
Genetic Signature of a Past Anthropogenic Transportation of a Far-Eastern Endemic Cladoceran (Crustacea: Daphniidae) to the Volga Basin

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
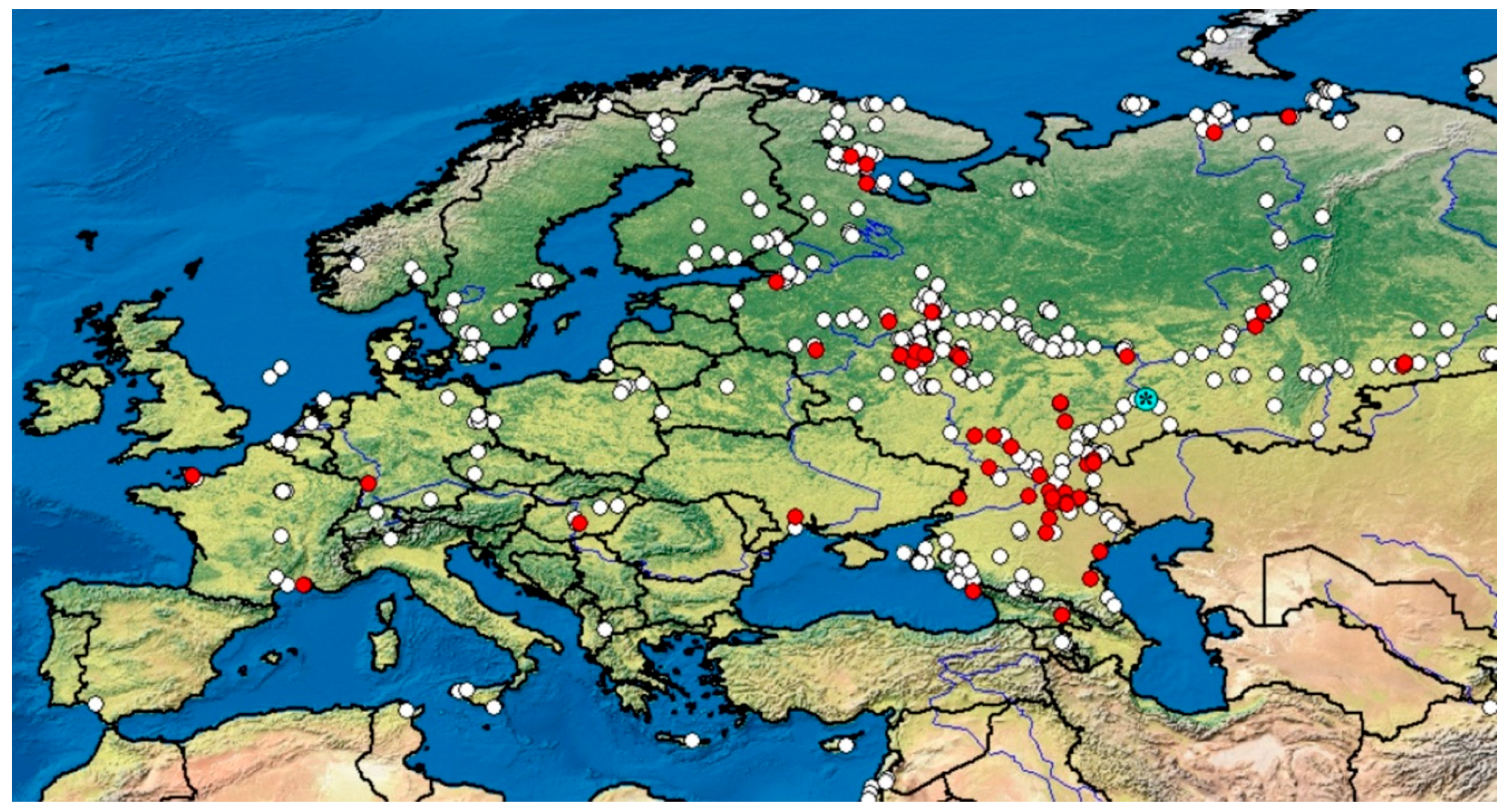
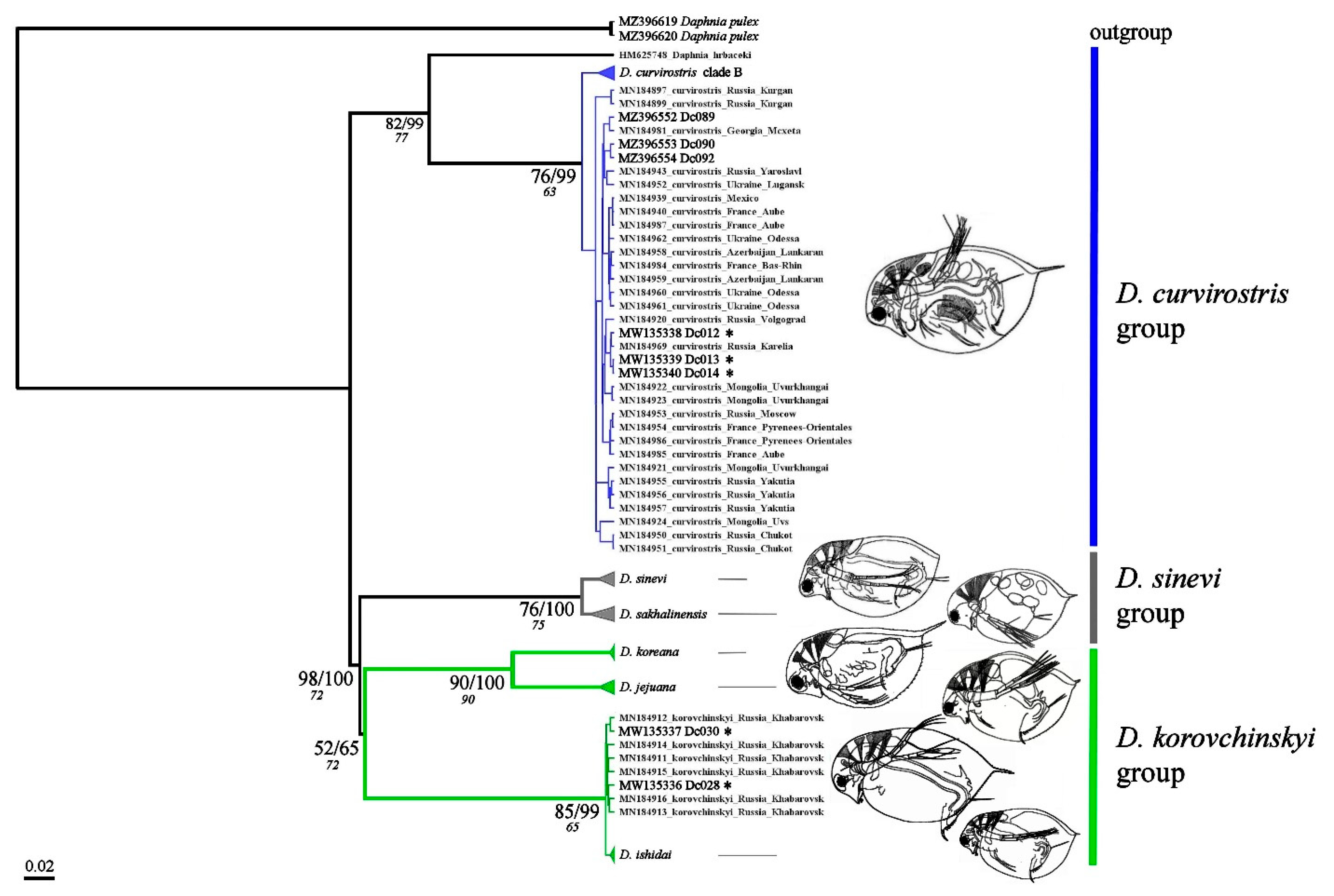
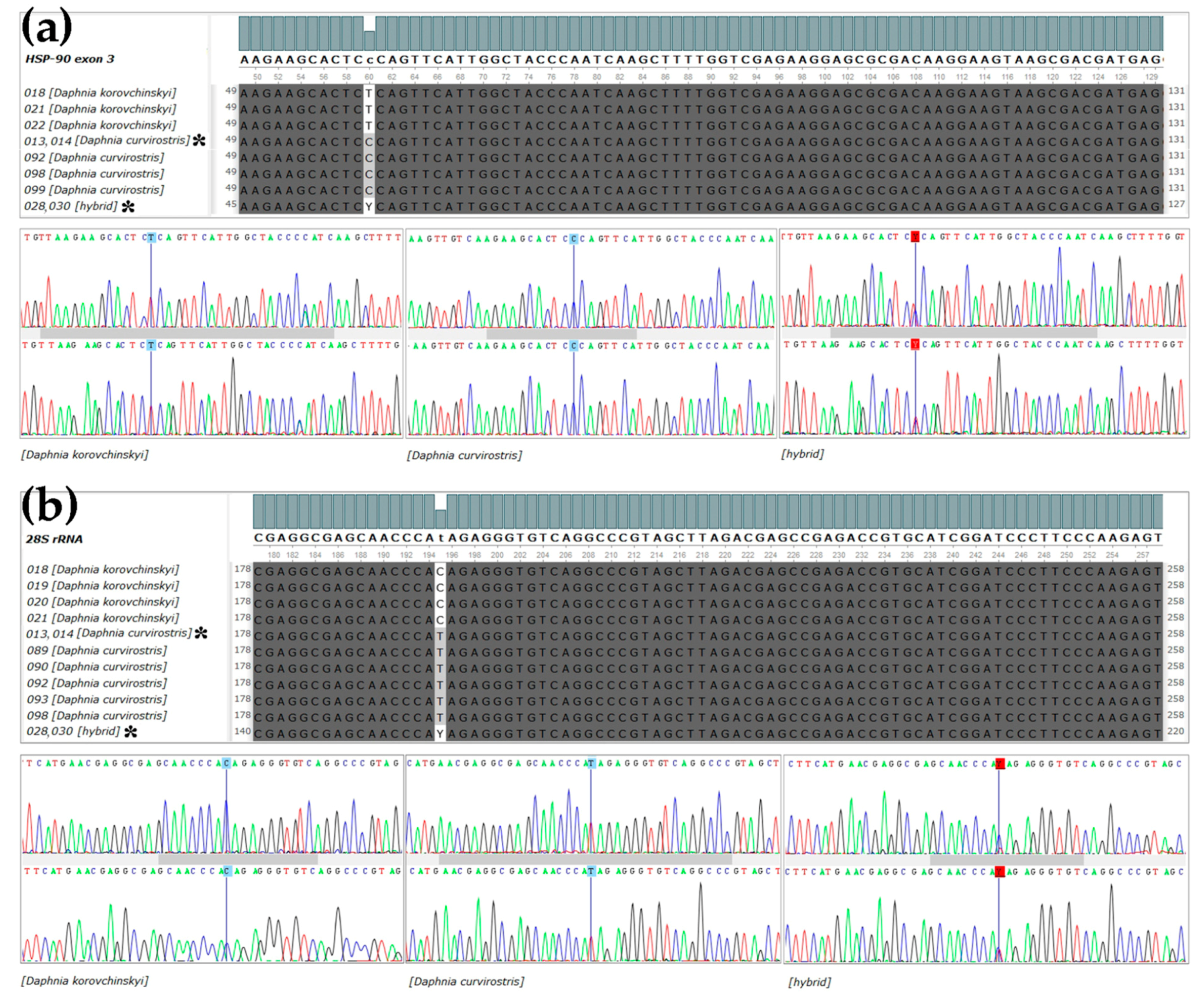
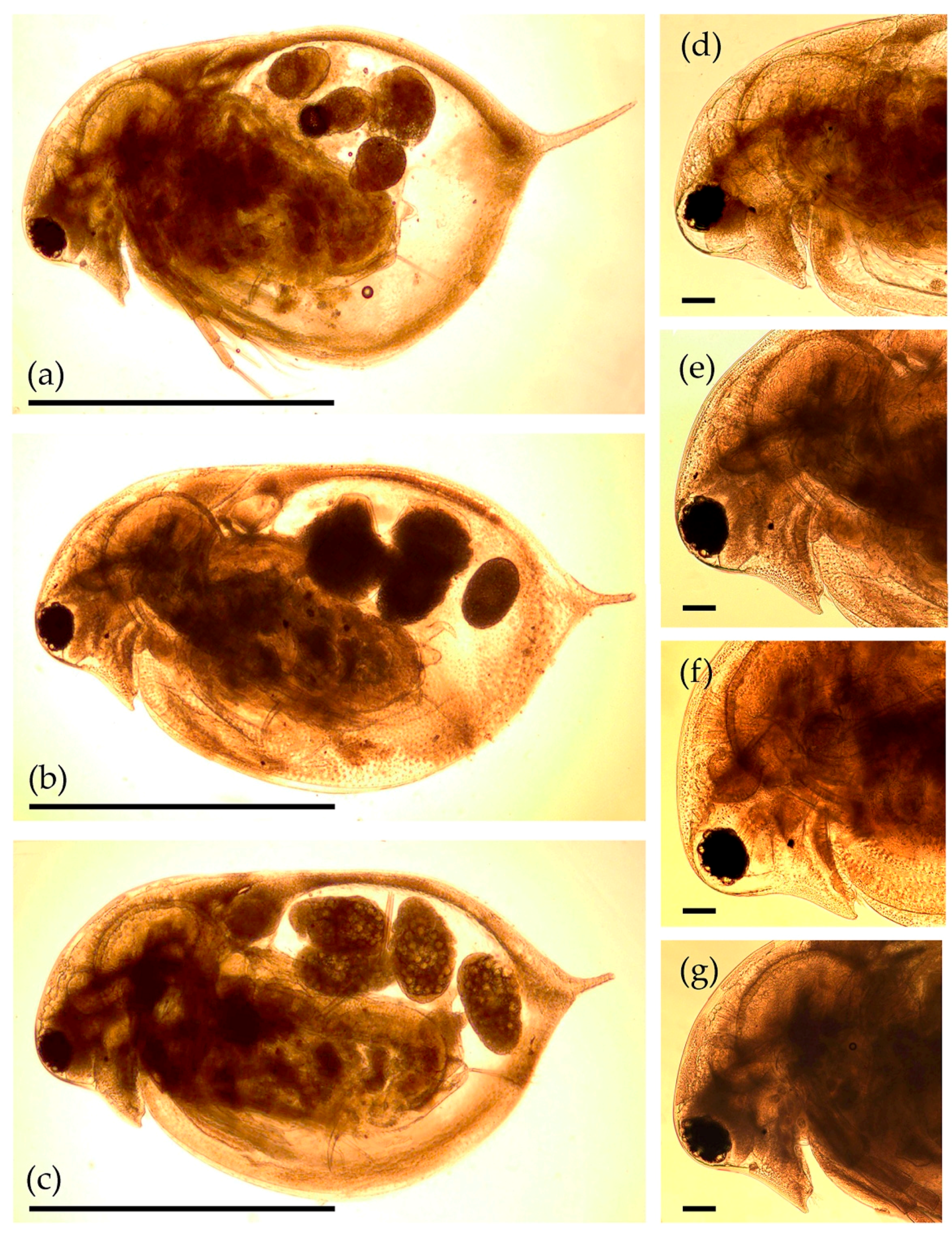
3. Results
3.1. Genetics Account
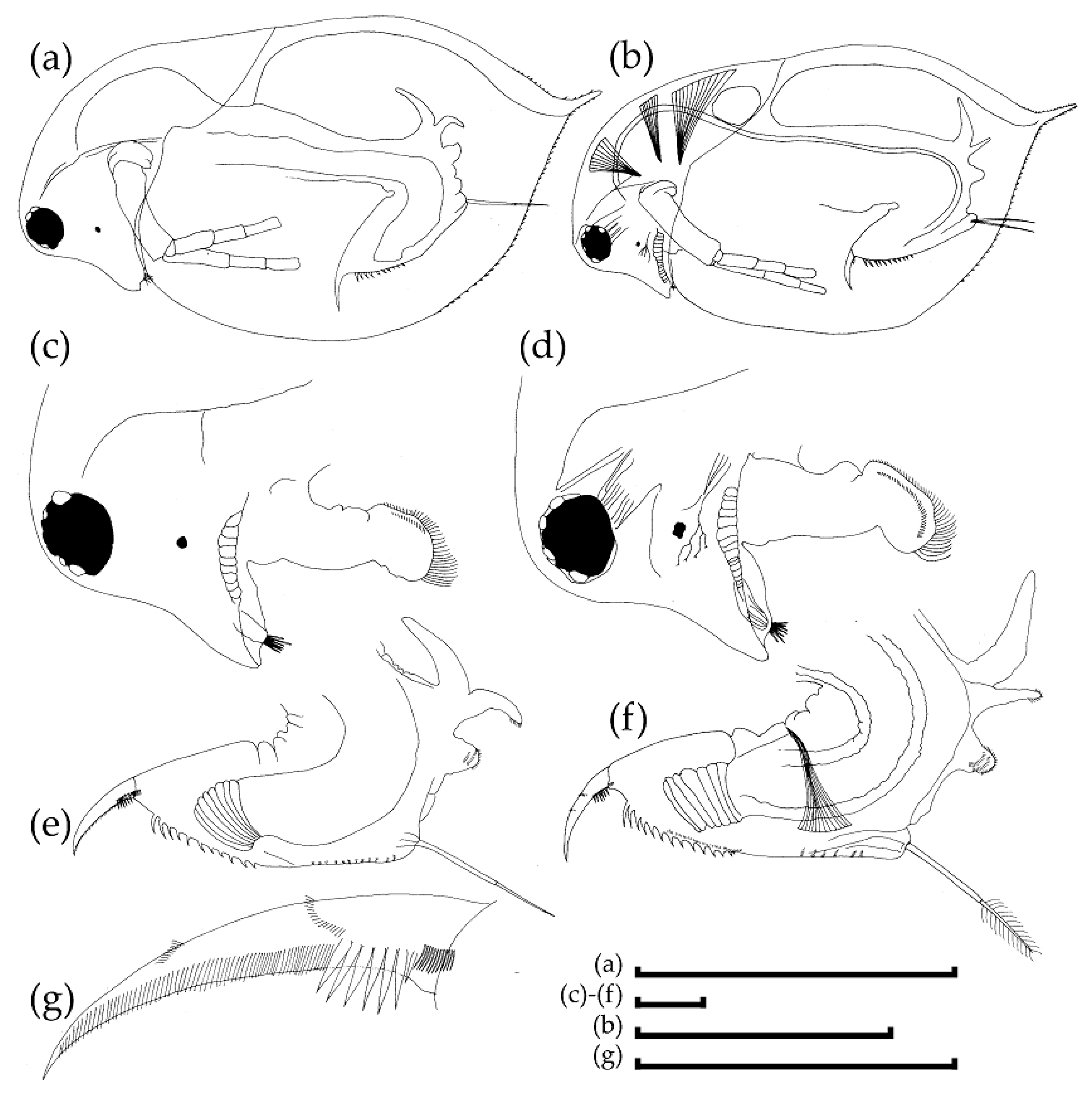
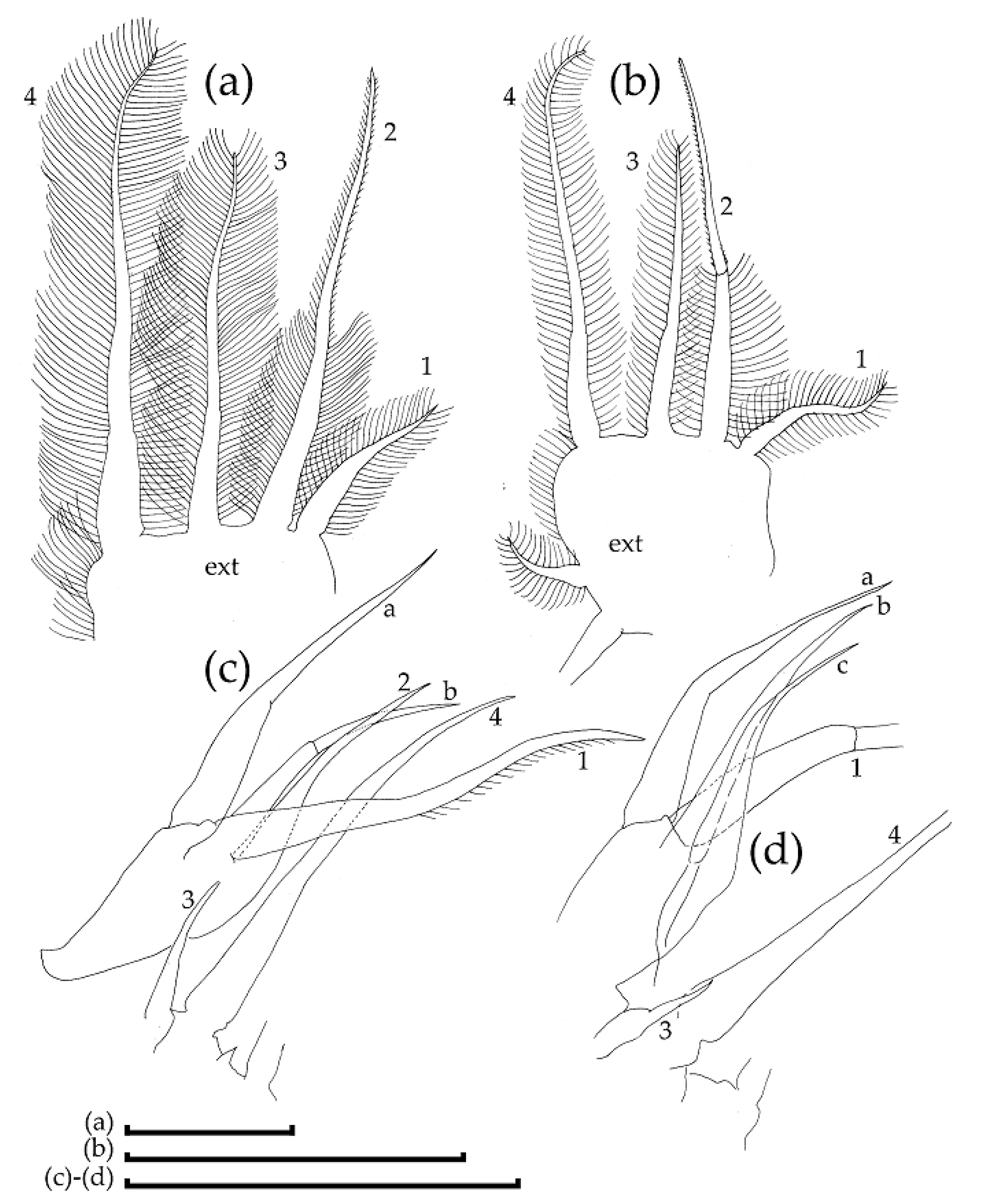
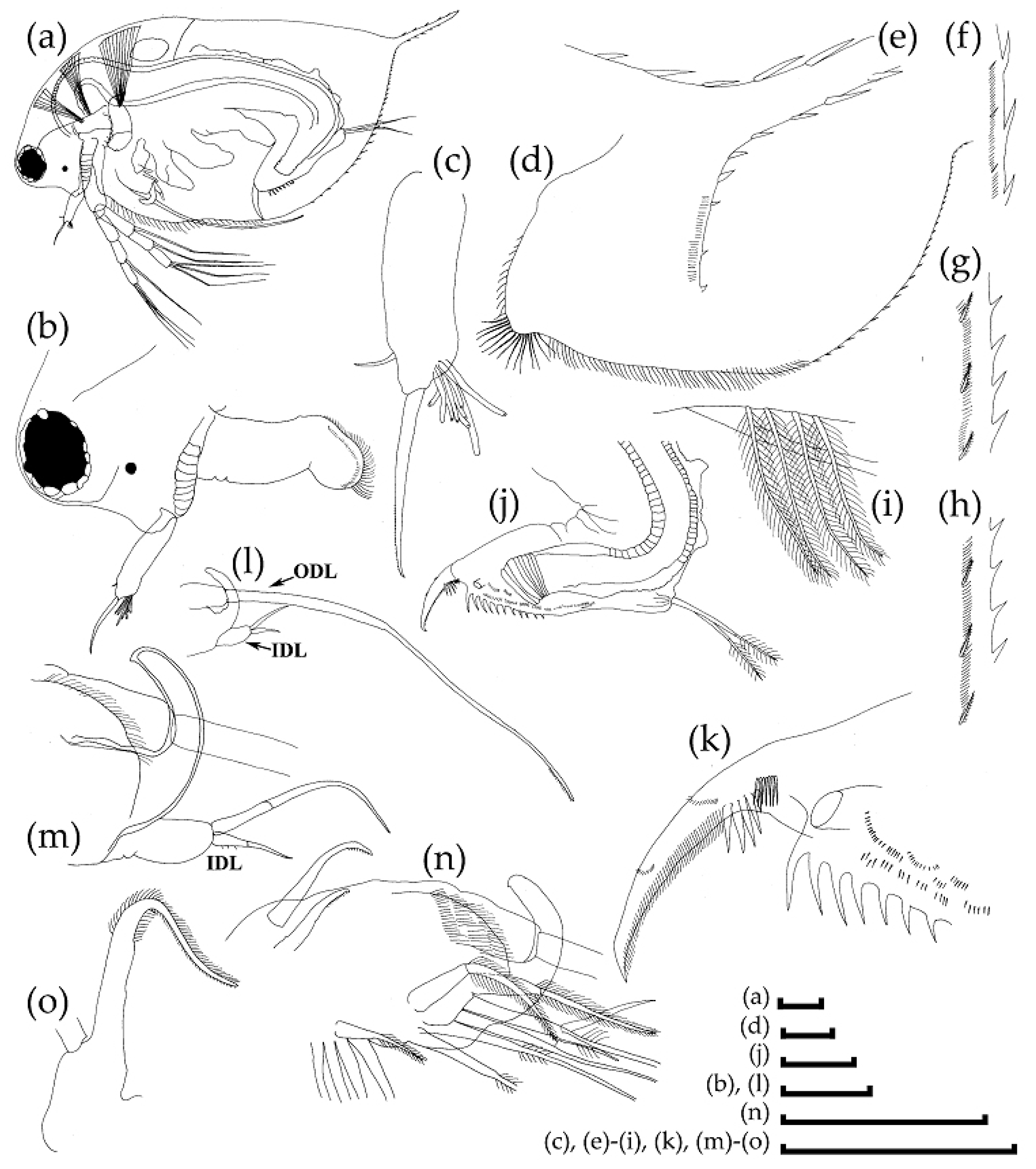
3.2. Morphological Account
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gorokhova, E.; Aladin, N.; Dumont, H.J. Further expansion of the genus Cercopagis (Crustacea, Branchiopoda, Onychopoda) in the Baltic Sea, with notes on the taxa present and their ecology. Hydrobiologia 2000, 429, 207–218. [Google Scholar] [CrossRef]
- Cristescu, M.E.A.; Hebert, P.D.N.; Witt, J.D.S.; MacIsaac, H.J.; Grigorovich, I.A. An invasion history for Cercopagis pengoi based on mitochondrial gene sequences. Limnol. Oceanogr. 2001, 46, 224–229. [Google Scholar] [CrossRef]
- Korovchinsky, N.M.; Arnott, S.E. Taxonomic resolution of the North American invasive species of the genus Bythotrephes Leydig, 1860 (Crustacea: Cladocera: Cercopagididae). Zootaxa 2019, 4691, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Ishida, S.; Taylor, D.J. Quaternary diversification in a sexual Holarctic zooplankter, Daphnia galeata. Mol. Ecol. 2007, 16, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Bekker, E.I.; Karabanov, D.P.; Galimov, Y.R.; Haag, C.R.; Neretina, T.V.; Kotov, A.A. Phylogeography of Daphnia magna Straus (Crustacea: Cladocera) in Northern Eurasia: Evidence for a deep longitudinal split between mitochondrial lineages. PLoS ONE 2018, 13, e0194045. [Google Scholar] [CrossRef]
- Karabanov, D.P.; Bekker, E.I.; Shiel, R.J.; Kotov, A.A. Invasion of a Holarctic planktonic cladoceran Daphnia galeata Sars (Crustacea: Cladocera) in the Lower Lakes of South Australia. Zootaxa 2018, 4402, 136–148. [Google Scholar] [CrossRef] [PubMed]
- Morais, P.; Reichard, M. Cryptic invasions: A review. Sci. Total Environ. 2018, 613–614, 1438–1448. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.J.; Ishikane, C.R.; Haney, R.A. The systematics of Holarctic bosminids and a revision that reconciles molecular and morphological evolution. Limnol. Oceanogr. 2002, 47, 1486–1495. [Google Scholar] [CrossRef] [Green Version]
- Kotov, A.A.; Taylor, D.J. Contrasting endemism in pond-dwelling cyclic parthenogens: The Daphnia curvirostris species group (Crustacea: Cladocera). Sci. Rep. 2019, 9, 6812. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Andujar, C.; Arribas, P.; Yu, D.W.; Vogler, A.P.; Emerson, B.C. Why the COI barcode should be the community DNA metabarcode for the metazoa. Mol. Ecol. 2018, 27, 3968–3975. [Google Scholar] [CrossRef]
- Coissac, E.; Hollingsworth, P.M.; Lavergne, S.; Taberlet, P. From barcodes to genomes: Extending the concept of DNA barcoding. Mol. Ecol. 2016, 25, 1423–1428. [Google Scholar] [CrossRef] [Green Version]
- Briski, E.; Cristescu, M.E.; Bailey, S.A.; MacIsaac, H.J. Use of DNA barcoding to detect invertebrate invasive species from diapausing eggs. Biol. Invasions 2011, 13, 1325–1340. [Google Scholar] [CrossRef]
- Comtet, T.; Sandionigi, A.; Viard, F.; Casiraghi, M. DNA (meta)barcoding of biological invasions: A powerful tool to elucidate invasion processes and help managing aliens. Biol. Invasions 2015, 17, 905–922. [Google Scholar] [CrossRef]
- Duggan, I.; Robinson, K.; Burns, C.; Banks, J.; Hogg, I. Identifying invertebrate invasions using morphological and molecular analyses: North American Daphnia ‘pulex’ in New Zealand fresh waters. Aquat. Invasions 2012, 7, 585–590. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Kotov, A.A. Establishment of Chydorus sphaericus (O.F. Muller, 1785) (Crustacea: Cladocera) in Australia: Consequences of mass fish stocking from Northern Europe? J. Limnol. 2015, 74, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Viard, F.; Riginos, C.; Bierne, N. Anthropogenic hybridization at sea: Three evolutionary questions relevant to invasive species management. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20190547. [Google Scholar] [CrossRef] [PubMed]
- Largiader, C.R. Hybridization and Introgression between Native and Alien Species. In Biological Invasions; Nentwig, W., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 275–292. ISBN 978-3-540-77375-7. [Google Scholar]
- Kodukhova, Y.V. Yearly variations of impact of natural hybrids of bream and roach (Abramis brama (L.) x Rutilus rutilus (L.)) in Rybinsk Reservoir. Russ. J. Biol. Invasions 2011, 2, 204–208. [Google Scholar] [CrossRef]
- Taylor, D.J.; Hebert, P.D.N. Cryptic intercontinental hybridization in Daphnia (Crustacea): The ghost of introductions past. Proc. R. Soc. Lond. B Biol. Sci. 1993, 254, 163–168. [Google Scholar] [CrossRef]
- Xu, S.; Innes, D.J.; Lynch, M.; Cristescu, M.E. The role of hybridization in the origin and spread of asexuality in Daphnia. Mol. Ecol. 2013, 22, 4549–4561. [Google Scholar] [CrossRef] [Green Version]
- Duffy, M.A.; Perry, L.J.; Kearns, C.M.; Weider, L.J.; Hairston, N.G. Paleogenetic evidence for a past invasion of Onondaga Lake, New York, by exotic Daphnia curvirostris using mtDNA from dormant eggs. Limnol. Oceanogr. 2000, 45, 1409–1414. [Google Scholar] [CrossRef] [Green Version]
- Nandini, S.; Silva-Briano, M.; García, G.G.; Sarma, S.S.S.; Adabache-Ortiz, A.; de La Rosa, R.G. First record of the temperate species Daphnia curvirostris Eylmann, 1887 emend. Johnson, 1952 (Cladocera: Daphniidae) in Mexico and its demographic characteristics in relation to algal food density. Limnology 2009, 10, 87–94. [Google Scholar] [CrossRef]
- Kotov, A.A.; Garibian, P.G.; Bekker, E.I.; Taylor, D.J.; Karabanov, D.P. A new species group from the Daphnia curvirostris species complex (Cladocera: Anomopoda) from the eastern Palaearctic: Taxonomy, phylogeny and phylogeography. Zool. J. Linn. Soc. 2021, 191, 772–822. [Google Scholar] [CrossRef]
- Kotov, A.A.; Ishida, S.; Taylor, D.J. A new species in the Daphnia curvirostris (Crustacea: Cladocera) complex from the eastern Palearctic with molecular phylogenetic evidence for the independent origin of neckteeth. J. Plankton Res. 2006, 28, 1067–1079. [Google Scholar] [CrossRef]
- Sinev, A.Y.; Karabanov, D.P.; Kotov, A.A. A new North Eurasian species of the Alona affinis complex (Cladocera: Chydoridae). Zootaxa 2020, 4767, 115–137. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [Green Version]
- Boratyn, G.M.; Camacho, C.; Cooper, P.S.; Coulouris, G.; Fong, A.; Ma, N.; Madden, T.L.; Matten, W.T.; McGinnis, S.D.; Merezhuk, Y.; et al. BLAST: A more efficient report with usability improvements. Nucleic Acids Res. 2013, 41, W29–W33. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, G. Estimating the dimension of a model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Xia, X. Nucleotide Substitution Models and Evolutionary Distances. In Bioinformatics and the Cell: Modern Computational Approaches in Genomics, Proteomics and Transcriptomics; Xuhua Xia, Ed.; Springer: Cham, Switzerland, 2018; pp. 269–314. ISBN 978-3-319-90684-3. [Google Scholar]
- Perlwitz, M.D.; Burks, C.; Waterman, M.S. Pattern analysis of the genetic code. Adv. Appl. Math. 1988, 9, 7–21. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Shimodaira, H. An approximately unbiased test of phylogenetic tree selection. Syst. Biol. 2002, 51, 492–508. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchene, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kuhnert, D.; de Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Collins, R.A.; Boykin, L.M.; Cruickshank, R.H.; Armstrong, K.F. Barcoding’s next top model: An evaluation of nucleotide substitution models for specimen identification. Methods Ecol. Evol. 2012, 3, 457–465. [Google Scholar] [CrossRef]
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar] [PubMed]
- Diniz-Filho, J.A.F.; Soares, T.N.; Lima, J.S.; Dobrovolski, R.; Landeiro, V.L.; de Campos Telles, M.P.; Rangel, T.F.; Bini, L.M. Mantel test in population genetics. Genet. Mol. Biol. 2013, 36, 475–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, M.S.; Anderson, C.D. PASSaGE: Pattern Analysis, Spatial Statistics and Geographic Exegesis. Version 2. Methods Ecol. Evol. 2011, 2, 229–232. [Google Scholar] [CrossRef]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000; ISBN 0195135857. [Google Scholar]
- Fu, Y.X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Artamonova, V.S.; Makhrov, A.A.; Karabanov, D.P.; Rolskiy, A.Y.; Bakay, Y.I.; Popov, V.I. Hybridization of beaked redfish (Sebastes mentella) with small redfish (Sebastes viviparus) and diversification of redfish (Actinopterygii: Scorpaeniformes) in the Irminger Sea. J. Nat. Hist. 2013, 47, 1791–1801. [Google Scholar] [CrossRef]
- Reuter, J.S.; Mathews, D.H. RNAstructure: Software for RNA secondary structure prediction and analysis. BMC Bioinform. 2010, 11, 129. [Google Scholar] [CrossRef] [Green Version]
- Karabanov, D.P.; Bekker, E.I.; Kotov, A.A. Underestimated consequences of biological invasions in phylogeographic reconstructions as seen in Daphnia magna (Crustacea, Cladocera). Zool. Zh. 2020, 99, 1232–1241. [Google Scholar] [CrossRef]
- Frey, D.G. Questions concerning cosmopolitanism in Cladocera. Arch. Hydrobiol. 1982, 93, 484–502. [Google Scholar]
- Van Damme, K.; Kotov, A.A. The fossil record of the Cladocera (Crustacea: Branchiopoda): Evidence and hypotheses. Earth-Sci. Rev. 2016, 163, 162–189. [Google Scholar] [CrossRef]
- Agar, W.E. The genetics of a Daphnia hybrid during parthenogenesis. J. Genet. 1920, 10, 303–330. [Google Scholar]
- Ishida, S.; Takahashi, A.; Matsushima, N.; Yokoyama, J.; Makino, W.; Urabe, J.; Kawata, M. The long-term consequences of hybridization between the two Daphnia species, D. galeata and D. dentifera, in mature habitats. BMC Evol. Biol. 2011, 11, 209. [Google Scholar] [CrossRef] [Green Version]
- Dlouha, S.; Thielsch, A.; Kraus, R.H.S.; Seda, J.; Schwenk, K.; Petrusek, A. Identifying hybridizing taxa within the Daphnia longispina species complex: A comparison of genetic methods and phenotypic approaches. Hydrobiologia 2010, 643, 107–122. [Google Scholar] [CrossRef]
- Benzie, J.A.H. The Genus Daphnia (including Daphniopsis): Anomopoda: Daphniidae; Kenobi Productions: Ghent, Belgium, 2005; ISBN 9057821516. [Google Scholar]
- Mordukhai-Boltovskoi, P.D. Caspian Polyphemids in the reservoirs of the Don and Dnieper Rivers. Tr. Inst. Biol. Vnutr. Vod AN SSSR 1965, 8, 37–43. [Google Scholar]
- Lazareva, V.I. Spreading of alien zooplankton species of Ponto-Caspian origin in the reservoirs of the Volga and Kama Rivers. Russ. J. Biol. Invasions 2019, 10, 328–348. [Google Scholar] [CrossRef]
- Korovchinsky, N.M. Cladocera: Ctenopoda: Families Sididae, Holopediidae & Pseudopenilidae (Branchiopoda: Cladocera); Backhuys Publishers, Margraf Publishers GmbH: Weikersheim, Germany, 2018; ISBN 978-3-8236-1756-3. [Google Scholar]
- Zhdanova, S.M. Diaphanosoma mongolianum Ueno, 1938 (Cladocera: Sididae) in Lakes of Yaroslavl Oblast (Russia). Inland Water Biol. 2018, 11, 145–152. [Google Scholar] [CrossRef]
- Reshetnikov, A.N. The current range of Amur sleeper Perccottus glenii Dybowski, 1877 (Odontobutidae, Pisces) in Eurasia. Russ. J. Biol. Invasions 2010, 1, 119–126. [Google Scholar] [CrossRef]
- Karabanov, D.P.; Kodukhova, Y.V.; Pashkov, A.N.; Reshetnikov, A.N.; Makhrov, A.A. “Journey to the West”: Three phylogenetic lineages contributed to the Invasion of Stone Moroko, Pseudorasbora parva (Actinopterygii: Cyprinidae). Russ. J. Biol. Invasions 2021, 12, 67–78. [Google Scholar] [CrossRef]
- Tyutin, A.V.; Verbitsky, V.B.; Verbitskaya, T.I.; Medyantseva, E.N. Parasites of alien aquatic animals in the upper Volga basin. Russ. J. Biol. Invasions 2013, 4, 54–59. [Google Scholar] [CrossRef]
- Zhokhov, A.E.; Pugacheva, M.N.; Molodozhnikova, N.M.; Berechikidze, I.A. Alien parasite species of the fish in the Volga River Basin: A review of data on the species number and distribution. Russ. J. Biol. Invasions 2019, 10, 136–152. [Google Scholar] [CrossRef]
- Voroshilova, I.S.; Pryanichnikova, E.G.; Prokin, A.A.; Sabitova, R.Z.; Karabanov, D.P.; Pavlov, D.D.; Kurina, E.M. Morphological and genetic traits of the first invasive population of the Asiatic Clam Corbicula fluminea (O.F. Müller, 1774) naturalized in the Volga River basin. Russ. J. Biol. Invasions 2021, 12, 36–43. [Google Scholar] [CrossRef]
- Shakirova, F. New records of the Chinese mitten crab, Eriocheir sinensis H. Milne Edwards, 1853, from the Volga River, Russia. Aquat. Invasions 2007, 2, 169–173. [Google Scholar] [CrossRef]
- Tishin, D.; Fardeeva, M.; Chizhikova, N.; Rizatdinov, R. Acclimation of Juglans mandshurica Maxim. and Phellodendron amurense Rupr. in the Middle Volga region. IOP Conf. Ser. Earth Environ. Sci. 2018, 107, 12094. [Google Scholar] [CrossRef] [Green Version]
- Dgebuadze, Y.Y.; Petrosyan, V.G.; Khlyap, L.A. (Eds.) The Most Dangerous Invasive Species in Russia (TOP-100); KMK Scientific press Ltd.: Moscow, Russia, 2018. [Google Scholar]
- Federal State Statistics Service. All-Russian Population Census—2010. Volume 1. Size and Distribution of the Population; Federal State Statistics Service: Moscow, Russia, 2011. [Google Scholar]
- Flossner, D. Zur Kenntnis einiger Daphnia-Hybriden (Crustacea: Cladocera). Limnologica 1993, 23, 71–79. [Google Scholar]
- Hobaek, A.; Skage, M.; Schwenk, K. Daphnia galeata x D. longispina hybrids in Western Norway. Hydrobiologia 2004, 526, 55–62. [Google Scholar] [CrossRef]
- Kotov, A.A. A critical review of the current taxonomy of the genus Daphnia O. F. Müller, 1785 (Anomopoda, Cladocera). Zootaxa 2015, 3911, 184–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Commission on Zoological Nomenclature. International Code of Zoological Nomenclature, 4th ed.; The Natural History Museum: London, UK, 2000. [Google Scholar]
- Kotov, A.A.; Forro, L.; Korovchinsky, N.M.; Petrusek, A. World Checklist of Freshwater Cladocera Species: World Wide Web Electronic Publication. Available online: http://fada.biodiversity.be/group/show/17 (accessed on 26 August 2021).







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karabanov, D.P.; Garibian, P.G.; Bekker, E.I.; Sabitova, R.Z.; Kotov, A.A. Genetic Signature of a Past Anthropogenic Transportation of a Far-Eastern Endemic Cladoceran (Crustacea: Daphniidae) to the Volga Basin. Water 2021, 13, 2589. https://doi.org/10.3390/w13182589
Karabanov DP, Garibian PG, Bekker EI, Sabitova RZ, Kotov AA. Genetic Signature of a Past Anthropogenic Transportation of a Far-Eastern Endemic Cladoceran (Crustacea: Daphniidae) to the Volga Basin. Water. 2021; 13(18):2589. https://doi.org/10.3390/w13182589
Chicago/Turabian StyleKarabanov, Dmitry P., Petr G. Garibian, Eugeniya I. Bekker, Rimma Z. Sabitova, and Alexey A. Kotov. 2021. "Genetic Signature of a Past Anthropogenic Transportation of a Far-Eastern Endemic Cladoceran (Crustacea: Daphniidae) to the Volga Basin" Water 13, no. 18: 2589. https://doi.org/10.3390/w13182589
APA StyleKarabanov, D. P., Garibian, P. G., Bekker, E. I., Sabitova, R. Z., & Kotov, A. A. (2021). Genetic Signature of a Past Anthropogenic Transportation of a Far-Eastern Endemic Cladoceran (Crustacea: Daphniidae) to the Volga Basin. Water, 13(18), 2589. https://doi.org/10.3390/w13182589