng, D.H.; Ma, X.Q.; Chang, S.X. Root rather than leaf litter input drives soil carbon sequestration after afforestation on a marginal cropland. For. Ecol. Manag. 2016, 362, 38–45. [Google Scholar] [CrossRef]
- Wan, X.H.; Xiao, L.Q.; Vadeboncoeur, M.A.; Johnson, C.H.; Huang, Z.Q. Response of mineral soil carbon storage to harvest residue retention depends on soil texture: A meta-analysis. For. Ecol. Manag. 2018, 408, 9–15. [Google Scholar] [CrossRef]
- Tian, L.; Zhao, L.; Wu, X.; Fang, H.; Zhao, Y.; Yue, G.; Liu, G.; Chen, H. Vertical patterns and controls of soil nutrients in alpine grassland: Implications for nutrient uptake. Sci. Total Environ. 2017, 607, 855–864. [Google Scholar] [CrossRef]
- Bernaschini, M.L.; Moreno, M.L.; Perez-Harguindeguy, N.; Valladares, G. Is litter decomposition influenced by forest size and invertebrate detritivores during the dry season in semiarid Chaco Serrano? J. Arid Environ. 2016, 127, 154–159. [Google Scholar] [CrossRef]
- Miranda, A.; Altamirano, A.; Cayuela, L.; Pincheira, F.; Lara, A. Different times, same story: Native forest loss and landscape homogenization in three physiographical areas of south-central of Chile. Appl. Geogr. 2015, 60, 20–28. [Google Scholar] [CrossRef]
- Zhao, S.J.; Wei, Q.; Zhao, X.F. Analysis of the change process and land use types in the main stream of Tarim River. J. Xinjiang Agric. Univ. 2018, 41, 298–306. [Google Scholar]
- Zhang, S.; Wang, Y.; Xia, T.T.; Chang, X.E.; Li, Z. Effects of land use/cover change on ecosystem service value under the ecological water conveyance of Tarim River. Arid. Land Geogr. 2021, 44, 741–749. (In Chinese) [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Equation | R2 | F | df | Sig. | b0 | b1 |
|---|---|---|---|---|---|---|
| Compound | 0.271 | 6.327 | 18 | 0.022 | 1192.1 | 1.422 |
| Growth | 0.271 | 6.327 | 18 | 0.022 | 7.083 | 0.352 |
| Methods | Minimum(Kg/Year) | Maximum (Kg/Year) | Mean (Kg/Year) |
|---|---|---|---|
| Field survey | 2.05 | 30.70 | 10.20 |
| y = e(11.121−10.97/x) | 3.63 | 22.53 | 14.83 |
| y = 3570.93e0.034x | 0.65 | 4.62 | 2.62 |
| Country/Region | Area (Ha) | Annual Leaf Litter Amount (Kg/Year) | ||
|---|---|---|---|---|
| Minimum | Maximum | Average | ||
| China | 395.200 | 13.15 × 107 | 26.11 × 108 | 8.59 × 108 |
| Central Asia | 200.000 | 6.65 × 107 | 13.21 × 108 | 4.35 × 108 |
| Iraq | 20.000 | 0.66 × 107 | 1.32 × 108 | 0.43 × 108 |
| Iran | 20.000 | 0.66 × 107 | 1.32 × 108 | 0.43 × 108 |
| Syria | 5818 | 0.19 × 107 | 0.38 × 108 | 0.13 × 108 |
| Turkey | 4900 | 0.16 × 107 | 0.33 × 108 | 0.10 × 108 |
| Total | 648.719 | 21.47 × 107 | 42.67 × 108 | 14.01 × 108 |
| Location | Year | Area (Ha) | Population Density (Plants/Ha) | Annual Leaf Litter Amount (Kg/Year) | ||
|---|---|---|---|---|---|---|
| Minimum | Maximum | Average | ||||
| The upper reaches | 1958 | 23.000 | 594.2 | 19.56 × 106 | 38.93 × 107 | 12.82 × 107 |
| 1978 | 58.200 | 49.50 × 106 | 98.53 × 107 | 32.44 × 107 | ||
| 1983 | 112.400 | 95.61 × 106 | 190.3 × 107 | 62.65 × 107 | ||
| The middle reaches | 1958 | 175.800 | 65.3 | 1.64 × 107 | 3.27 × 108 | 1.08 × 108 |
| 1978 | 100.200 | 0.94 × 107 | 1.86 × 108 | 0.61 × 108 | ||
| 1983 | 110.800 | 1.04 × 107 | 2.06 × 108 | 0.68 × 108 | ||
| The lower reaches | 1958 | 54.000 | 36.1 | 2.79 × 106 | 5.55 × 107 | 1.83 × 107 |
| 1978 | 16.400 | 0.85 × 106 | 1.69 × 107 | 0.56 × 107 | ||
| 1983 | 7333 | 0.38 × 106 | 0.75 × 107 | 0.25 × 107 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Zhou, H.; Ye, Z.; Zhu, C. Estimation of Populuseuphratica Forest Leaf Litterfall and Time Variation of Nutrient in Leaf Litter during Decomposition along the Main Channel of the Tarim River, China. Water 2021, 13, 2514. https://doi.org/10.3390/w13182514
Yang Y, Zhou H, Ye Z, Zhu C. Estimation of Populuseuphratica Forest Leaf Litterfall and Time Variation of Nutrient in Leaf Litter during Decomposition along the Main Channel of the Tarim River, China. Water. 2021; 13(18):2514. https://doi.org/10.3390/w13182514
Chicago/Turabian StyleYang, Yuhai, Honghua Zhou, Zhaoxia Ye, and Chenggang Zhu. 2021. "Estimation of Populuseuphratica Forest Leaf Litterfall and Time Variation of Nutrient in Leaf Litter during Decomposition along the Main Channel of the Tarim River, China" Water 13, no. 18: 2514. https://doi.org/10.3390/w13182514
APA StyleYang, Y., Zhou, H., Ye, Z., & Zhu, C. (2021). Estimation of Populuseuphratica Forest Leaf Litterfall and Time Variation of Nutrient in Leaf Litter during Decomposition along the Main Channel of the Tarim River, China. Water, 13(18), 2514. https://doi.org/10.3390/w13182514