
Open Lake Phosphorus Forcing of Cladophora Growth: Modeling the Dual Challenge in Great Lakes Trophic State Management



Abstract
:1. Introduction
- With respect to efficacy, will reduction in local source phosphorus loading lead to measurable improvement if offshore forcing through cross-margin transport of SRP (for direct uptake by Cladophora) and PP (for conversion to SRP in the nearshore by dreissenids) have the capacity to sustain nuisance conditions of algal growth [11,14,15,16]?
- With respect to consequences, will reductions in phosphorus loading seeking mitigation of nuisance conditions in the nearshore promote oligotrophication and exacerbate the reductions in lake-wide ecosystem productivity presently observed to impact commercial, recreational, and native fish populations in the offshore? ([17]; see also [18], with respect to the Dual Challenge).
2. Objectives and Approach
- Would ambient nearshore SRP concentrations, determined solely by cross-margin transport of offshore SRP and PP (the latter subsequently mineralized to SRP in the nearshore by mussels), be sufficient to support nuisance growth of Cladophora?
3. Methods
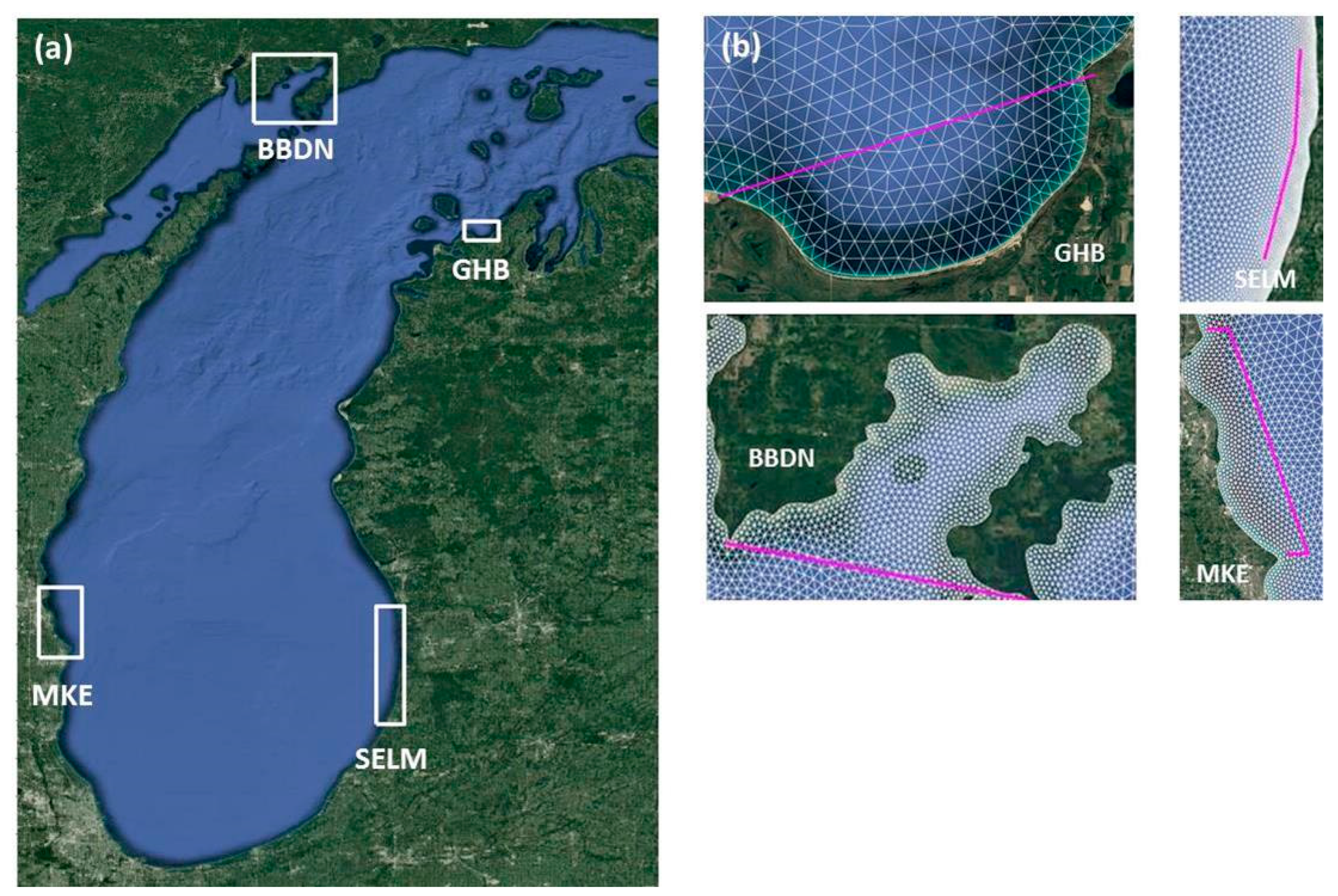
3.1. Study Site
3.2. Framework for Modeling the Mussel-Phosphorus Dynamic
- parameterization of the biokinetic coefficients Fvol and f (Section 3.2.1);
- specification of mussel biomass density (Section 3.1);
- specification of offshore PP and SRP concentrations (Section 3.2.2);
- specification of control volume physical characteristics (H and V; (Section 3.2.3)); and
- determination of the rate of cross margin flow (Q; Section 3.2.3).
3.2.1. Parameterization of the Biokinetic Coefficients Fvol and f
3.2.2. The Phosphorus Environment
3.2.3. The 3-D Hydrodynamic Model: Simulating Cross-Margin Transport
3.3. Design and Execution of Numerical Experiments
3.4. Interpreting the Results of Numerical Experiments
4. Results and Discussion
4.1. Sensitivity Analysis
4.1.1. Sensitivity to Biokinetic Coefficients
4.1.2. Sensitivity to the Rate of Cross-Margin Transport
4.1.3. Sensitivity to Offshore SRP Concentration
4.2. Management Analysis for Good Harbor Bay
4.3. Extending the Model Application
5. Cross-Margin Transport in a Management Context
- 1.
- In the absence of local sources of phosphorus, rates of Cladophora growth and biomass accrual are controlled by cross-margin transport of the nutrient from offshore to nearshore waters. The development of Cladophora beds and attendant beach accumulation will, in the absence of local sources, vary with the concentrations of SRP and PP in the open lake, i.e., the lake’s trophic status.
- 2.
- The potential for offshore stimulation of nuisance conditions on Lake Michigan was examined at Good Harbor Bay, a location absent local forcing. Simulations were performed for representative offshore concentrations of SRP and PP and other model inputs and coefficients. The results indicate that offshore forcing would not support SRP concentrations in the Good Harbor Bay nearshore sufficient to cause nuisance conditions. We posit that observations of accumulating algal debris in water adjoining Good Harbor Bay (e.g., Sleeping Bear Dunes National Lakeshore) reflects habitat expansion due to water-column clearing by mussels.
- 3.
- The analysis was extended beyond Good Harbor Bay to three additional locations on Lake Michigan to examine the sensitivity of nearshore SRP levels to variability in physical (cross-margin transport, flushing time) and biological (mussel biomass) forcing. Model projections indicate that nearshore SRP levels are insensitive to these factors for these four sites and that results evolving from analysis of conditions at Good Harbor Bay are applicable lake wide.
- The combined effect of reductions in external phosphorus loading to Lake Michigan and the benthification of paths of phosphorus recycling through mussel filtration of particulate phosphorus have driven offshore waters to a state of oligotrophy. This change has cascaded through the ecosystem, resulting in a reduction in prey species abundance and a decline in the number and condition of recreational and commercial fish species.
- In the broadest sense, it is the point and nonpoint discharges of phosphorus received in the nearshore that provide the foundation for a food web that can support the desired fishery. However, as those discharges remain resident in the nearshore for a period of time before being delivered to the offshore by cross-margin transport, they provide phosphorus enrichment that can stimulate nuisance growth of Cladophora.
- The conflicting nature of managing these two water-quality issues lies at the heart of what has been termed the Dual Challenge [18]. We refer to the two issues, Cladophora in the nearshore and the food web in the offshore, as endmembers for phosphorus-management scenarios. The phosphorus concentration required to eliminate nuisance Cladophora is the nearshore endmember and that supporting a healthy fishery is the offshore endmember. In this work, we applied a biophysical phosphorus–mussel model in establishing the nearshore endmember SRP concentration.
- As illustrated by the map of nearshore SRP (Figure 13), reductions in total phosphorus manifested in the transition from the pre- to post-dreissenid period are driven by changes in the offshore PP concentration (SRP increased modestly). If one assumes that pre-dreissenid production in the offshore food web was sufficient to support the fishery, then it is the difference in PP offshore levels between the pre- and post-dreissenid periods that must be “made up” in order to satisfy the objectives for the offshore endmember.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Study Sites on Lake Michigan


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Q (108 m3·d−1) | τ (d) | H (m) | Acs (105 m2) | Ab (105 m2) | V (109 m3) |
|---|---|---|---|---|---|---|
| Embayments | ||||||
| Good Harbor Bay | 2.10 | 4.47 | 10 | 2.79 | 430 | 0.934 |
| Big Bay De Noc | 2.19 | 15.3 | 7.63 | 2.51 | 4400 | 3.35 |
| Nearshore Regions | ||||||
| Milwaukee, WI | 4.70 | 5.44 | 13.03 | 7.60 | 1965 | 2.56 |
| Southeast Lake Michigan | 2.90 | 7.48 | 12.29 | 9.55 | 1770 | 2.17 |

Appendix B. The Lake Michigan Phosphorus Environment
| (a) Spring cruise, surface water (Spring Surface) concentrations of total, particulate, dissolved organic, and soluble reactive phosphorus for offshore sites on Lake Michigan over the entire period of record, 1983–2018 (n = 940). Data extracted from the U.S. EPA Great Lakes Water Quality Survey GLENDA database prior to partitioning with respect to region (north south; east west) or era (pre- and post-dreissenid periods). | ||||||||
| Mean (mgP·m−3) | S.D. (mgP·m−3) | Median (mgP·m−3) | ||||||
| TP | 4.17 | 1.44 | 4.03 | |||||
| PP | 1.86 | 1.20 | 1.75 | |||||
| DOP | 1.41 | 0.70 | 1.30 | |||||
| SRP | 0.90 | 0.49 | 0.82 | |||||
| (b) Spring cruise, surface water (Spring Surface) concentrations of total, particulate, dissolved organic, and soluble reactive phosphorus for offshore sites on Lake Michigan, partitioned into data for the pre-dreissenid (1983–1992; n = 339) and post-dreissenid (2009–2018; n = 244) periods. Data extracted from the U.S. EPA Great Lakes Water Quality Survey GLENDA database. | ||||||||
| Mean (mgP·m−3) | S.D. (mgP·m−3) | Median (mgP·m−3) | ||||||
| Pre | Post | Pre | Post | Pre | Post | |||
| TP | 5.02 | 3.06 | 1.56 | 0.76 | 4.80 | 3.03 | ||
| PP | 2.32 | 1.17 | 1.45 | 0.73 | 2.10 | 1.05 | ||
| DOP | 1.72 | 1.04 | 0.82 | 0.44 | 1.60 | 1.00 | ||
| SRP | 0.97 | 0.86 | 0.70 | 0.33 | 0.80 | 0.84 | ||
| (c) Concentrations of total, soluble reactive, and particulate phosphorus for nearshore (depth < 20 m) sites on Lake Michigan, partitioned into data for east (lower levels of watershed activity) and west (higher levels of watershed activity) stations. Data extracted from a monitoring survey performed in September 2010 by Yurista et al. [43] (Anne Cotter, U.S. EPA MED, personal communication). | ||||||||
| Nearshore East Stations | Nearshore West Stations | |||||||
| Mean (mgP·m−3) | S.D. (mgP·m−3) | n | Median (mgP·m−3) | Mean (mgP·m−3) | S.D. (mgP·m−3) | n | Median (mgP·m−3) | |
| TP | 7.87 | 2.28 | 8 | 7.53 | 11.15 | 4.77 | 6 | 10.56 |
| PP | 6.11 | 1.28 | 8 | 5.70 | 6.39 | 1.09 | 4 | 6.12 |
| SRP | 1.54 | 0.38 | 9 | 1.45 | 2.27 | 0.50 | 6 | 2.34 |
References
- Higgins, S.N.; Malkin, S.Y.; Howell, E.T.; Guildford, S.J.; Campbell, L.; Hiriart-Baer, V.; Hecky, R.E. An ecological review of Cladophora glomerata (Chlorophyta) in the Laurentian Great Lakes. J. Phycol. 2008, 44, 839–854. [Google Scholar] [CrossRef]
- Kuczynski, A.; Bakshi, A.; Auer, M.T.; Chapra, S.C. The canopy effect in filamentous algae: Improved modeling of Cladophora growth via a mechanistic representation of self-shading. Ecol. Model. 2020, 418, 108906. [Google Scholar] [CrossRef]
- Kuczynski, A.; Auer, M.T.; Brooks, C.N.; Grimm, A.G. The Cladophora resurgence in Lake Ontario: Characterization and implications for management. Can. J. Fish. Aquat. Sci. 2016, 73, 999–1013. [Google Scholar] [CrossRef] [Green Version]
- Chun, C.L.; Ochsner, U.; Byappanahalli, M.N.; Whitman, R.L.; Tepp, W.H.; Lin, G.; Johnson, E.A.; Peller, J.; Sadowsky, M.J. Association of toxin-producing Clostridium botulinum with the macroalga Cladophora in the Great Lakes. Environ. Sci. Technol. 2013, 47, 2587–2594. [Google Scholar] [CrossRef] [PubMed]
- Ishii, S.; Yan, T.; Shively, D.A.; Byappanahalli, M.N.; Whitman, R.L.; Sadowsky, M.J. Cladophora (Chlorophyta) spp. harbor human bacterial pathogens in nearshore water of Lake Michigan. Appl. Environ. Microbiol. 2006, 72, 4545–4553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auer, M.T.; Canale, R.P. Ecological studies and mathematical modeling of Cladophora in Lake Huron: 2. Phosphorus uptake kinetics. J. Gt. Lakes Res. 1982, 8, 84–92. [Google Scholar] [CrossRef]
- Higgins, S.N.; Pennuto, C.M.; Howell, E.T.; Lewis, T.W.; Makarewicz, J.C. Urban influences on Cladophora blooms in Lake Ontario. J. Gt. Lakes Res. 2012, 38, 116–123. [Google Scholar] [CrossRef]
- Hecky, R.E.; Smith, R.E.; Barton, D.R.; Guildford, S.J.; Taylor, W.D.; Charlton, M.N.; Howell, T. The nearshore phosphorus shunt: A consequence of ecosystem engineering by dreissenids in the Laurentian Great Lakes. Can. J. Fish. Aquat. Sci. 2004, 61, 1285–1293. [Google Scholar] [CrossRef]
- Dove, A.; Chapra, S.C. Long-term trends of nutrients and trophic response variables for the Great Lakes. Limnol. Oceanogr. 2015, 60, 696–721. [Google Scholar] [CrossRef]
- Painter, D.S.; Kamaitis, G. Reduction of Cladophora biomass and tissue phosphorus in Lake Ontario 1972–83. Can. J. Fish. Aquat. Sci. 1987, 44, 2212–2215. [Google Scholar] [CrossRef]
- Bootsma, H.A.; Rowe, M.D.; Brooks, C.N.; Vanderploeg, H.A. Commentary: The need for model development related to Cladophora and nutrient management in Lake Michigan. J. Gt. Lakes Res. 2015, 41, 7–15. [Google Scholar] [CrossRef]
- Auer, M.T.; McDonald, C.P.; Kuczynski, A.; Huang, C.; Xue, P. Management of the phosphorus–Cladophora dynamic at a site on lake ontario using a multi-module bioavailable P model. Water 2021, 13, 375. [Google Scholar] [CrossRef]
- Lambert, R.S.; Auer, M.T.; Effler, S.W.; Greene, M.R.; Downer, B.E.; Kuczynski, A. Onondaga to Ontario: Management of bioavailable phosphorus in municipal wastewaters for control of Cladophora. J. Gt. Lakes Res. 2015, 41, 1106–1113. [Google Scholar] [CrossRef]
- Bootsma, H.A.; Young, E.B.; Berges, J.A. Water quality management options to control Cladophora growth in the Milwaukee region of Lake Michigan. In Final Report for the Milwaukee Metropolitan Sewerage District; Univercity Wisconsin-Milwaukee: Milwaukee, WI, USA, 2008. [Google Scholar]
- Bootsma, H.A.; Waples, J.T.; Liao, Q. Identifying major phosphorus pathways in the Lake Michigan nearshore zone. In MMSD Contract M03029P05; Milwaukee Metropolitan Sewerage District: Milwaukee, WI, USA, 2012. [Google Scholar]
- Howell, E.T. Cladophora (green algae) and dreissenid mussels over a nutrient loading gradient on the north shore of Lake Ontario. J. Gt. Lakes Res. 2018, 44, 86–104. [Google Scholar] [CrossRef]
- Shen, C.; Liao, Q.; Bootsma, H.A. Modelling the influence of invasive mussels on phosphorus cycling in Lake Michigan. Ecol. Model. 2020, 416, 108920. [Google Scholar] [CrossRef]
- Hecky, R.E.; DePinto, J.V. Understanding Declining Productivity in the Offshore Regions of the Great Lakes; International Joint Commission: Windsor, ON, Canada, 2020; p. 62. [Google Scholar]
- Bravo, H.R.; Bootsma, H.; Khazaei, B. Fate of phosphorus from a point source in the Lake Michigan nearshore zone. J. Gt. Lakes Res. 2019, 45, 1182–1196. [Google Scholar] [CrossRef]
- Ozersky, T.; Malkin, S.Y.; Barton, D.R.; Hecky, R.E. Dreissenid phosphorus excretion can sustain C. glomerata growth along a portion of Lake Ontario shoreline. J. Gt. Lakes Res. 2009, 35, 321–328. [Google Scholar] [CrossRef]
- Dayton, A.I.; Auer, M.T.; Atkinson, J.F. Cladophora, mass transport, and the nearshore phosphorus shunt. J. Gt. Lakes Res. 2014, 40, 790–799. [Google Scholar] [CrossRef]
- Bootsma, H.A. Causes, consequences and management of nuisance Cladophora. In US Enviromental Protection Agency Project GL-00E06901; Great Lakes National Program Office: Chicago, IL, USA, 2009. [Google Scholar]
- Przybyla-Kelly, K.; Nevers, M.B.; Shively, D.A.; Benson, S.P.; Carter, G.M.; Dwyer, S.C.; Lewan, M.E.; Picard, K.R.; Richards, L.C.; Sopovski, D.S.; et al. Cladophora Biomass and Supporting Data Collected in the Great Lakes 2019; U.S. Geological Survey: Reston, VA, USA, 2019. [CrossRef]
- Nalepa, T.F.; Fanslow, D.L.; Lang, G.A.; Mabrey, K.; Rowe, M. Lake-Wide Benthic Surveys in Lake Michigan in 1994–95, 2000, 2005, and 2010: Abundances of the Amphipod Diporeia spp. and Abundances and Biomass of the Mussels Dreissena Polymorpha and Dreissena Rostriformis Bugensis; NOAA Technical MemorandumGLERL-164; NOAA, Great Lakes Environmental Research Laboratory: AnnArbor, MI, USA, 2014. [Google Scholar]
- Limno, T. Good Harbor Bay Dreissenid Mussel Control Demonstration Project Final Project Report; Great Lakes Commission and Invasive Mussel Collaborative Partners: Ann Arbor, MI, USA, 2020; p. 70. [Google Scholar]
- Nalepa, T.F.; Cavaletto, J.F.; Ford, M.; Gordon, W.M.; Wimmer, M. Seasonal and annual variation in weight and biochemical content of the zebra mussel, Dreissena polymorpha, in Lake St. Clair. J. Gt. Lakes Res. 1993, 19, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Glyshaw, P.W.; Riseng, C.M.; Nalepa, T.F.; Pothoven, S.A. Temporal trends in condition and reproduction of quagga mussels (Dreissena rostriformis bugensis) in southern Lake Michigan. J. Gt. Lakes Res. 2015, 41, 16–26. [Google Scholar] [CrossRef]
- Li, J.; Ianaiev, V.; Huff, A.; Zalusky, J.; Ozersky, T.; Katsev, S. Benthic invaders control the phosphorus cycle in the world’s largest freshwater ecosystem. Proc. Natl. Acad. Sci. USA 2021, 118, e2008223118. [Google Scholar] [CrossRef] [PubMed]
- Rowe, M.D.; Anderson, E.J.; Vanderploeg, H.A.; Pothoven, S.A.; Elgin, A.K.; Wang, J.; Yousef, F. Influence of invasive quagga mussels, phosphorus loads, and climate on spatial and temporal patterns of productivity in Lake Michigan: A biophysical modeling study. Limnol. Oceanogr. 2017, 62, 2629–2649. [Google Scholar] [CrossRef]
- Bierman, V.J., Jr.; Kaur, J.; DePinto, J.V.; Feist, T.J.; Dilks, D.W. Modeling the role of zebra mussels in the proliferation of blue-green algae in Saginaw Bay, Lake Huron. J. Gt. Lakes Res. 2005, 31, 32–55. [Google Scholar] [CrossRef]
- James, W.F.; Barko, J.W.; Davis, M.; Eakin, H.L.; Rogala, J.T.; Miller, A.C. Filtration and excretion by zebra mussels: Implications for water quality impacts in Lake Pepin, upper Mississippi River. J. Freshw. Ecol. 2000, 15, 429–437. [Google Scholar] [CrossRef]
- Turner, C.B. Influence of zebra (Dreissena polymorpha) and quagga (Dreissena rostriformis) mussel invasions on benthic nutrient and oxygen dynamics. Can. J. Fish. Aquat. Sci. 2010, 67, 1899–1908. [Google Scholar] [CrossRef]
- Mosley, C.; Bootsma, H. Phosphorus recycling by profunda quagga mussels (Dreissena rostriformis bugensis) in Lake Michigan. J. Gt. Lakes Res. 2015, 41, 38–48. [Google Scholar] [CrossRef]
- Vanderploeg, H.A.; Liebig, J.R.; Nalepa, T.F.; Fahnenstiel, G.L.; Pothoven, S.A. Dreissena and the disappearance of the spring phytoplankton bloom in Lake Michigan. J. Gt. Lakes Res. 2010, 36, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Fanslow, D.L.; Nalepa, T.F.; Lang, G.A. Filtration rates of the zebra mussel (Dreissena polymorpha) on natural seston from Saginaw Bay, Lake Huron. J. Gt. Lakes Res. 1995, 21, 489–500. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, B.S.; Mayer, M.S.; Dayton, J.; Pau, N.; Mendilla, J.; Sullivan, M.; Moore, A.; Ma, A.; Mills, E.L. Comparative growth and feeding in zebra and quagga mussels (Dreissena polymorpha and Dreissena bugensis): Implications for North American lakes. Can. J. Fish. Aquat. Sci. 2002, 59, 680–694. [Google Scholar] [CrossRef]
- Lei, J.; Payne, B.S.; Wang, S.Y. Filtration dynamics of the zebra mussel, Dreissena polymorpha. Can. J. Fish. Aquat. Sci. 1996, 53, 29–37. [Google Scholar] [CrossRef]
- Marescaux, J.; von Oheimb, K.C.; Etoundi, E.; von Oheimb, P.V.; Albrecht, C.; Wilke, T.; Van Doninck, K. Unravelling the invasion pathways of the quagga mussel (Dreissena rostriformis) into Western Europe. Biol. Invasions 2016, 18, 245–264. [Google Scholar] [CrossRef]
- Diggins, T.P. A seasonal comparison of suspended sediment filtration by quagga (Dreissena bugensis) and zebra (D. polymorpha) mussels. J. Gt. Lakes Res. 2001, 27, 457–466. [Google Scholar] [CrossRef]
- Xia, Z.; MacIsaac, H.J.; Hecky, R.E.; Depew, D.C.; Haffner, G.D.; Weidman, R.P. Multiple factors regulate filtration by invasive mussels: Implications for whole-lake ecosystems. Sci. Total Environ. 2021, 765, 144435. [Google Scholar] [CrossRef] [PubMed]
- Stoeckmann, A.M.; Garton, D.W. A seasonal energy budget for zebra mussels (Dreissena polymorpha) in western Lake Erie. Can. J. Fish. Aquat. Sci. 1997, 54, 2743–2751. [Google Scholar] [CrossRef]
- Pothoven, S.A.; Fahnenstiel, G.L. Recent change in summer chlorophyll a dynamics of southeastern Lake Michigan. J. Gt. Lakes Res. 2013, 39, 287–294. [Google Scholar] [CrossRef]
- Yurista, P.M.; Kelly, J.R.; Cotter, A.M.; Miller, S.E.; Van Alstine, J.D. Lake Michigan: Nearshore variability and a nearshore–offshore distinction in water quality. J. Gt. Lakes Res. 2015, 41, 111–122. [Google Scholar] [CrossRef]
- Huang, C.; Kuczynski, A.; Auer, M.T.; O’Donnell, D.M.; Xue, P. Management transition to the Great Lakes nearshore: Insights from hydrodynamic modeling. J. Mar. Sci. Eng. 2019, 7, 129. [Google Scholar] [CrossRef] [Green Version]
- Kelley, J.G.W.; Chen, Y.; Anderson, E.J.; Lang, G.A.; Peng, M. Upgrade of NOS Lake Michigan and Lake Huron Operational Forecast Systems to FVCOM: Model Development and Hindcast Skill Assessment; NOAA Technical Memorandum NOS CS, 42; National Ocean Service Coast Survey Development Laboratory: Silver Spring, MD, USA, 2020.
- Benjamin, S.G.; Weygandt, S.S.; Brown, J.M.; Hu, M.; Alexander, C.R.; Smirnova, T.G.; Olson, J.B.; James, E.P.; Dowell, D.C.; Grell, G.A.; et al. A North American hourly assimilation and model forecast cycle: The rapid refresh. Mon. Weather Rev. 2016, 144, 1669–1694. [Google Scholar] [CrossRef]
- Peng, M.; Zhang, A.; Anderson, E.J.; Lang, G.A.; Kelley, J.G.W.; Chen, Y. Implementation of the Lakes Michigan and Huron Operational Forecast System (LMHOFS) and the Nowcast/Forecast Skill Assessment; NOAA Technical Report NOS CO-OPS, 091; National Ocean Service Center for Operational Oceangraphic Products and Services: Silver Spring, MD, USA, 2019.
- Auer, M.T.; Canale, R.P. Ecological studies and mathematical modeling of Cladophora in Lake Huron: 3. The dependence of growth rates on internal phosphorus pool size. J. Gt. Lakes Res. 1982, 8, 93–99. [Google Scholar] [CrossRef]
- Tomlinson, L.M.; Auer, M.T.; Bootsma, H.A.; Owens, E.M. The Great Lakes Cladophora model: Development, testing, and application to Lake Michigan. J. Gt. Lakes Res. 2010, 36, 287–297. [Google Scholar] [CrossRef]
- Kuczynski, A.; Auer, M.T.; Taylor, W.D.; Chapra, S.C. Keeping up with the math: Advancing the ecological foundation of the Great Lakes Cladophora Model. Ecol. Model. In Review.
- Waples, J.T.; Klump, J.V. Vertical and horizontal particle transport in the coastal waters of a large lake: An assessment by sediment trap and thorium-234 measurements. J. Geophys. Res. Ocean. 2013, 118, 5376–5397. [Google Scholar] [CrossRef]
- Waples, J.T.; Bootsma, H.A.; Klump, J.V. How are coastal benthos fed? Limnol. Oceanogr. Lett. 2017, 2, 18–28. [Google Scholar] [CrossRef]
- Xue, P.; Schwab, D.J.; Hu, S. An investigation of the thermal response to meteorological forcing in a hydrodynamic model of Lake Superior. J. Geophys. Res. Ocean. 2015, 120, 5233–5253. [Google Scholar] [CrossRef]
- Xue, P.; Pal, J.S.; Ye, X.; Lenters, J.D.; Huang, C.; Chu, P.Y. Improving the simulation of large lakes in regional climate modeling: Two-way lake–atmosphere coupling with a 3D hydrodynamic model of the Great Lakes. J. Clim. 2017, 30, 1605–1627. [Google Scholar] [CrossRef]
- Ye, X.; Anderson, E.J.; Chu, P.Y.; Huang, C.; Xue, P. Impact of water mixing and ice formation on the warming of Lake Superior: A model-guided mechanism study. Limnol. Oceanogr. 2019, 64, 558–574. [Google Scholar] [CrossRef]
- Ye, X.; Chu, P.Y.; Anderson, E.J.; Huang, C.; Lang, G.A.; Xue, P. Improved thermal structure simulation and optimized sampling strategy for Lake Erie using a data assimilative model. J. Gt. Lakes Res. 2020, 46, 144–158. [Google Scholar] [CrossRef]













| Model Input or Coefficient | Value | Range |
|---|---|---|
| (a). Model inputs and coefficients for Good Harbor Bay, post-dreissenid period | ||
| Mussel biomass density (B, mgAFDW·m−2) | 15,266 | 0–50,000 |
| Mussel filtration rate (Fvol, ×10−4 m3·mgAFDW−1·d−1) | 4.75 | 3.04, 4.75, 6.46 |
| Mussel PP to SRP conversion efficiency (f, d’less) | 0.29 | 0.2, 0.3, 0.4, 0.5 |
| Rate of cross-margin flow (Q, ×108 m3·d−1) | 2.1 | 0.25, 1.0, 2.0,4.0, 6.0 |
| (b). Phosphorus levels for the Lake Michigan offshore, post–dreissenid period | ||
| SRP in the offshore, SRPoffshore, mgP·m−3 | 0.84 | 0.50, 0.80, 1.00, 1.25, 1.50 |
| PP in the offshore, PPoffshore, mgP·m−3 | 1.05 | 0–7 |
| (c). Phosphorus levels for the Lake Michigan offshore, pre-dreissenid period | ||
| SRP in offshore waters (SRPoffshore, mgP·m−3) | 0.80 | |
| PP in offshore waters, (PPoffshore, mgP·m−3) | 2.10 | |
| (d). Phosphorus levels for the Lake Michigan nearshore, post-dreissenid period | ||
| SRP at nearshore east stations, SRPnearshore, mgP·m−3 | 1.45 | |
| SRP at nearshore west stations, SRPnearshore, mgP·m−3 | 2.34 | |
| PP at nearshore east stations, PPnearshore, mgP·m−3 | 5.70 | |
| PP at nearshore west stations, PPnearshore, mgP·m−3 | 6.12 | |
| Site on Lake Michigan | H (m) | τ (Days) | B Contemporary (mgAFDW·m−2) | α Contemporary (Dimensionless) | B No Local P Sources (mgAFDW·m−2) | α No Local P Sources (Dimensionless) |
|---|---|---|---|---|---|---|
| Good Harbor Bay | 10 | 4.47 | 15,266 [21,23,25] | 3.24 | 10,069 | 2.13 |
| Milwaukee, WI nearshore region | 13.03 | 5.44 | 98,560 [15] | 19.55 | 10,069 | 2.00 |
| Big Bay De Noc | 7.63 | 15.3 | 4510 [24] | 4.29 | 10,069 | 9.59 |
| Southeast nearshore region | 12.29 | 7.48 | 10,431 [24] | 3.01 | 10,069 | 2.91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Auer, M.T.; Xue, P. Open Lake Phosphorus Forcing of Cladophora Growth: Modeling the Dual Challenge in Great Lakes Trophic State Management. Water 2021, 13, 2680. https://doi.org/10.3390/w13192680
Zhou X, Auer MT, Xue P. Open Lake Phosphorus Forcing of Cladophora Growth: Modeling the Dual Challenge in Great Lakes Trophic State Management. Water. 2021; 13(19):2680. https://doi.org/10.3390/w13192680
Chicago/Turabian StyleZhou, Xing, Martin T. Auer, and Pengfei Xue. 2021. "Open Lake Phosphorus Forcing of Cladophora Growth: Modeling the Dual Challenge in Great Lakes Trophic State Management" Water 13, no. 19: 2680. https://doi.org/10.3390/w13192680
APA StyleZhou, X., Auer, M. T., & Xue, P. (2021). Open Lake Phosphorus Forcing of Cladophora Growth: Modeling the Dual Challenge in Great Lakes Trophic State Management. Water, 13(19), 2680. https://doi.org/10.3390/w13192680