
Photosynthetic Characteristics and Yield Response of Isatis indigotica to Regulated Deficit Irrigation in a Cold and Arid Environment

Abstract
:1. Introduction
2. Materials and Methods
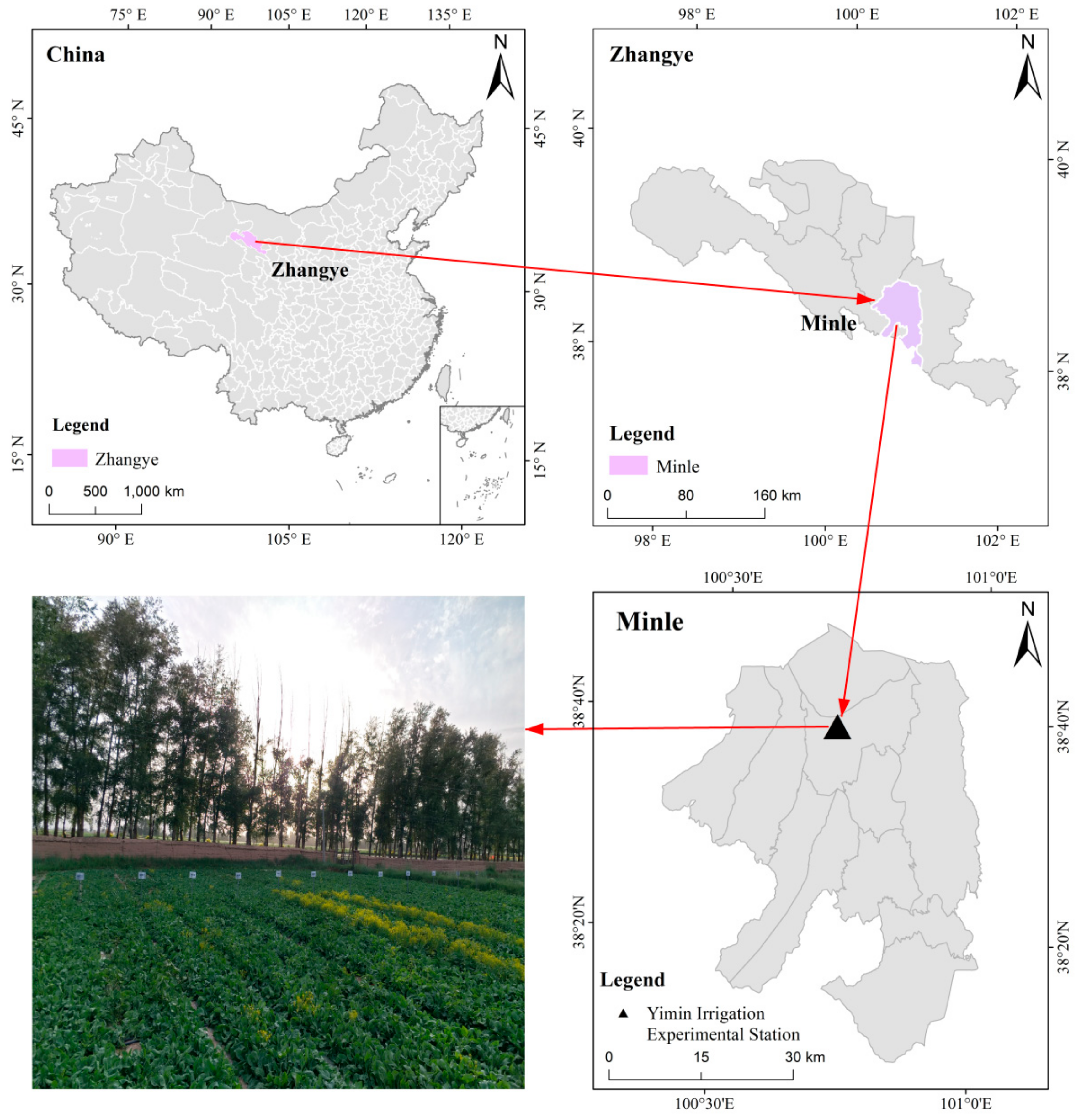
2.1. Experimental Site
2.2. Experimental Design and Method
2.3. Plant Sampling and Measurements
2.3.1. Photosynthetic Physiological Indices
2.3.2. Dry Matter
2.3.3. Yield and Harvest Index
2.3.4. Water Use Efficiency and Irrigation Water Use Efficiency
2.4. Statistical Analysis
3. Results
3.1. Photosynthetic Characteristics
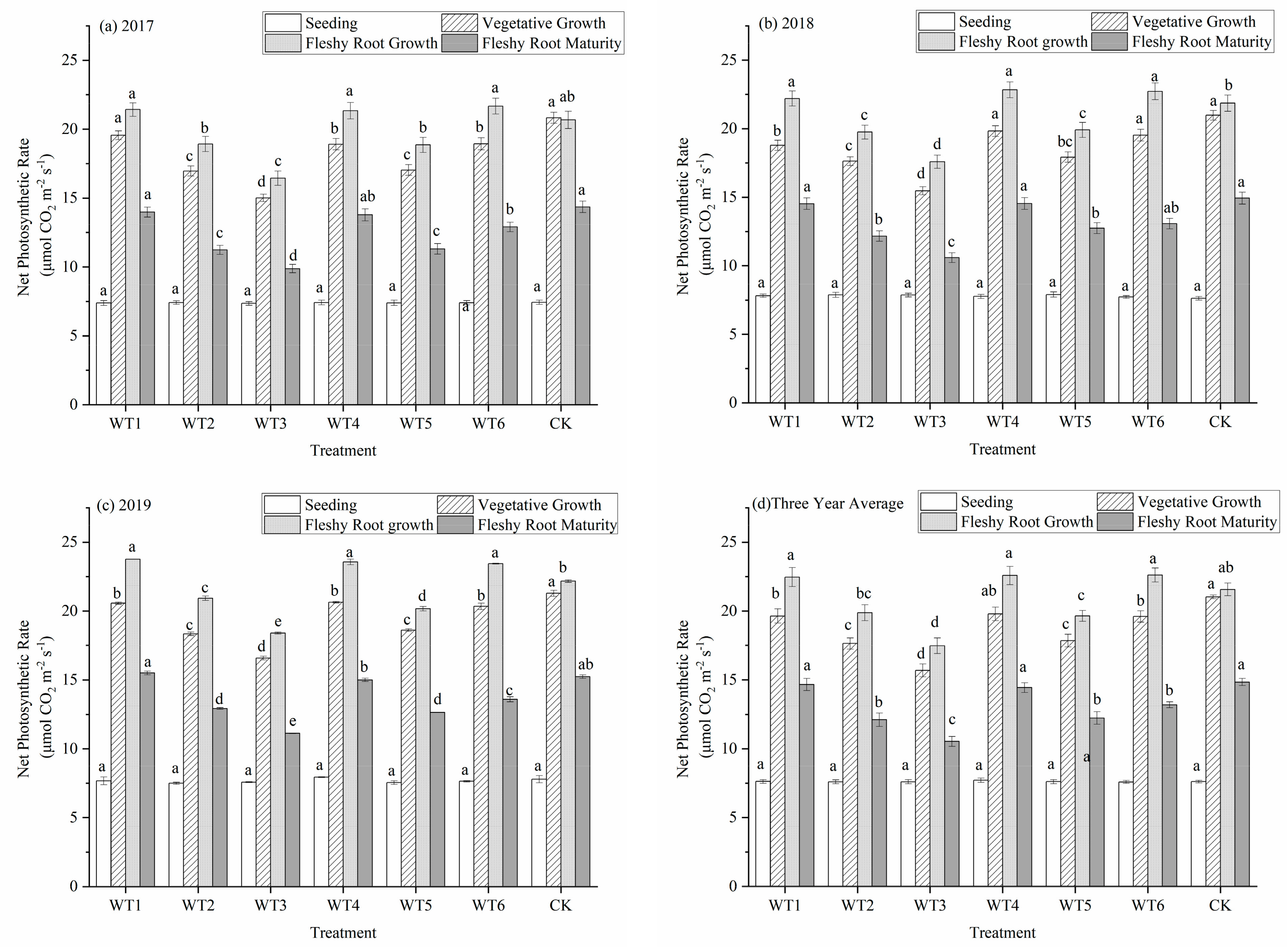
3.1.1. Net Photosynthetic Rate
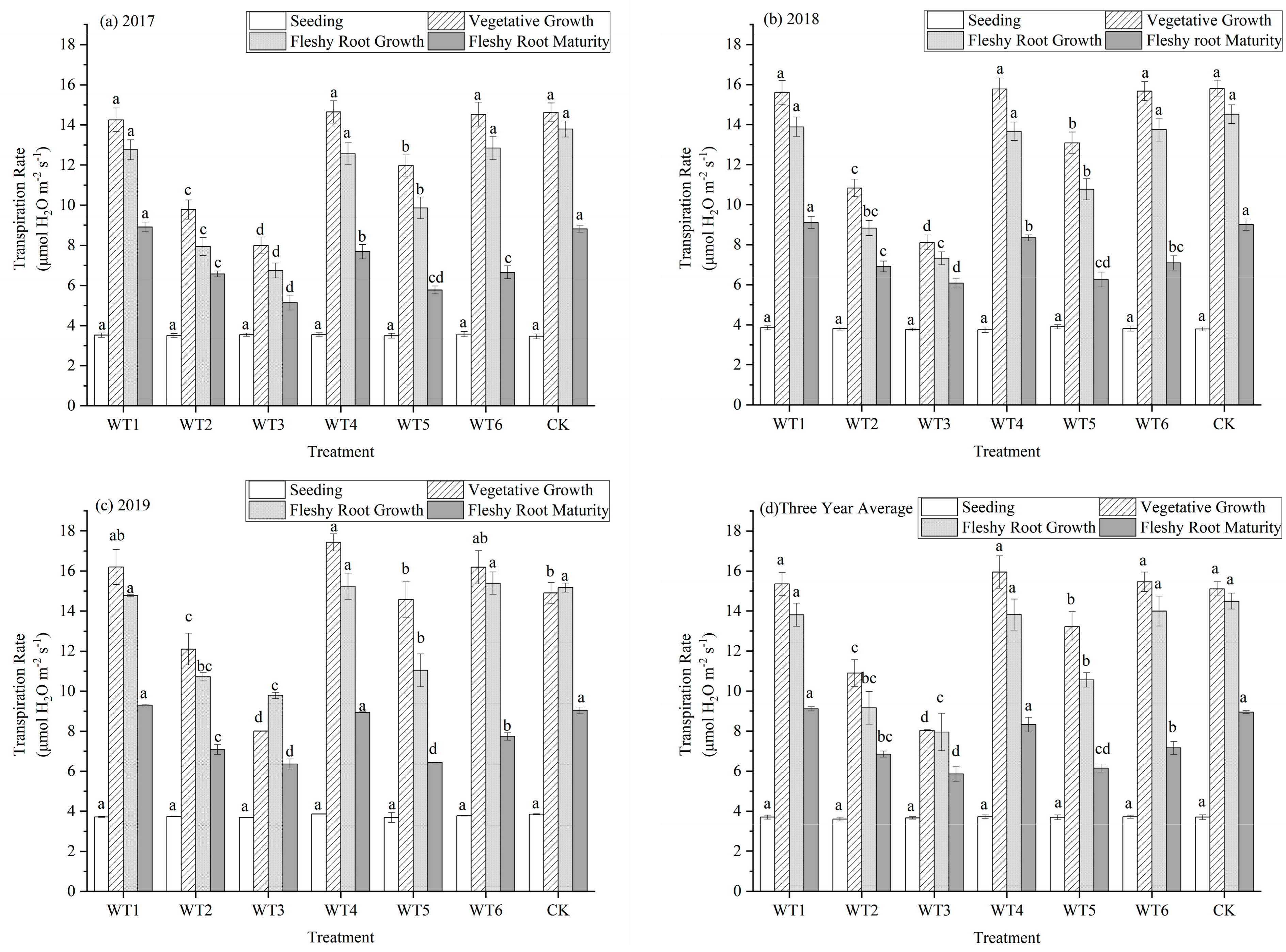
3.1.2. Transpiration Rate
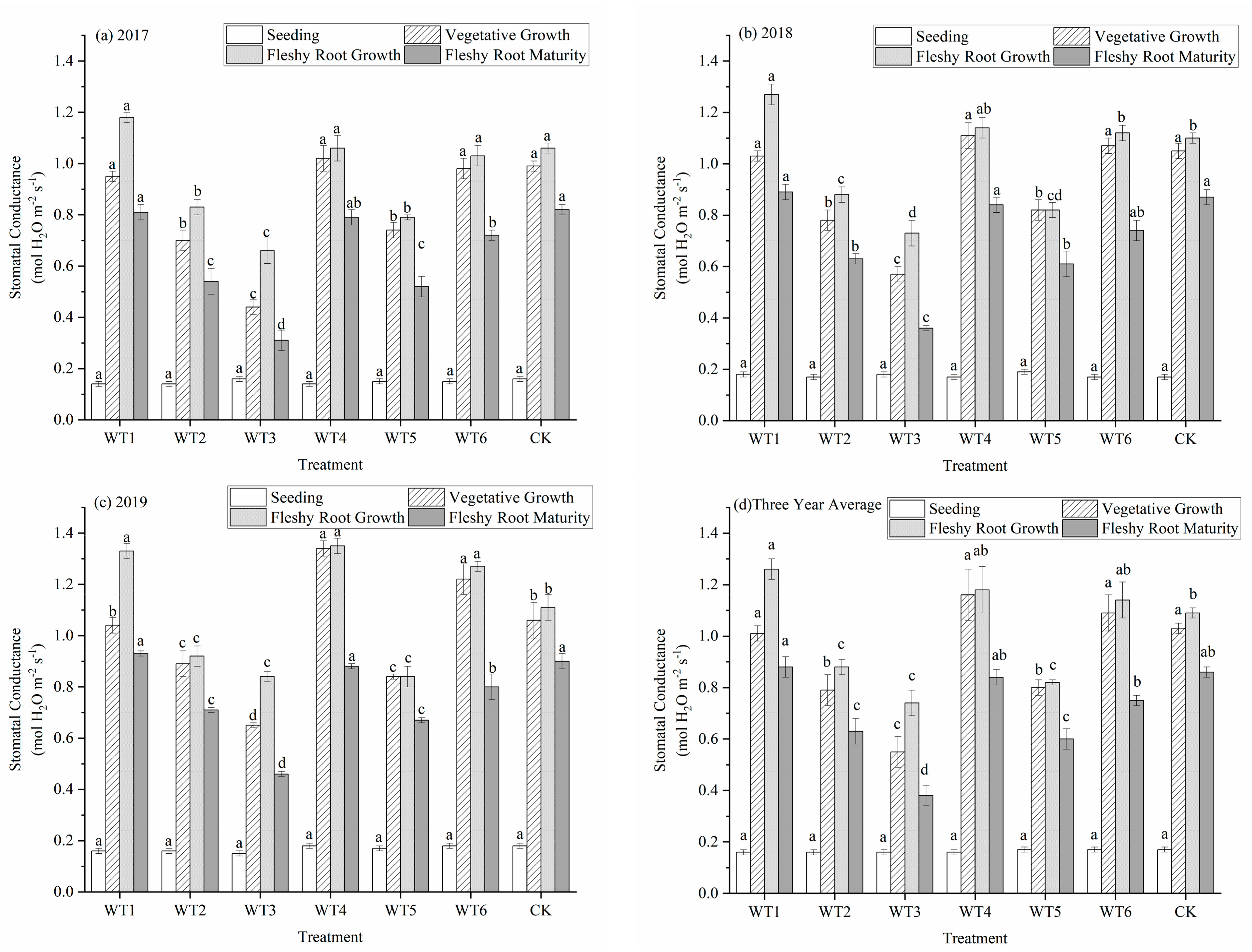
3.1.3. Stomatal Conductance
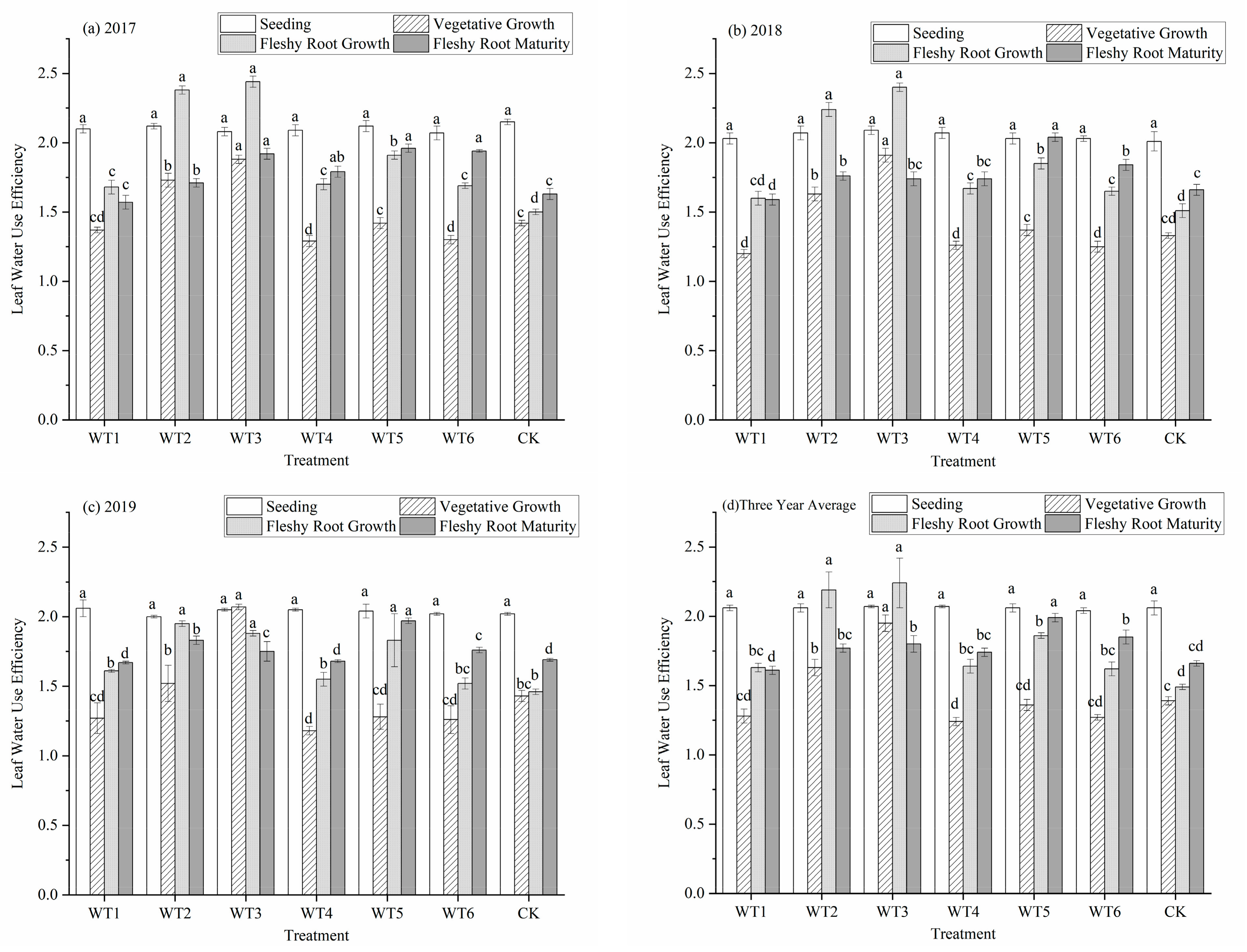
3.1.4. Leaf Water Use Efficiency
3.2. Economic Yield and Yield Components
3.2.1. Economic Yield
3.2.2. Yield Components
3.3. Total Biomass and Harvest Index
3.4. Water Consumption, Water Use Efficiency, and Irrigation Water Use Efficiency
3.4.1. Water Consumption
3.4.2. Water Use Efficiency and Irrigation Water Use Efficiency
3.5. Correlations among Various Indices
4. Discussion
4.1. Photosynthetic Characteristics
4.2. Yield and Yield Components
4.3. Total Biomass and Harvest Index
4.4. Water Consumption
4.5. Water Use Efficiency and Irrigation Water Use Efficiency
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Han, J.; Jiang, X.; Zhang, L. Optimisation of extraction conditions for polysaccharides from the roots of Isatis tinctoria L. by response surface methodology and their in vitro free radicals scavenging activities and effects on IL-4 and IFN-γ mRNA expression in chicken lymphocytes. Carbohydr. Polym. 2011, 86, 1320–1326. [Google Scholar] [CrossRef]
- Hamburger, M. Isatis tinctorial—From the rediscovery of an ancient medicinal plant towards a novel anti-inflammatory phytopharmaceutical. Phytochem. Rev. 2002, 1, 333. [Google Scholar] [CrossRef]
- Recio, M.C.; Cerdá-Nicolás, M.; Potterat, O.; Hamburger, M.; Ríos, J. Anti-Inflammatory and Antiallergic Activity in vivo of Lipophilic Isatis tinctoria Extracts and Tryptanthrin. Planta Med. 2006, 72, 539–546. [Google Scholar] [CrossRef] [Green Version]
- Condurso, C.; Verzera, A.; Romeo, V.; Ziino, M.; Ragusa, S. The Leaf Volatile Constituents of Isatis tinctoria by Solid-Phase Microextraction and Gas Chromatography/Mass Spectrometry. Planta Med. 2006, 72, 924–928. [Google Scholar] [CrossRef]
- Ke, L.; Wen, T.; Bradshaw, J.P.; Zhou, J.; Rao, P. Antiviral Decoction of Isatidis Radix (bn lán gēn) Inhibited Influenza Virus Adsorption on MDCK Cells by Cytoprotective Activity. J. Tradit. Complement. Med. 2012, 2, 47–51. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.; Liu, H.; Zhang, Z.; Li, P. Antioxidant and anti-inflammatory activities of Radix Isatidis polysaccharide in murine alveolar macrophages. Int. J. Biol. Macromol. 2013, 58, 329–335. [Google Scholar] [CrossRef]
- Nagendrappa, G. Chemistry triggered the first civil disobedience movement in India. J. Reson. 2003, 8, 42–48. [Google Scholar] [CrossRef]
- Spataro, G.; Negri, V. Adaptability and variation in Isatis tinctoria L.: A new crop for Europe. Euphytica 2008, 163, 89–102. [Google Scholar] [CrossRef]
- Author Chai, Q.; Gan, Y.T.; Zhao, C.; Xu, H.L.; Waskom, R.M.; Niu, Y.N.; Siddique, K.H.M. Regulated deficit irrigation for crop production under drought stress. A review. Agron. Sustain. Dev. 2016, 36, 3. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Du, T.; Qiu, R.; Chen, J.; Wang, F.; Li, Y.; Wang, C.; Gao, L.; Kang, S. Improved water use efficiency and fruit quality of greenhouse crops under regulated deficit irrigation in northwest China. Agric. Water Manag. 2017, 179, 193–204. [Google Scholar] [CrossRef]
- Kang, S.; Zhang, L.; Liang, Y.; Hu, X.; Cai, H.; Gu, B. Effects of limited irrigation on yield and water use efficiency of winter wheat in the Loess Plateau of China. Agric. Water Manag. 2002, 55, 203–216. [Google Scholar] [CrossRef]
- Du, T.; Kang, S.; Sun, J.; Zhang, X.; Zhang, J. An improved water use efficiency of cereals under temporal and spatial deficit irrigation in north China. Agric. Water Manag. 2010, 97, 66–74. [Google Scholar] [CrossRef]
- Tolk, J.A.; Howell, T.A. Water use efficiencies of grain sorghum grown in three USA southern Great Plains soils. Agric. Water Manag. 2003, 59, 97–111. [Google Scholar] [CrossRef]
- Pascual-Seva, N.; San Bautista, A.; Lopez-Galarza, S.; Maroto, J.V.; Pascual, B. Response of drip-irrigated chufa (Cyperus esculentus L. var. sativus Boeck.) to different planting configurations: Yield and irrigation water-use efficiency. Agric. Water Manag. 2016, 170, 140–147. [Google Scholar] [CrossRef]
- Costa, J.M.; Ortuño, M.; Chaves, M.M. Deficit Irrigation as a Strategy to Save Water: Physiology and Potential Application to Horticulture. J. Integr. Plant Biol. 2010, 49, 1421–1434. [Google Scholar] [CrossRef]
- Capra, A.; Consoli, S.; Scicolone, B. Water management strategies under deficit irrigation. J. Agric. Eng. 2008, 39, 27. [Google Scholar] [CrossRef]
- Ruiz-Sanchez, M.C.; Domingo, R.; Castel, J.R. Review. Deficit irrigation in fruit trees and vines in Spain. Span J. Agric. Res. 2010, 8, S5–S20. [Google Scholar] [CrossRef] [Green Version]
- Fereres, E.; Soriano, M.A. Deficit irrigation for reducing agricultural water use. J. Exp. Bot. 2007, 58, 147–159. [Google Scholar] [CrossRef] [Green Version]
- Kuscu, H.; Turhan, A.; Demir, A.O. The response of processing tomato to deficit irrigation at various phenological stages in a sub-humid environment. Agric. Water Manag. 2014, 133, 92–103. [Google Scholar] [CrossRef]
- Liu, E.K.; Mei, X.R.; Yan, C.R.; Gong, D.Z.; Zhang, Y.Q. Effects of water stress on photosynthetic characteristics, dry matter translocation and WUE in two winter wheat genotypes. Agric. Water Manag. 2016, 167, 75–85. [Google Scholar] [CrossRef]
- Igbadun, H.E.; Ramalan, A.A.; Oiganji, E. Effects of regulated deficit irrigation and mulch on yield, water use and crop water productivity of onion in Samaru, Nigeria. Agric. Water Manag. 2012, 109, 162–169. [Google Scholar] [CrossRef]
- Conesa, M.R.; Garcia-Salinas, M.D.; de la Rosa, J.M.; Fernandez-Trujillo, J.P.; Domingo, R.; Perez-Pastor, A. Effects of deficit irrigation applied during fruit growth period of late mandarin trees on harvest quality, cold storage and subsequent shelf-life. Sci. Hortic. 2014, 165, 344–351. [Google Scholar] [CrossRef]
- Marsal, J.; Casadesus, J.; Lopez, G.; Mata, M.; Bellvert, J.; Girona, J. Sustainability of regulated deficit irrigation in a mid-maturing peach cultivar. Irrig. Sci. 2016, 34, 201–208. [Google Scholar] [CrossRef]
- Erdem, Y.; Yuksel, A.N. Yield response of watermelon to irrigation shortage. Sci. Hortic. 2003, 98, 365–383. [Google Scholar] [CrossRef]
- Ghosh, A.; Agrawal, M.; Agrawal, S.B. Effect of water deficit stress on an Indian wheat cultivar (Triticum aestivum L. HD 2967) under ambient and elevated level of ozone. Sci. Total Environ. 2020, 714, 16. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Cai, F.; Zhao, Z.Q.; Meng, Y.; Gao, L.Y.; Zhao, T.H. Physiological and Dry Matter Characteristics of Spring Maize in Northeast China under Drought Stress. Water 2018, 10, 1561. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Wang, Y.; Wang, Q.; Guo, S.; Liu, Y.; Liu, J.; Tang, Z.; Wang, Z. Changes in Growth and Photosynthetic Parameters and Medicinal Compounds in Eleutherococcus senticosus Harms under Drought Stress. Hortscience 2019, 54, 2202–2208. [Google Scholar] [CrossRef] [Green Version]
- Chastain, D.R.; Snider, J.L.; Collins, G.D.; Perry, C.D.; Whitaker, J.; Byrd, S.A. Water deficit in field-grown Gossypium hirsutum primarily limits net photosynthesis by decreasing stomatal conductance, increasing photorespiration, and increasing the ratio of dark respiration to gross photosynthesis. J. Plant Physiol. 2014, 171, 1576–1585. [Google Scholar] [CrossRef]
- Tesfaye, S.G.; Ismail, M.R.; Ramlan, M.F.; Marziah, M.; Kausar, H. Effect of soil drying on rate of stress development, leaf gas exchange and proline accumulation in robusta coffee (Coffea canephora Pierre ex Froehner) clones. Exp. Agric. 2014, 50, 458–479. [Google Scholar] [CrossRef] [Green Version]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.C.; Duan, A.W.; Wang, R.; Guan, Z.M.; Yang, S.J.; Ma, S.T.; Shao, Y. Root-sourced signal and photosynthetic traits, dry matter accumulation and remobilization, and yield stability in winter wheat as affected by regulated deficit irrigation. Agric. Water Manag. 2015, 148, 123–129. [Google Scholar] [CrossRef]
- Miyashita, K.; Tanakamaru, S.; Maitani, T.; Kimura, K. Recovery responses of photosynthesis, transpiration, and stomatal conductance in kidney bean following drought stress. Environ. Exp. Bot. 2005, 53, 205–214. [Google Scholar] [CrossRef]
- Ali, S.; Xu, Y.; Jia, Q.; Ahmad, I.; Wei, T.; Ren, X.; Zhang, P.; Din, R.; Cai, T.; Jia, Z. Cultivation techniques combined with deficit irrigation improves winter wheat photosynthetic characteristics, dry matter translocation and water use efficiency under simulated rainfall conditions. Agric. Water Manag. 2018, 201, 207–218. [Google Scholar] [CrossRef]
- Ahmed, C.B.; Rouina, B.B.; Boukhris, M. Effects of water deficit on olive trees cv. Chemlali under field conditions in arid region in Tunisia. Sci. Hortic. 2007, 113, 267–277. [Google Scholar] [CrossRef]
- Cui, N.; Du, T.; Li, F.; Ling, T.; Kang, S.; Wang, M.; Liu, X.; Li, Z. Response of vegetative growth and fruit development to regulated deficit irrigation at different growth stages of pear-jujube tree. Agric. Water Manag. 2009, 96, 1237–1246. [Google Scholar] [CrossRef]
- Yan, S.C.; Wu, Y.; Fan, J.L.; Zhang, F.C.; Qiang, S.C.; Zheng, J.; Xiang, Y.Z.; Guo, J.J.; Zou, H.Y. Effects of water and fertilizer management on grain filling characteristics, grain weight and productivity of drip-fertigated winter wheat. Agric. Water Manag. 2019, 213, 983–995. [Google Scholar] [CrossRef]
- Anda, A.; Soos, G.; Menyhart, L.; Kucserka, T.; Simon, B. Yield features of two soybean varieties under different water supplies and field conditions. Field Crop. Res. 2020, 245, 107673. [Google Scholar] [CrossRef]
- Fabeiro, C.; de Santa Olalla, F.M.; Lopez, R.; Dominguez, A. Production and quality of the sugar beet (Beta vulgaris L.) cultivated under controlled deficit irrigation conditions in a semi-arid climate. Agric. Water Manag. 2003, 62, 215–227. [Google Scholar] [CrossRef]
- Zhang, H.P.; Turner, N.C.; Poole, M.L. Increasing the harvest index of wheat in the high rainfall zones of southern Australia. Field Crop. Res. 2012, 129, 111–123. [Google Scholar] [CrossRef]
- Abid, M.; Shao, Y.H.; Liu, S.X.; Wang, F.; Gao, J.W.; Jiang, D.; Tian, Z.W.; Dai, T.B. Pre-drought priming sustains grain development under post-anthesis drought stress by regulating the growth hormones in winter wheat (Triticum aestivum L.). Planta 2017, 246, 509–524. [Google Scholar] [CrossRef]
- Iqbal, M.; Ashraf, M. Gibberellic acid mediated induction of salt tolerance in wheat plants: Growth, ionic partitioning, photosynthesis, yield and hormonal homeostasis. Environ. Exp. Bot. 2013, 86, 76–85. [Google Scholar] [CrossRef]
- Mitchell, J.H.; Rebetzke, G.J.; Chapman, S.C.; Fukai, S. Evaluation of reduced-tillering (tin) wheat lines in managed, terminal water deficit environments. J. Exp. Bot. 2013, 64, 3439–3451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashid, M.A.; Andersen, M.N.; Wollenweber, B.; Zhang, X.; Olesen, J.E. Acclimation to higher VPD and temperature minimized negative effects on assimilation and grain yield of wheat. Agric. For. Meteorol. 2018, 248, 119–129. [Google Scholar] [CrossRef]
- Faloye, O.T.; Alatise, M.O.; Ajayi, A.E.; Ewulo, B.S. Effects of biochar and inorganic fertiliser applications on growth, yield and water use efficiency of maize under deficit irrigation. Agric. Water Manag. 2019, 217, 165–178. [Google Scholar] [CrossRef]
- Marsal, J.; Lopez, G.; del Campo, J.; Mata, M.; Arbones, A.; Girona, J. Postharvest regulated deficit irrigation in ‘Summit’ sweet cherry: Fruit yield and quality in the following season. Irrig. Sci. 2010, 28, 181–189. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Huang, Z.; Zhu, Q.; Wang, L. Remobilization of Carbon Reserves Is Improved by Controlled Soil-Drying during Grain Filling of Wheat. Crop Sci. 2000, 40, 1645–1655. [Google Scholar] [CrossRef]
- Chen, J.L.; Kang, S.Z.; Du, T.S.; Qiu, R.J.; Guo, P.; Chen, R.Q. Quantitative response of greenhouse tomato yield and quality to water deficit at different growth stages. Agric. Water Manag. 2013, 129, 152–162. [Google Scholar] [CrossRef]
- Wang, F.; Xie, R.; Ming, B.; Wang, K.; Hou, P.; Chen, J.; Liu, G.; Zhang, G.; Xue, J.; Li, S. Dry matter accumulation after silking and kernel weight are the key factors for increasing maize yield and water use efficiency. Agric. Water Manag. 2021, 254, 106938. [Google Scholar] [CrossRef]
- Sun, H.Y.; Liu, C.M.; Zhang, X.Y.; Shen, Y.J.; Zhang, Y.Q. Effects of irrigation on water balance, yield and WUE of winter wheat in the North China Plain. Agric. Water Manag. 2006, 85, 211–218. [Google Scholar] [CrossRef]
- Belay, S.A.; Schmitter, P.; Worqlul, A.W.; Steenhuis, T.S.; Reyes, M.R.; Tilahun, S.A. Conservation Agriculture Saves Irrigation Water in the Dry Monsoon Phase in the Ethiopian Highlands. Water 2019, 11, 2103. [Google Scholar] [CrossRef] [Green Version]
- Patane, C.; Tringali, S.; Sortino, O. Effects of deficit irrigation on biomass, yield, water productivity and fruit quality of processing tomato under semi-arid Mediterranean climate conditions. Sci. Hortic. 2011, 129, 590–596. [Google Scholar] [CrossRef]
- Bloch, D.; Hoffmann, C.M. Impact of water supply on photosynthesis, water use and carbon isotope discrimination of sugar beet genotypes. Eur. J. Agron. 2006, 24, 218–225. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Water Deficit Timing | Water Deficit Severity | Seeding | Vegetative Growth | Fleshy Root Growth | Flesh Root Maturity |
|---|---|---|---|---|---|---|
| WT1 | Vegetative growth (VG) | Mild | 75–85% a | 65–75% | 75–85% | 75–85% |
| WT2 | VG | Moderate | 75–85% | 55–65% | 75–85% | 75–85% |
| WT3 | VG | Severe | 75–85% | 45–55% | 75–85% | 75–85% |
| WT4 | VG + Fleshy root growth | Mild | 75–85% | 65–75% | 65–75% | 75–85% |
| WT5 | VG + Fleshy root growth | Moderate | 75–85% | 55–65% | 55–65% | 75–85% |
| WT6 | VG + Fleshy root maturity | Mild | 75–85% | 65–75% | 75–85% | 65–75% |
| CK | N/A | None | 75–85% | 75–85% | 75–85% | 75–85% |
| Year | Treatments | Lateral Root Number | Taproot Length (cm) | Taproot Diameter (cm) | Root Biomass (g) | Total Biomass (kg·ha−2) | Yield (kg·ha−2) | Harvest Index |
|---|---|---|---|---|---|---|---|---|
| 2017 | WT1 | 10.70a | 23.24a | 1.64a | 13.55a | 12,476.23a | 8390.80a | 0.6725ab |
| WT2 | 9.30ab | 22.20ab | 1.55bc | 12.02bc | 11,015.42d | 7462.24b | 0.6774a | |
| WT3 | 7.30b | 18.67c | 1.40d | 10.59d | 10,186.77e | 6800.36c | 0.6676b | |
| WT4 | 10.30ab | 23.25a | 1.66a | 11.08cd | 12,084.28b | 8235.32a | 0.6815a | |
| WT5 | 9.30ab | 19.93b | 1.44c | 11.01cd | 10,111.34e | 6819.79c | 0.6745a | |
| WT6 | 9.33ab | 22.71a | 1.59b | 11.88c | 11,542.37c | 7713.45b | 0.6683b | |
| CK | 10.70a | 23.21a | 1.63a | 12.66ab | 12,489.96a | 8322.25a | 0.6663b | |
| 2018 | WT1 | 10.67a | 24.33a | 1.66a | 13.47a | 12,577.33a | 8475.38a | 0.6739ab |
| WT2 | 9.36b | 23.26b | 1.47bc | 11.94b | 11,116.52bc | 7638.14b | 0.6871a | |
| WT3 | 8.03c | 19.77d | 1.42c | 10.49d | 10,487.87d | 6986.12c | 0.6661b | |
| WT4 | 10.63a | 24.38a | 1.71a | 11.07c | 12,185.38ab | 8308.44a | 0.6818a | |
| WT5 | 9.67b | 20.78c | 1.46bc | 10.94cd | 10,212.44d | 6923.72c | 0.6780ab | |
| WT6 | 8.67bc | 23.86ab | 1.53b | 11.27bc | 11,524.31b | 7846.42b | 0.6809a | |
| CK | 11.00a | 23.20b | 1.65a | 12.58ab | 12,591.06a | 8348.91a | 0.6631b | |
| 2019 | WT1 | 11.33a | 31.28ab | 1.92bc | 14.77a | 12,637.75a | 8554.18a | 0.6769bc |
| WT2 | 9.00b | 30.07c | 1.72d | 13.26b | 11,174.88b | 7623.76c | 0.6822b | |
| WT3 | 7.67c | 26.86d | 1.69d | 11.31d | 10,350.15c | 6959.82d | 0.6724c | |
| WT4 | 11.00a | 31.41ab | 1.97a | 15.08a | 12,247.66a | 8398.70a | 0.6857b | |
| WT5 | 8.67bc | 28.08d | 1.71d | 11.69cd | 10,272.86c | 6979.25d | 0.6794bc | |
| WT6 | 9.33b | 30.45bc | 1.88c | 13.07bc | 11,424.15b | 7934.63b | 0.6945a | |
| CK | 11.67a | 31.37ab | 1.91bc | 13.29b | 12,649.42a | 8521.77a | 0.6737c | |
| Average | WT1 | 10.90a | 26.28a | 1.74a | 13.93a | 12,563.77a | 8473.45a | 0.6744ab |
| WT2 | 9.22b | 25.18b | 1.58c | 12.41ab | 11,102.27c | 7574.71c | 0.6823a | |
| WT3 | 7.67c | 21.77d | 1.50d | 10.80d | 10,341.60d | 6915.43d | 0.6687b | |
| WT4 | 10.64a | 26.35a | 1.78a | 12.41ab | 12,172.44a | 8314.15a | 0.6830a | |
| WT5 | 9.21bc | 22.93c | 1.54cd | 11.21c | 10,198.88d | 6907.59d | 0.6773ab | |
| WT6 | 9.11b | 25.67ab | 1.67b | 12.07b | 11,496.94b | 7831.50b | 0.6812a | |
| CK | 11.12a | 25.93a | 1.73a | 12.84ab | 12,576.81a | 8397.64a | 0.6677b |
| Year | Treatments | Water Consumption (mm) | Irrigation Amount (mm) | WUE (kg·ha−2·mm−1) | IWUE (kg·ha−2·mm−1) |
|---|---|---|---|---|---|
| 2017 | WT1 | 355.25c | 153.76b | 23.62a | 54.57a |
| WT2 | 366.06b | 149.57b | 20.39c | 49.89c | |
| WT3 | 343.62d | 132.12c | 19.79b | 51.47d | |
| WT4 | 353.93c | 152.42b | 23.27a | 54.03a | |
| WT5 | 347.35d | 135.85c | 19.63bc | 50.20d | |
| WT6 | 367.23b | 152.61b | 21.00b | 50.54bc | |
| CK | 381.75a | 165.26a | 21.80bc | 50.36b | |
| 2018 | WT1 | 363.32bc | 153.02b | 23.33a | 53.39a |
| WT2 | 371.17b | 150.87b | 20.58c | 50.63c | |
| WT3 | 348.46bcd | 133.16c | 20.05d | 52.46b | |
| WT4 | 362.12bc | 151.81b | 22.94a | 54.73a | |
| WT5 | 355.95c | 137.64c | 19.45d | 50.30c | |
| WT6 | 362.94bc | 149.35b | 21.62b | 52.34b | |
| CK | 386.55a | 166.26a | 21.60b | 50.22c | |
| 2019 | WT1 | 367.09bc | 170.49ab | 23.20a | 50.17a |
| WT2 | 373.30b | 163.00bc | 20.40bc | 46.77b | |
| WT3 | 351.41d | 145.41c | 19.81bc | 47.86b | |
| WT4 | 359.89cd | 163.69ab | 23.34a | 51.31a | |
| WT5 | 361.75cd | 156.15bc | 19.29c | 44.70c | |
| WT6 | 367.92bc | 171.52ab | 21.57b | 46.26bc | |
| CK | 393.85a | 178.35a | 21.63b | 47.78ab | |
| Average | WT1 | 361.89bcd | 159.09ab | 23.38a | 53.38a |
| WT2 | 370.18b | 154.48ab | 20.46c | 49.10ab | |
| WT3 | 347.83e | 136.90c | 19.88d | 50.60ab | |
| WT4 | 358.65cd | 155.97ab | 23.18a | 53.36a | |
| WT5 | 355.02de | 143.21bc | 19.46d | 48.40b | |
| WT6 | 366.03bc | 157.83ab | 21.40b | 49.71ab | |
| CK | 387.38a | 169.96a | 21.68b | 49.45ab |
| Correlation Coefficient | Pn | Tr | Gs | Taproot Length | Taproot Diameter | Lateral Root Number | Root Biomass | Irrigation Amount | Water Consumption | Total Biomass | Yield |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Pn | 1 | ||||||||||
| Tr | 0.953 ** | 1 | . | ||||||||
| Gs | 0.986 ** | 0.925 ** | 1 | ||||||||
| Taproot length | 0.888 ** | 0.862 * | 0.938 ** | 1 | |||||||
| Taproot diameter | 0.955 ** | 0.929 ** | 0.946 ** | 0.915 ** | 1 | ||||||
| Lateral root number | 0.970 ** | 0.942 ** | 0.945 ** | 0.835 * | 0.890 ** | 1 | |||||
| Root biomass | 0.831 * | 0.907 ** | 0.859 * | 0.863 * | 0.792 * | 0.839 * | 1 | ||||
| Irrigation amount | 0.871 * | 0.861 * | 0.885 ** | 0.888 ** | 0.814 * | 0.840 * | 0.795 * | 1 | |||
| Water consumption | 0.600 | 0.599 | 0.606 | 0.621 | 0.483 | 0.621 | 0.543 | 0.891 ** | 1 | ||
| Total biomass | 0.929 ** | 0.984 ** | 0.912 ** | 0.897 ** | 0.944 ** | 0.892 ** | 0.890 ** | 0.888 ** | 0.625 | 1 | |
| Yield | 0.934 ** | 0.973 ** | 0.931 ** | 0.935 ** | 0.962 ** | 0.891 ** | 0.895 ** | 0.886 ** | 0.603 | 0.994 ** | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, C.; Zhang, H.; Li, F.; Wang, Z.; Wang, Y. Photosynthetic Characteristics and Yield Response of Isatis indigotica to Regulated Deficit Irrigation in a Cold and Arid Environment. Water 2021, 13, 3510. https://doi.org/10.3390/w13243510
Zhou C, Zhang H, Li F, Wang Z, Wang Y. Photosynthetic Characteristics and Yield Response of Isatis indigotica to Regulated Deficit Irrigation in a Cold and Arid Environment. Water. 2021; 13(24):3510. https://doi.org/10.3390/w13243510
Chicago/Turabian StyleZhou, Chenli, Hengjia Zhang, Fuqiang Li, Zeyi Wang, and Yucai Wang. 2021. "Photosynthetic Characteristics and Yield Response of Isatis indigotica to Regulated Deficit Irrigation in a Cold and Arid Environment" Water 13, no. 24: 3510. https://doi.org/10.3390/w13243510
APA StyleZhou, C., Zhang, H., Li, F., Wang, Z., & Wang, Y. (2021). Photosynthetic Characteristics and Yield Response of Isatis indigotica to Regulated Deficit Irrigation in a Cold and Arid Environment. Water, 13(24), 3510. https://doi.org/10.3390/w13243510