
Three New Species of Placoneis Mereschkowsky (Bacillariophyceae: Cymbellales) with Comments on Cryptic Diversity in the P. elginensis—Group

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
Molecular Methods
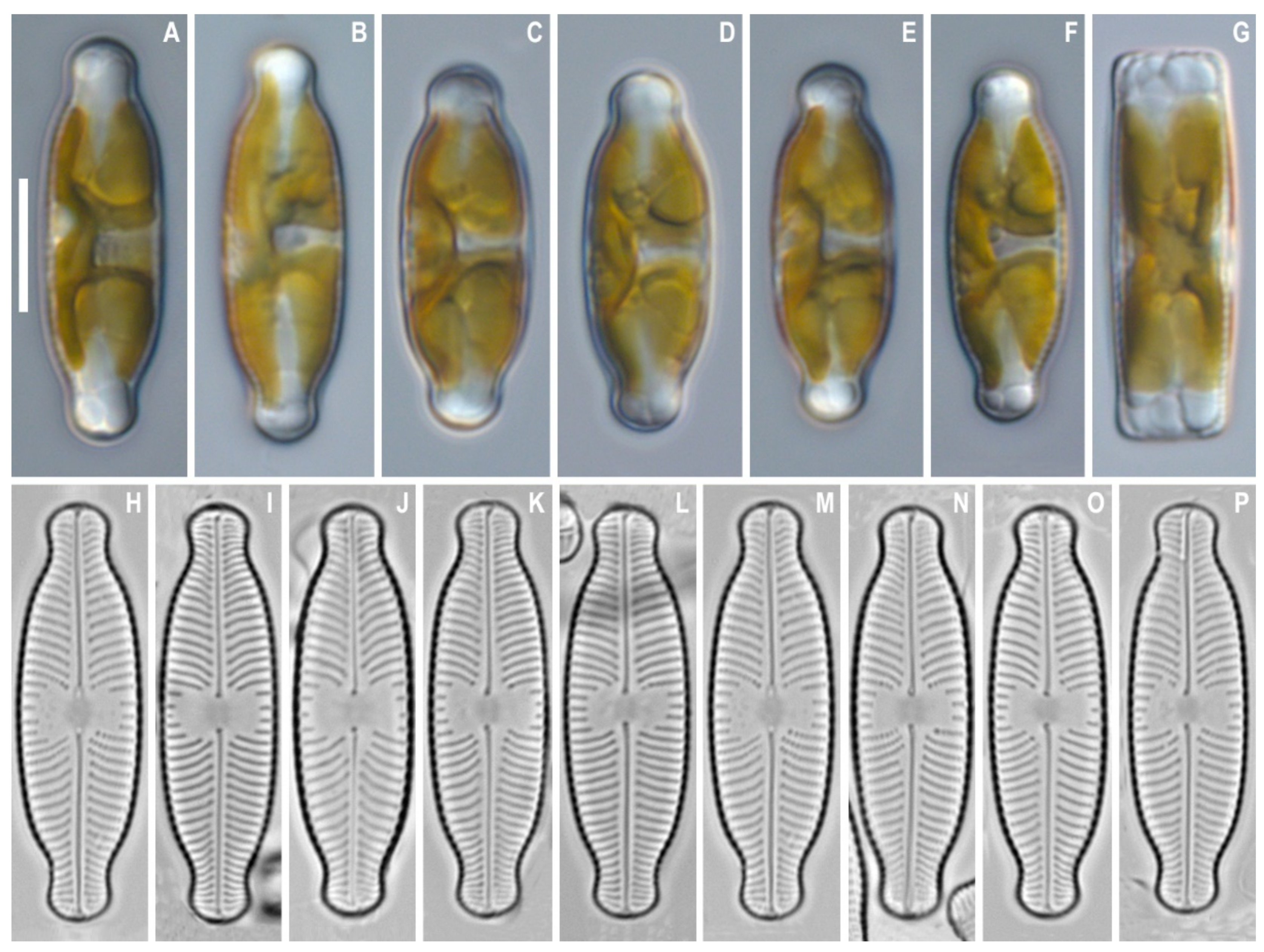
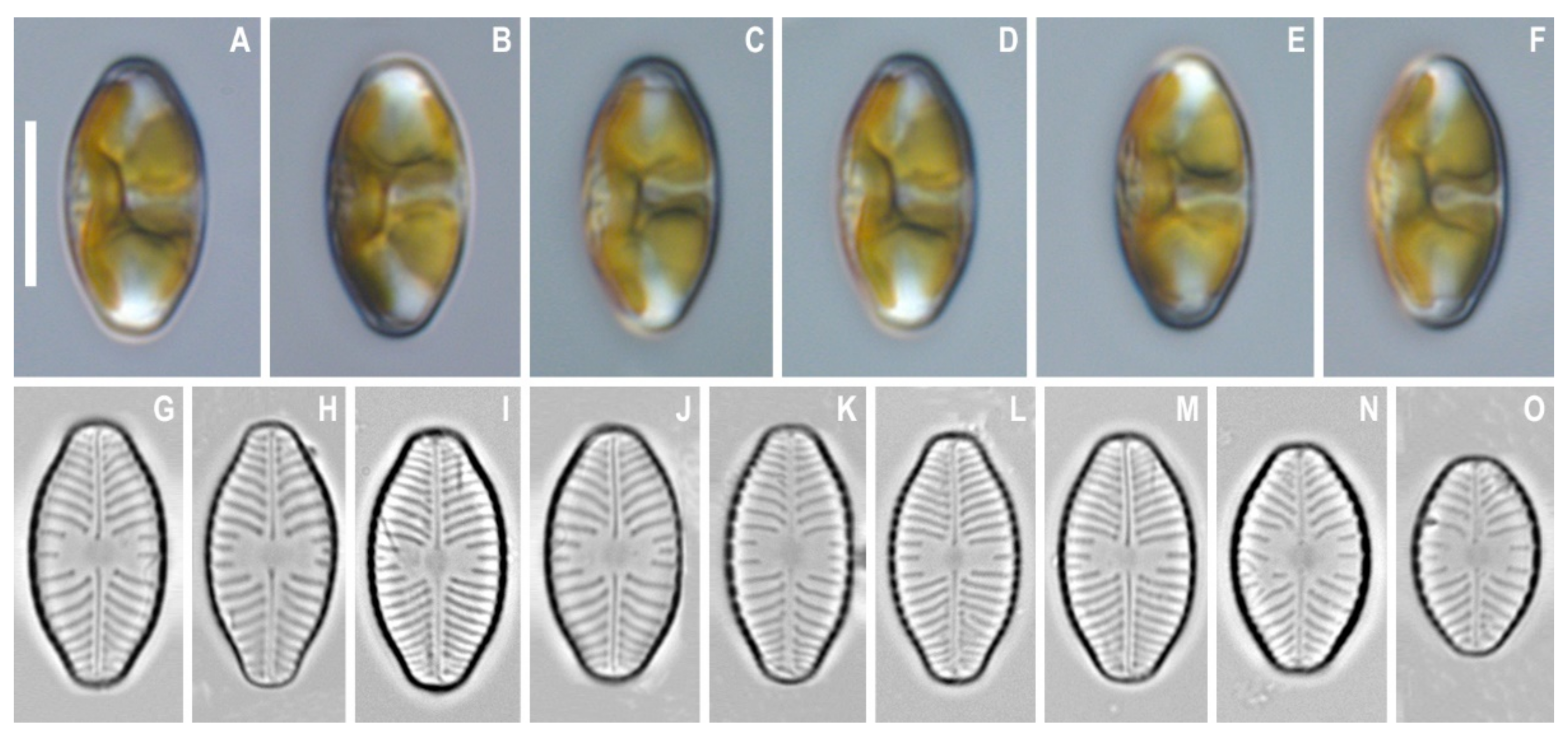
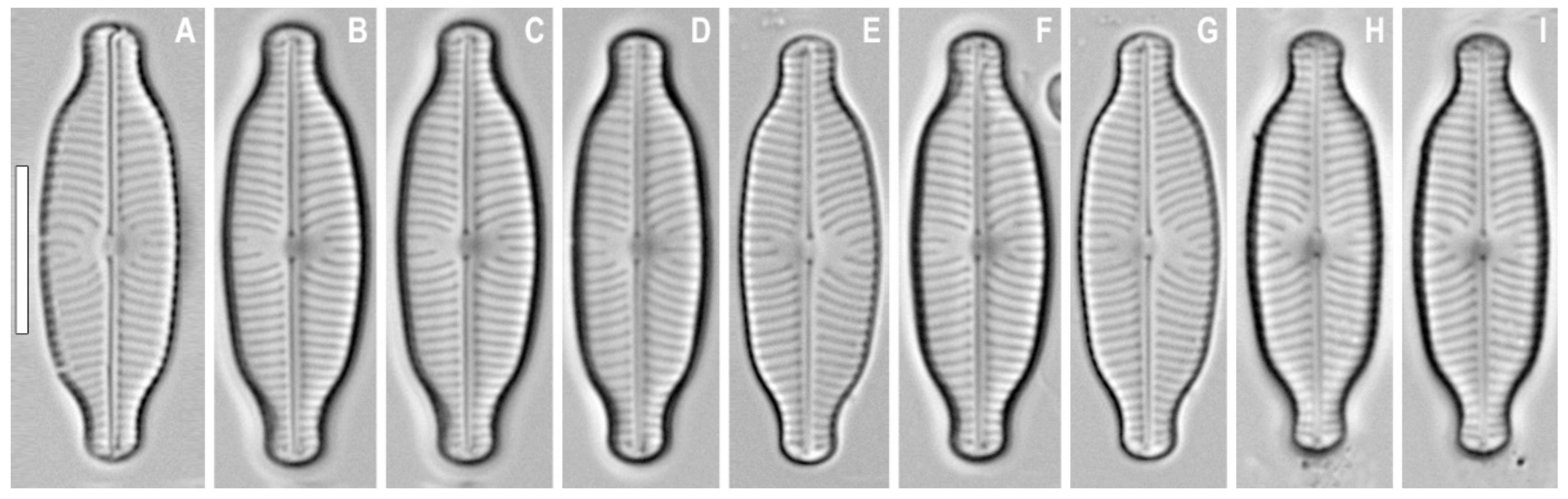
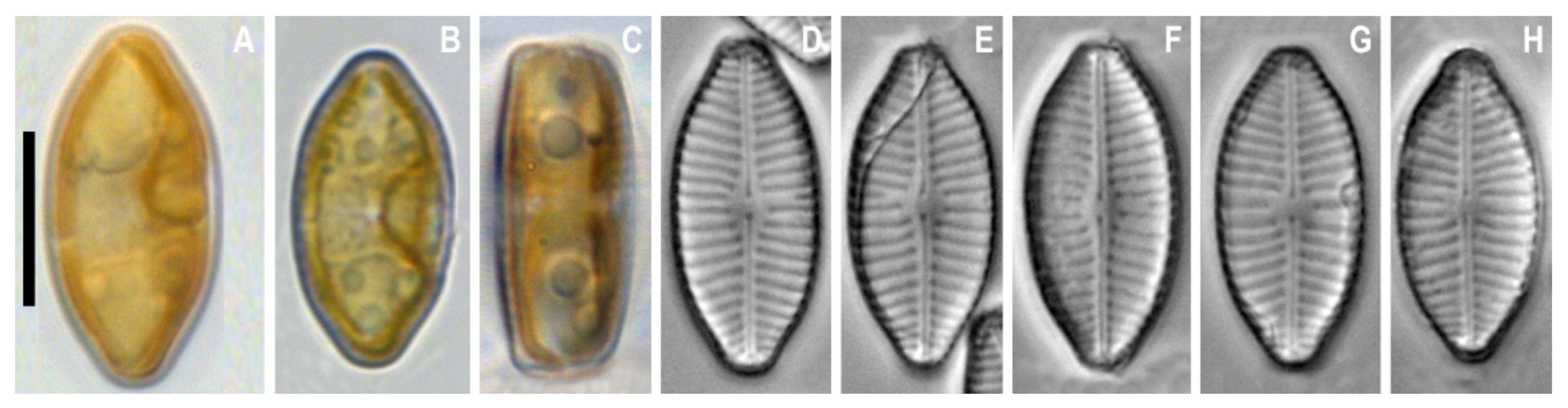
3. Results
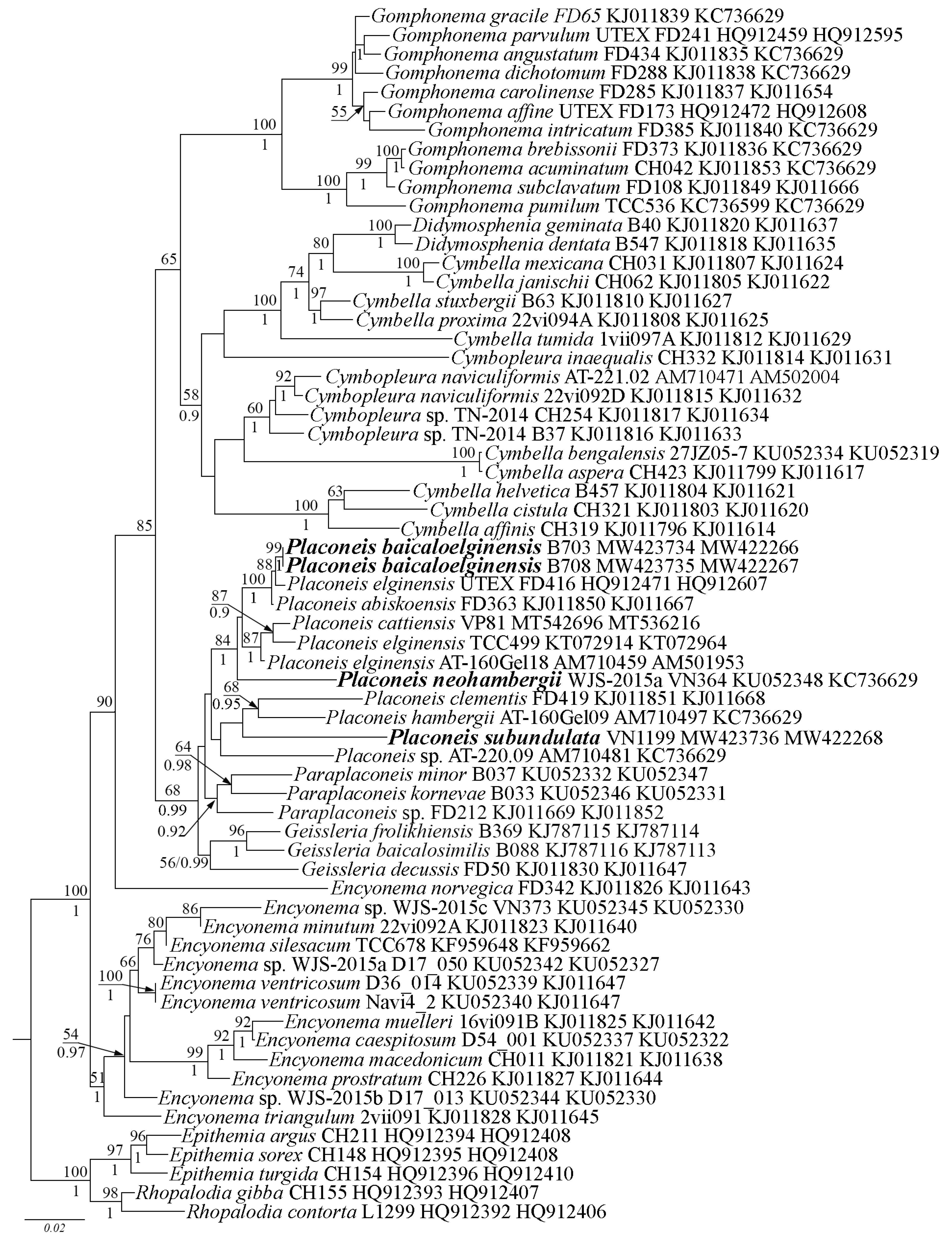
Molecular Investigation
4. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. subundulata sp. nov. | P. undulata | P. cattiensis | P. ignorata | P. neohambergii sp. nov. | P. hambergii | P. opportuna | P. witkowskii | P. ovilus | |
|---|---|---|---|---|---|---|---|---|---|
| Outline | Linear-elliptical, with slightly triundulate valve margins | Elliptic-undulate | Linear-elliptical | Linear-elliptical to elliptic-lanceolate | elliptical- lanceolate | Elliptic-lanceolate | Elliptic-lanceolate to elliptic | Elliptic | Strictly elliptic |
| Apex shape | Rostrate to subcapitate | Rostrate | subcapitate | Rostrate to broadly rounded | barely protracted broadly rounded | Slightly or distinct rostrate | Broadly rounded | Broadly protracted subrostrate | Slightly cuneate |
| Apex width (μm) | 3–3.5 | 2.5 | 2.5–3 | 2.8–3 * | 2–3 | 2–2.5 | - | - | - |
| Axial area | Narrow, linear | Narrow, slightly widening to the middle of the valve | Narrow, slightly widening to the middle of the valve | Narrow, linear | Narrow, linear | Linear—lanceolate | Narrow, linear | Narrow, linear | Narrow, slightly expanded towards the center |
| Central area | Transversely-expanded, asymmetrical, rounded or butterfly-shaped confined by 3–4 shortened striae from ¼ to ½ width of valve | Transverse elliptic, confined by 3–4 shortened striae to ½ width of valve | Large, transverse, rarely asymmetrical, from ½ to ⅔ width of valve | Transversely-expanded, rectangular, confined by 2 shortened striae | Small, rounded or not distinct, confined one or two shorter striae | Not expressed, confined by two shortened striae | Small, weakly expressed, confined by shortened striae | Small, ill-defined in outline by single longer stria in the middle and two shorter striae on either side | Small, irregularly confined by a few alternating longer and shorter striae |
| Valve length (μm) | 25.5–27 | 18–19 | 24–25 | 12–25 | 17–19 | 16–25 | 7.9–14.4 (20) | 14–24 | 18–23 |
| Valve breadth (μm) | 7.3–8 | About 7 | 6.7–7.4 | 7–8 | 7.5–8 | 6–8 | 5.4–7.3 (8) | 8–10 | 8.6–9.3 |
| Striae type, number in 10 μm | Uniseriate, radiate becoming subparallel in the ends,14–15 | Uniseriate, radiate,12 | Uniseriate, radiate becoming subparallel in the ends,12–14 | Uniseriate, radiate, 11–14 | Uniseriate, slightly radiate or subparallel,12–14 | Uniseriate, radiate,15–18 | Uniseriate, radiate,15.1–16.6 (18) | Uniseriate, radiate throughout,12–15 | Uniseriate, radiate throughout and somewhat curved,12–13.5 |
| Areolae, number in 10 μm | Indiscernible in LM, 35 | Indiscernible in LM | Not discernible in the LM, 45–50 | Not discernible in the LM | Indiscernible in LM, 35 | Indiscernible in LM, 45* | Indiscernible in LM, 40–42.7 (45.7) | Not to discern in the LM, (mach more 30) | Discernible in the LM, 27 |
| Distribution | Indonesia | North Tirol, Austria | Vietnam | Widespread species | Vietnam | Holarctic | Widespread species | Laguna Blanca, Department of Maldonado, Uruguay | Arroyo Sause, Arroyo del Aiguá, Department of Maldonado, Uruguay |
| References | This study | [5,20,36,49] | [50] | [20,33,36,42,44] | This study | [10,33,48] | [33,45] | [48] | [48] |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bory, J.B.G.M. Navicula, N. V. pl. de ce Dict., Bacillariées, fig. 5. In Dictionnaire Classique d’Histoire Naturelle 2; Rey et Gravier and Baudoin Frères: Paris, France, 1822; Volume 2, pp. 127–129. [Google Scholar]
- Mereschkowsky, C. Über Placoneis, ein neues Diatomeen-Genus. Beih. Zum Bot. Cent. 1903, 15, 30. [Google Scholar]
- Hustedt, F. Bacillariophyta (Diatomeae) zweite auflage. In Die Süβwasser-Flora Mitteleuropas; Pascher, A., Ed.; Verlag von Gustav Fischer: Jena, Germany, 1930; pp. 1–466. [Google Scholar]
- Patrick, R.; Reimer, C.W. The Diatoms of the United States (Exclusive of Alaska and Hawaii), Volume 1. Chesap. Sci. 1966, 7, 115. [Google Scholar] [CrossRef]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. 1. Teil: Naviculaceae. In Süsswasserflora von Mitteleuropa; Ettl, H.J., Gerloff, H., Heynig, H., Mollenhauer, D., Eds.; Gustav Fisher Verlag: Jena, Germany, 1986; Volume 2/1, 876p. [Google Scholar]
- Cox, E.J. Placoneis Mereschkowsky: The re-evaluation of a diatom genus originally characterized by its chloroplast type. Diatomic Res. 1987, 2, 145–157. [Google Scholar] [CrossRef]
- Cox, E.J. Pore occlusions in raphid diatoms—A reassessment of their structure and terminology, with particular reference to members of the Cymbelales. Diatom 2004, 20, 33–46. [Google Scholar]
- Kulikovskiy, M.S.; Lange-Bertalot, H.; Metzeltin, D.; Witkowski, A. Lake Baikal: Hotspot of endemic diatoms I. Iconogr. Diatomol. 2012, 23, 7–608. [Google Scholar]
- Fujita, Y.; Ohtsuka, T. Diatoms from paddy fields in northern Laos. Diatom 2005, 21, 71–89. [Google Scholar]
- Bruder, K.; Medlin, L.K. Molecular assessment of phylogenetic relationships in selected species/genera in the naviculoid diatoms (Bacillariophyta). I. The genus Placoneis. Nova Hedwig. 2007, 85, 331–352. [Google Scholar] [CrossRef]
- Mann, D.G.; McDonald, S.M.; Bayer, M.M.; Droop, S.J.M.; Chepurnov, V.A.; Loke, R.E.; Ciobanu, A.; Du Buf, J.M.H. The Sellaphora pupula species complex (Bacillariophyceae): Morphometric analysis, ultrastructure and mating data provide evidence for five new species. Phycologia 2004, 43, 459–482. [Google Scholar] [CrossRef] [Green Version]
- Evans, K.M.; Wortley, A.H.; Simpson, G.E.; Chepurnov, V.A.; Mann, D.G. A molecular systematic approach to explore diversity within the Sellaphora pupula species complex (Bacillariophyta). J. Phycol. 2008, 44, 215–231. [Google Scholar] [CrossRef]
- Trobajo, R.; Clavero, E.; Chepurnov, V.A.; Sabbe, K.; Mann, D.G.; Ishihara, S.; Cox, E.J. Morphological, genetic, and mating diversity within the widespread bioindicator Nitzschia palea (Bacillariophyceae). Phycologia 2009, 48, 443–459. [Google Scholar] [CrossRef]
- Poulíčková, A.; Veselá, J.; Neustupa, J.; Škaloud, P. Pseudocryptic diversity versus cosmopolitanism in diatoms: A case study Navicula cryptocephala Kütz. (Bacillariophyceae) and morfologically similar taxa. Protist 2010, 161, 353–369. [Google Scholar] [CrossRef]
- Abarca, N.; Jahn, R.; Zimmermann, J.; Enke, N. Does the cosmopolitan diatom Gomphonema parvulum (Kützing) Kützing have a biogeography? PLoS ONE 2014, 9, e86885. [Google Scholar] [CrossRef]
- Pinseel, E.; Kulichová, J.; Scharfen, V.; Urbánková, P.; Van de Vijver, B.; Vyverman, W. Extensive Cryptic Diversity in the Terrestrial Diatom Pinnularia borealis (Bacillariophyceae). Protist 2018, 170, 121–140. [Google Scholar] [CrossRef]
- Kollár, J.; Pinseel, E.; Vanormelingen, P.; Poulíčková, A.; Souffreau, C.; Dvořák, P.; Vyverman, W. A polyphasic approach to the delimitation of diatom species: A case study for the genus Pinnularia (Bacillariophyta). J. Phycol. 2018, 55, 365–379. [Google Scholar] [CrossRef]
- Kollár, J.; Pinseel, E.; Vyverman, W.; Poulíčková, A. A time-calibrated multi-gene phylogeny provides insights into the evolution, taxonomy and DNA barcoding of the Pinnularia gibba group (Bacillariophyta). Fottea 2021, 21, 62–72. [Google Scholar] [CrossRef]
- Maltsev, Y.; Maltseva, S.; Kociolek, J.P.; Jahn, R.; Kulikovskiy, M. Biogeography of the cosmopolitan terrestrial diatom Hantzschia amphioxys sensu lato based on molecular and morphological data. Sci. Rep. 2021, 11, 4266. [Google Scholar] [CrossRef]
- Zimmermann, J.; Jahn, R.; Gemeinholzer, B. Barcoding diatoms: Evaluation of the V4 subregion on the 18S rRNA gene, including new primers and protocols. Org. Divers. Evol. 2011, 11, 173–192. [Google Scholar] [CrossRef]
- Ruck, E.C.; Theriot, E.C. Origin and Evolution of the Canal Raphe System in Diatoms. Protist 2011, 162, 723–737. [Google Scholar] [CrossRef]
- Alverson, A.J.; Jansen, R.K.; Theriot, E.C. Bridging the Rubicon: Phylogenetic analysis reveals repeated colonizations of marine and fresh waters by thalassiosiroid diatoms. Mol. Phylogenet. Evol. 2007, 45, 193–210. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Toh, H. Parallelization of the MAFFT multiple sequence alignment program. Bioinformatics 2010, 26, 1899–1900. [Google Scholar] [CrossRef]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A Rapid Bootstrap Algorithm for the RAxML Web Servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Nakov, T.; Ruck, E.C.; Galachyants, Y.; Spaulding, S.A.; Theriot, E.C. Molecular phylogeny of the Cymbellales (Bacillariophyceae, Heterokontophyta) with a comparison of models for accommodating rate variation across sites. Phycologia 2014, 53, 359–373. [Google Scholar] [CrossRef]
- Thomas, E.W.; Stepanek, J.G.; Kociolek, J.P. Historical and Current Perspectives on the Systematics of the ‘Enigmatic’ Diatom Genus Rhoicosphenia (Bacillariophyta), with Single and Multi-Molecular Marker and Morphological Analyses and Discussion on the Monophyly of ‘Monoraphid’ Diatoms. PLoS ONE 2016, 11, e0152797. [Google Scholar] [CrossRef]
- Kociolek, J.P.; Stoermer, E.F. A Preliminary Investigation of the Phylogenetic Relationships among the Freshwater, Apical Pore Field-Bearing Cymbelloid and Gomphonemoid Diatoms (Bacillariophyceae). J. Phycol. 1988, 24, 377–385. [Google Scholar] [CrossRef]
- Krammer, K. Cymbopleura, Delicata, Navicymbula, Gomphocymbellopsis, Afrocymbella. Diatoms Eur. 2003, 4, 1–530. [Google Scholar]
- Kociolek, J.P.; Blanco, S.; Coste, M.; Ector, L.; Liu, Y.; Karthick, B.; Kulikovskiy, M.; Lundholm, N.; Ludwig, T.; Potapova, M.; et al. DiatomBase. 2021. Available online: https://www.diatombase.org (accessed on 22 September 2021).
- Kulikovskiy, M.S.; Glushchenko, A.M.; Genkal, S.I.; Kuznetsova, I.V. Identification Book of Diatoms from Russia; Filigran: Yaroslavl, Russia, 2016; p. 804. (In Russian) [Google Scholar]
- Kociolek, J.P. A worldwide listing and biogeography of freshwater diatom genera: A phylogenetic perspective. Diatom Res. 2018, 33, 509–534. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, A. A History of Infusoria, Including the Desmidiaceae and Diatomaceae, British and Foreign, 4th ed.; Archer, W., Arlidge, J.T., Ralfs, J., Williamson, W.C., Eds.; Whittaker and Co.: London, UK, 1861; Volume XII, 968p. [Google Scholar]
- Cox, E.J. Placoneis Mereschkowsky revisited: Resolution of several typification and nomenclatural problems, including the generitype. Bot. J. Linn. Soc. 2003, 141, 53–83. [Google Scholar] [CrossRef]
- Lange-Bertalot, H.; Kulbs, K.; Lauser, T.; Norpel-Schempp, M.; Willmann, M. Diatom taxa introduced by Georg Krasske: Documentation and revision. Iconogr. Diatomol. 1996, 3, 1–358. [Google Scholar]
- Rumrich, U.; Lange-Bertalot, H.; Rumrich, M. Diatoms of the Andes. From Venezuela to Patagonia/Tierra del Fuego and two additional contributions. Iconogr. Diatomol. 2000, 9, 1–673. [Google Scholar]
- Jahn, R.; Kusber, W.-H. AlgaTerra Information System. Botanic Garden and Botanical Museum Berlin-Dahlem, Freie Universität Berlin. Available online: https://www.algaterra.org (accessed on 20 September 2021).
- Rimet, F.; Chardon, C.; Lainé, L.; Bouchez, A.; Domaizon, I.; Guillard, J.; Jacquet, S. Thonon Culture Collection—TCC—A Freshwater Microalgae Collection. Portail Data Inra, V1. Available online: https://doi.org/10.15454/UQEMVW (accessed on 20 September 2021).
- Rimet, F. Photos of the TCC Cultures and Samples. Zenodo. Available online: http://doi.org/10.5281/zenodo.2545613(accessed on 20 September 2021).
- Reichardt, E. Die Diatomeen im Gebiet der Stadt Treuchtlingen, 3rd ed.; Bayerische Botanische Gesellschaft: Munich, Germany, 2018; Volumes 1–2, pp. 1–576, 579–1184. [Google Scholar]
- Metzeltin, D.; Witkowski, A. Diatomeen der Bären-Insel. Süsswasser- und marine Arten. Iconogr. Diatomol. 1996, 4, 1–232. [Google Scholar]
- Cantonati, M.; Kelly, M.G.; Lange-Bertalot, H. Freshwater Benthic Diatoms of Central Europe: Over 800 Common Species Used in Ecological Assessment, 3rd ed.; Koeltz Botanical Books: Oberreifenberg, Germany, 2017; 942p. [Google Scholar]
- Chudaev, D.A.; Gololobova, M.A. Diatoms of the Glubokoe Lake (Moscow Region), 3rd ed.; KMK: Moscow, Russia, 2016; p. 446. (In Russian) [Google Scholar]
- Miho, A.; Lange-Bertalot, H. Diversity of the genus Placoneis in Lake Ohrid and other freshwater habitats of Albania. In Proceedings of the 18th International Diatom Symposium, Miedzyzdroje, Poland, 2–7 September 2006; pp. 301–313. [Google Scholar]
- Genkal, S.I.; Kulikovskiy, M.S.; Mikheeva, T.M.; Kuznetsova, I.V.; Lukyanova, E.V. Diatoms in Plankton of the Svisloch River and Its Reservoirs; Sientific World: Moscow, Russia, 2013; p. 236. (In Russian) [Google Scholar]
- Metzeltin, D.; Lange-Bertalot, H.; García-Rodrígues, F. Diatoms of Uruguay. Compared with other taxa from South America and elsewhere. Iconogr. Diatomol. 2005, 15, 1–736. [Google Scholar]
- Ohtsuka, T.; Fujita, Y. The diatom flora and its seasonal changes in a paddy field in Central Japan. Nova Hedwig. 2001, 73, 97–128. [Google Scholar] [CrossRef]
- Kezlya, E.M.; Glushchenko, A.M.; Maltsev, Y.I.; Gusev, E.S.; Genkal, S.I.; Kuznetsov, A.N.; Kociolek, J.P.; Kulikovskiy, M.S. Placoneis cattiensis sp. nov. – a new, diatom (Bacillariophyceae: Cymbellales) soil species from Cát Tiên National Park (Vietnam). Phytotaxa 2020, 460, 237–248. [Google Scholar] [CrossRef]
- Cox, E.J. Coscinodiscophyceae, Mediophyceae, Fragilariophyceae, Bacillariophyceae (diatoms). In Syllabus of Plant Families: Adolf Engler’s Syllabus der Pflanzenfamilien: Photoautotrophic Eukaryotic Algae: Glaucocystophyta, Cryptophyta, Dinophyta/Dinozoa, Haptophyta, Heterokontophyta/Ochrophyta, Chlorarachniophyta/Cercozoa, Euglenophyta/Euglenozoa, Chlorophyta, Streptophyta p.p., 2nd ed.; Jaklitsch, W., Baral, H.O., Lücking, R., Lumbsch, H.T., Frey, W., Eds.; Borntraeger Verlagsbuchhandlung: Stuttgart, Germany, 2015; Volume 2/1, pp. 64–103. [Google Scholar]
- Mann, D.G. The species concept in diatoms. Phycologia 1999, 38, 437–495. [Google Scholar] [CrossRef] [Green Version]









| P. elginensis TCC499 | P. elginensis AT160Gel18 | P. elginensis UTEX FD416 | P. elginensis FD212 | P. elginensis Type [36] | |
|---|---|---|---|---|---|
| Outline | Linear | Linear | Elliptic-lanceolate to elliptic | Linear | Linear |
| Apex width (μm) | 2.8–3.2 * | 3.1 * | no | 2.8–3 * | 4–4.5 |
| Valve length (μm) | 27.6–28.4 * | 25–26 * | 8.5–10 * | 14–18 * | 30–36 |
| Valve breadth (μm) | 8–8.8 * | 6.6 * | 5.44–6.46 * | 5.6–6 * | 9–10 |
| Striae | 12–14 * | 15 * | 16–17 * | 13–15 * | 11 |
| Sampling origin | France, Ile de Mayotte Kwale River upstream site Pierre Rivière | Europe, Germany 52°57.65′ N; 08°20.67′ E AlgaTerra Culture | Minnesota, USA | Minnesota, USA | Elgin, Scotland |
| Locality | Thonon Culture Collection | AlgaTerra Culture Collection | UTEX Culture Collection | UTEX Culture Collection | Natural History Museum Collection, London. Slide BM 11751 |
| P. baikaloelginensis sp. nov. | P. abiskoensis | P. elginensis | P. paraelginensis | P. significans | P. subgastriformis sp. nov. | |
|---|---|---|---|---|---|---|
| Outline | Linear-elliptical | Linear | Linear | Linear | Linear-elliptical to elliptic-lanceolate | Lanceolate -elliptical |
| Apex shape | Subcapitate | Rostrate to subcapitate | Subcapitate | Subcapitate | Rostrate, more or less protected, obtusely rounded | Subcapitate |
| Apex width (μm) | 4.9–5 | 5–6 * | 4–4.5 | 3–4 * | 3.9–4 * | 3.8–4.3 |
| Axial area | Narrow, slightly widening to the middle of the valve | Narrow, slightly widening to the middle of the valve | Narrow, linear, barely or not broadened towards the center | Narrow, linear | Narrow, linear | Narrow, linear |
| Central area | Large, transversely-expanded or butterfly-shaped to ¾ width of valve | Transversely-expanded, butterfly-shaped or rounded, confined by 3–4 shortened striae | Larger than half of the valve width, butterfly-shaped | Rounded, slightly transversely-expanded, confined by 3–4 shortened striae | Small, with one stigma | Rounded, with one stigma |
| Valve length (μm) | 12.2–31.6 | 38–47 | 30–36 | 20–30 | 20–30 | 26–33 |
| Valve breadth (μm) | 7.8–9 | 9–11 | 8.2–10 | 6.5–8 | 10–10.5 | 9–11 |
| Striae type, number in 10 μm | Uniseriate, radiate, becoming parallel to convergent at the valve ends, 13–15 | Uniseriate, radiate, 7.5–12 | Uniseriate, radiate (angle to the raphe: 74), with one pair perpendicular to raphe very close to the ends, 9–12.4 | Uniseriate, radiate, becoming subparallel to convergent at the valve ends, 12–18 | Uniseriate, radiate, 10–11 | Uniseriate, radiate, slightly curved 9–11 |
| Areolae, number in 10 μm | Not discernible in the LM, 30 | Distinct in the LM, 26–30 | Difficult to resolve in the LM, 22.4–28.9 | Difficult to resolve in the LM, 36* | Indiscernible in LM | Not discernible in the LM |
| Distribution | Baikal | Holarctic | Holarctic | Lake “Schmaler Luzin” in North Germany; Ohrid and Prespa lakes, Shkodra (Albanian part) | Widespread species | |
| References | This study | [33,36,42,43] | [36,44,45] | [20,33,36,44,46,47] | [5,46] | [33,47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kezlya, E.; Glushchenko, A.; Maltsev, Y.; Gusev, E.; Genkal, S.; Kociolek, J.P.; Kulikovskiy, M. Three New Species of Placoneis Mereschkowsky (Bacillariophyceae: Cymbellales) with Comments on Cryptic Diversity in the P. elginensis—Group. Water 2021, 13, 3276. https://doi.org/10.3390/w13223276
Kezlya E, Glushchenko A, Maltsev Y, Gusev E, Genkal S, Kociolek JP, Kulikovskiy M. Three New Species of Placoneis Mereschkowsky (Bacillariophyceae: Cymbellales) with Comments on Cryptic Diversity in the P. elginensis—Group. Water. 2021; 13(22):3276. https://doi.org/10.3390/w13223276
Chicago/Turabian StyleKezlya, Elena, Anton Glushchenko, Yevhen Maltsev, Evgeniy Gusev, Sergey Genkal, John Patrick Kociolek, and Maxim Kulikovskiy. 2021. "Three New Species of Placoneis Mereschkowsky (Bacillariophyceae: Cymbellales) with Comments on Cryptic Diversity in the P. elginensis—Group" Water 13, no. 22: 3276. https://doi.org/10.3390/w13223276
APA StyleKezlya, E., Glushchenko, A., Maltsev, Y., Gusev, E., Genkal, S., Kociolek, J. P., & Kulikovskiy, M. (2021). Three New Species of Placoneis Mereschkowsky (Bacillariophyceae: Cymbellales) with Comments on Cryptic Diversity in the P. elginensis—Group. Water, 13(22), 3276. https://doi.org/10.3390/w13223276