
Multislot Fishway Improves Entrance Performance and Fish Transit Time over Vertical Slots

 , ,
, ,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
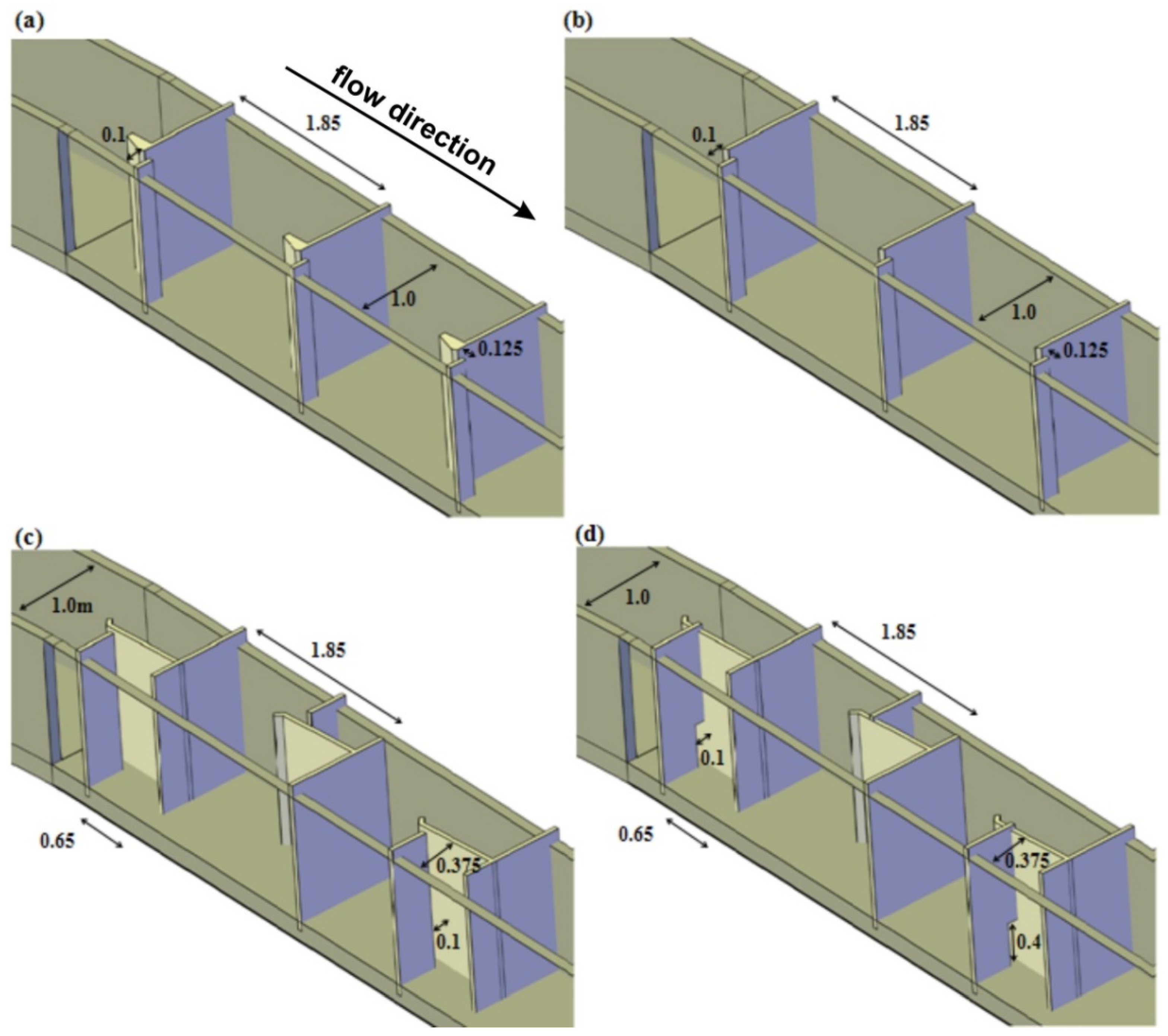
2.1. Experimental Facility
2.2. Fish Capture
2.3. Fish Experiments
2.4. Data Analysis
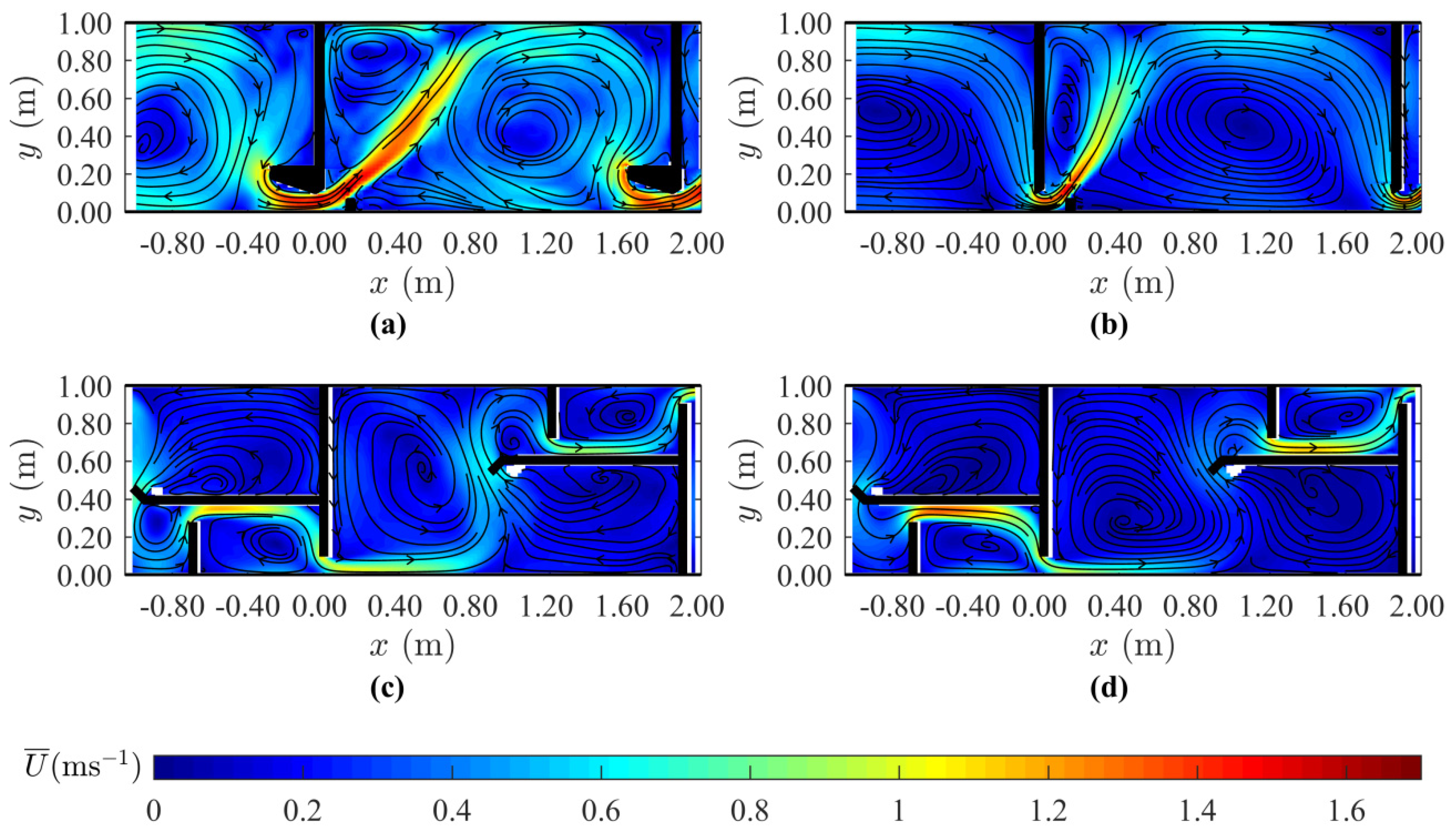
2.5. Hydrodynamic Characterization
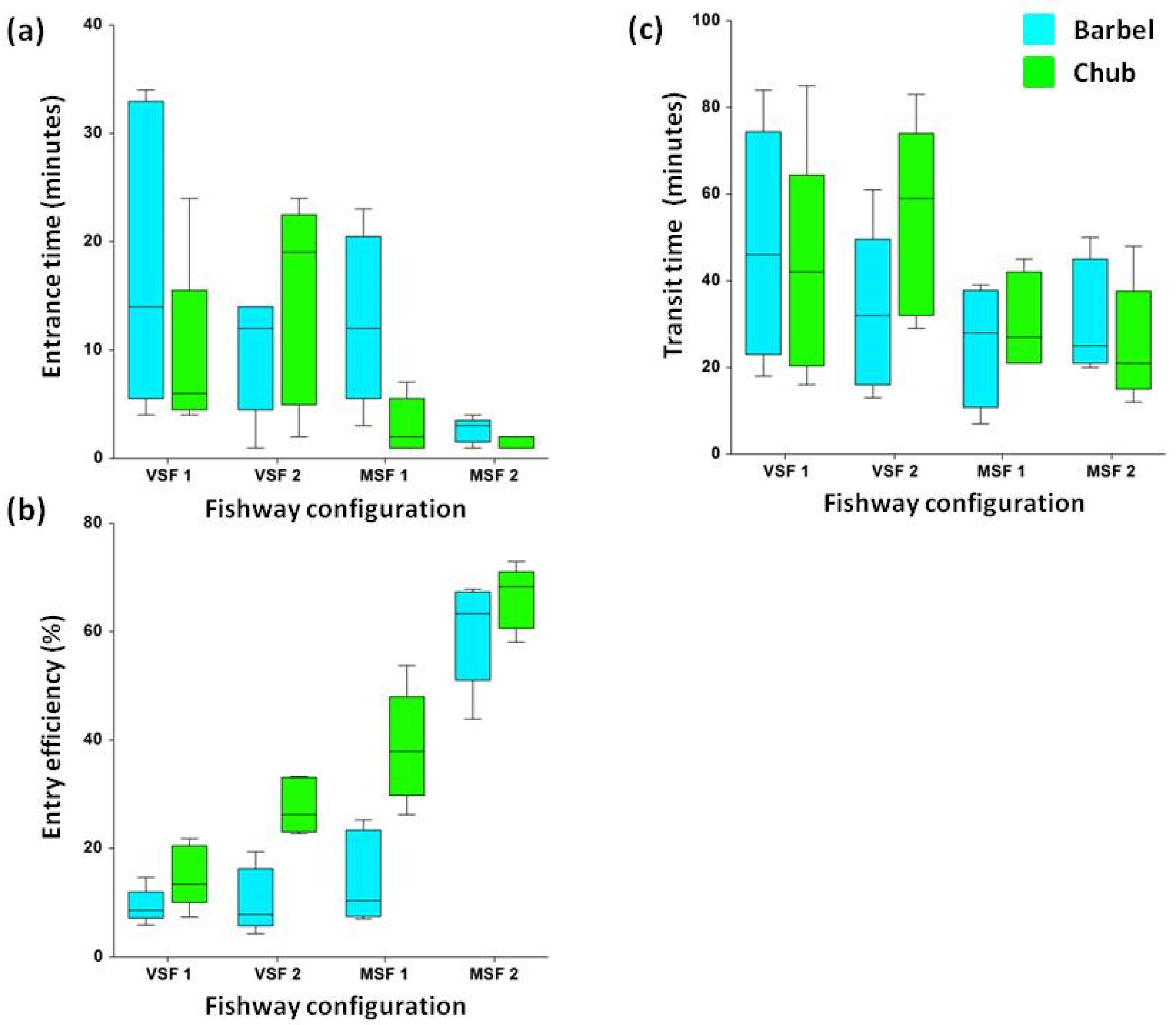
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.-I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.-H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- AMBER: Adaptive Management of Barriers in European Rivers. Available online: http://www.amber.international (accessed on 5 August 2020).
- Katopodis, C. Developing a toolkit for fish passage, ecological flow management and fish habitat works. J. Hydraul. Res. 2005, 43, 451–467. [Google Scholar] [CrossRef]
- Santos, J.M.; Silva, A.; Katopodis, C.; Pinheiro, P.; Pinheiro, A.; Bochechas, J.; Ferreira, M.T. Ecohydraulics of pool-type fishways: Getting past the barriers. Ecol. Eng. 2012, 48, 38–50. [Google Scholar] [CrossRef]
- Clay, C.H. Design of Fishways and Other Fish Facilities, 2nd ed.; Taylor & Francis Group: Oxfordshire, UK, 2017; p. 256. [Google Scholar] [CrossRef]
- Katopodis, C.; Williams, J.G. The development of fish passage research in a historical context. Ecol. Eng. 2012, 28, 407–417. [Google Scholar] [CrossRef]
- Nilsson, C.; Reidy, C.A.; Dynesius, M.; Revenga, C. Fragmentation and flow regulation of the world’s large river systems. Science 2005, 308, 405–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pegram, G.; Li, Y.; Le Quesne, T.; Speed, R.; Li, J.; Shen, F. River Basin Planning: Principles, Procedures and Approaches for Strategic Basin Planning; UNESCO: Paris, France, 2013; p. 181.
- Zarfl, C.; Lumsdon, A.E.; Berlekamp, J.; Tydecks, L.; Tockner, K. A global boom in hydropower dam construction. Aquat. Sci. 2015, 77, 161–170. [Google Scholar] [CrossRef]
- Romão, F.; Quaresma, A.L.; Santos, J.M.; Branco, P.; Pinheiro, A.N. Cyprinid passage performance in an experimental multislot fishway across distinct seasons. Mar. Freshw. Res. 2019, 70, 881–890. [Google Scholar] [CrossRef]
- FAO/Dvwk. Fish Passes—Design, Dimensions and Monitoring; FAO: Rome, Italy, 2002; p. 119. [Google Scholar]
- Valbuena-Castro, J.; Fuentes-Pérez, J.F.; García-Vega, A.; Bravo-Córdoba, F.J.; Ruiz-Legazpi, J.; de Azagra Paredes, A.M.; Sanz-Ronda, F.J. Coarse fishway assessment to prioritize retrofitting efforts: A case study in the Duero River basin. Ecol. Eng. 2020, 155, 105946. [Google Scholar] [CrossRef]
- Romão, F.; Quaresma, A.L.; Branco, P.; Santos, J.M.; Amaral, S.; Ferreira, M.T.; Katopodis, C.; Pinheiro, A.N. Passage performance of two cyprinids with different ecological traits in a fishway with distinct vertical slot configurations. Ecol. Eng. 2017, 108, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Fuentes-Pérez, J.F.; Sanz-Ronda, F.J.; de Martínez Azagra Paredes, A.; García-Vega, A. Modeling water-depth distribution in vertical-slot fishways under uniform and nonuniform scenarios. J. Hydraul. Eng. 2014, 140, 06014016. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Rajaratnam, N.; Zhu, D.Z. Mean flow and turbulence structure in vertical slot fishways. J. Hydraul. Eng. 2006, 132, 765–777. [Google Scholar] [CrossRef]
- Romão, F.; Branco, P.; Quaresma, A.L.; Amaral, S.D.; Pinheiro, A.N. Effectiveness of a multi-slot vertical slot fishway versus a standard vertical slot fishway for potamodromous cyprinids. Hydrobiologia 2018, 816, 153–163. [Google Scholar] [CrossRef]
- Mader, H.; Brandl, A.; Käfer, S. Design and Function Monitoring of an Enature® Vertical Slot Fish Pass in a Large Potamal River in Carinthia/Austria. Water 2020, 12, 551. [Google Scholar] [CrossRef] [Green Version]
- Cooke, S.J.; Hinch, S.G. Improving the reliability of fishway attraction and passage efficiency estimates to inform fishway engineering, science, and practice. Ecol. Eng. 2013, 58, 123–132. [Google Scholar] [CrossRef]
- Lucas, M.C.; Baras, E. Migration of Freshwater Fishes, 1st ed.; Blackwell Science Ltd.: Oxford, UK, 2001; p. 440. [Google Scholar]
- Thiem, J.D.; Dawson, J.W.; Hatin, D.; Danylchuk, A.J.; Dumont, P.; Gleiss, A.C.; Wilson, P.; Cooke, S.J. Swimming activity and energetic costs of adult lake sturgeon during fishway passage. J. Exp. Biol. 2016, 219, 2534–2544. [Google Scholar] [CrossRef] [Green Version]
- Lennox, R.J.; Paukert, C.; Aarestrup, K.; Auger-Méthé, M.; Baumgartner, L.; Birnie-Gauvin, K.; Davidsen, J.G. One hundred pressing questions on the future of global fish migration science, conservation, and policy. Front. Ecol. Evol. 2019, 7, 286. [Google Scholar] [CrossRef] [Green Version]
- Birnie-Gauvin, K.; Aarestrup, K.; Riis, T.M.; Jepsen, N.; Koed, A. Shining a light on the loss of rheophilic fish habitat in lowland rivers as a forgotten consequence of barriers, and its implications for management. Aquat. Conserv. 2017, 27, 1345–1349. [Google Scholar] [CrossRef]
- Santos, J.M.; Rivaes, R.; Boavida, I.; Branco, P. Structural microhabitat use by endemic cyprinids in a Mediterranean-type river: Implications for restoration practices. Aquatic Conserv Mar Freshw. Ecosyst. 2018, 28, 26–36. [Google Scholar] [CrossRef]
- Boix, D.; García-Berthou, E.; Gascón, S.; Benejam, L.; Tornés, E.; Sala, J.; Benito, J.; Munné, A.; Solà, C.; Sabater, S. Response of community structure to sustained drought in Mediterranean rivers. J. Hydrol. 2010, 383, 135–146. [Google Scholar] [CrossRef]
- Kingsford, R.T. Conservation management of rivers and wetlands under climate change—A synthesis. Mar. Freshwater Res. 2011, 62, 217–222. [Google Scholar] [CrossRef]
- Silva, A.T.; Lucas, M.C.; Castro-Santos, T.; Katopodis, C.; Baumgartner, L.J.; Thiem, J.D.; Aaretrup, K.; Pompeu, P.S.; O’Brien, G.C.; Braun, D.C.; et al. The future of fish passage science, engineering, and practice. Fish Fish. 2017, 19, 340–362. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.G.; Darwall, W.R. The status and distribution of freshwater fish endemic to the Mediterranean Basin. In IUCN Red list of Threatened Species; IUCN: Gland, Switzerland; Cambridge, UK, 2006; Volume 1, 34p. [Google Scholar]
- Miranda, R.; Pino-del-Carpio, A. Analyzing freshwater fish biodiversity records and respective conservation areas in Spain. J. Appl. Ichthyol. 2016, 32, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Branco, P.; Segurado, P.; Santos, J.M.; Pinheiro, P.J.; Ferreira, M.T. Does longitudinal connectivity loss affect the distribution of freshwater fish? Ecol. Eng. 2012, 48, 70–78. [Google Scholar] [CrossRef]
- Tummers, J.S.; Hudson, S.; Lucas, M.C. Evaluating the effectiveness of restoring longitudinal connectivity for stream fish communities: Towards a more holistic approach. Sci. Total Environ. 2016, 569, 850–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajaratnam, N.; Katopodis, C.; Solanki, S. New designs for vertical slot fishways. Can. J. Civ. Eng. 1992, 19, 402–414. [Google Scholar] [CrossRef]
- Tauber, M.; Mader, H. Development of an Economical and Ecological Optimized Multi Slot Fish Pass. In Proceedings of the Small Hydro 2009, Vancouver, BC, Canada, 28–29 April 2009. [Google Scholar]
- Tauber, M.; Mader, H. Hydraulic comparison of standard vertical slot and multi structure slot fish bypass. In Proceedings of the 1st European IAHR Congress, Edinburgh, UK, 4–6 May 2010. [Google Scholar]
- Quaresma, A.L.; Romão, F.; Branco, P.; Ferreira, M.T.; Pinheiro, A.N. Multi slot versus single slot pool-type fishways: A modelling approach to compare hydrodynamics. Ecol. Eng. 2018, 122, 197–206. [Google Scholar] [CrossRef]
- Fuentes-Pérez, J.F.; Tuhtan, J.A.; Eckert, M.; Romão, F.; Ferreira, M.T.; Kruusmaa, M.; Branco, P. Hydraulics of Vertical-Slot Fishways: Nonuniform Profiles. J. Hydraul. Eng. 2019, 145, 06018020. [Google Scholar] [CrossRef] [Green Version]
- [CEN] European Committee for Standardization. Water Quality: Sampling of Fish with Electricity; European Standard EN 14011: 2003 E; CEN: Brussels, Belgium, 2003. [Google Scholar]
- Plaut, I. Critical swimming speed: Its ecological relevance. Comp. Biochem. Physiol. A 2001, 131, 41–50. [Google Scholar] [CrossRef]
- Amaral, S.D.; Branco, P.; Romão, F.; Viseu, T.; Ferreira, M.T.; Pinheiro, A.N.; Santos, J.M. The effect of weir crest width and discharge on passage performance of a potamodromous cyprinid. Mar. Freshw. Res. 2018, 69, 1795–1804. [Google Scholar] [CrossRef]
- Mallen-Cooper, M. Swimming ability of adult golden perch, Macquaria ambigua (Percichthyidae), and adult silver perch, Bidyanus bidyanus (Teraponidae), in an experimental vertical-slot fishway aust. J. Mar. Freshw. Res. 1994, 45, 191–198. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA); Statistics Reference Online; Wiley StatsRef, John Wiley & Sons: Hoboken, NJ, USA, 2017. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ For PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Linnansaari, T.; Wallace, B.; Curry, R.; Yamazaki, G. Mactaquac Aquatic Ecosystem Study Report Series 2015-016, Fish Passage in Large Rivers: A literature Review; Canadian Rivers Institute: New Brunswick, CA, Canada, 2015. [Google Scholar]
- Sanz-Ronda, F.J.; Bravo-Córdoba, F.J.; Sánchez-Pérez, A.; García-Vega, A.; Valbuena-Castro, J.; Fernandes-Celestino, L.; Torralva, M.; Oliva-Paterna, F.J. Passage Performance of Technical Pool-Type Fishways for Potamodromous Cyprinids: Novel Experiences in Semiarid Environments. Water 2019, 11, 2362. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.G.; Amstrong, G.; Katopodis, C.; Larinier, M.; Travade, F. Thinking like a fish: A key ingredient for development of effective fish passage facilities at river obstructions. River Res. Appl. 2012, 28, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Baumgartner, L.J.; Boys, C.; Marsden, T.; McPherson, J.; Ning, N.; Phonekhampheng, O.; Robinson, W.; Singhanouvong, D.; Stuart, I.G.; Thorncraft, G. A Cone Fishway Facilitates Lateral Migrations of Tropical River-Floodplain Fish Communities. Water 2020, 12, 513. [Google Scholar] [CrossRef] [Green Version]
- Tamario, C.; Degerman, E.; Donadi, S.; Spjut, D.; Sandin, L. Nature-like fishways as compensatory lotic habitats. River Res Applic. 2018, 34, 253–261. [Google Scholar] [CrossRef]
- Cea, L.; Pena, L.; Puertas, J.; Vazquez-Cendon, M.E.; Pena, E. Application of several depth-averaged turbulence models to simulate flow in vertical slot fishways. J. Hydraul. Eng. 2007, 133, 160–172. [Google Scholar] [CrossRef]
- Domenici, P.; Kapoor, B.G. Fish Locomotion: An Eco-Ethological Perspective, 1st ed.; Taylor & Francis Group: Boca Raton, FL, USA, 2010; p. 549. [Google Scholar] [CrossRef]
- Elder, J.; Coombs, S. The influence of turbulence on the sensory basis of rheotaxis. J. Com. Physiol. A 2015, 201, 667–680. [Google Scholar] [CrossRef]
- Branco, P.; Santos, J.M.; Katopodis, C.; Pinheiro, A.; Ferreira, M.T. Pool-type fishways: Two different morpho-ecological cyprinid species facing plunging and streaming flows. PLoS ONE 2013, 8, e65089. [Google Scholar] [CrossRef]
- Bermúdez, M.; Puertas, J.; Cea, L.; Pena, L.; Balairón, L. Influence of pool geometry on the biological efficiency of vertical slot fishways. Ecol. Eng. 2010, 36, 1355–1364. [Google Scholar] [CrossRef]
- Mateus, C.S.; Quintella, B.R.; Almeida, P.R. The critical swimming speed of Iberian barbel Barbus bocagei in relation to size and sex. J. Fish Biol. 2008, 73, 1783–1789. [Google Scholar] [CrossRef]
- Romão, F.; Quintella, B.R.; Pereira, T.J.; Almeida, P.R. Swimming performance of two Iberian cyprinids: The Tagus nase Pseudochondrostoma polylepis (Steindachner, 1864) and the bordallo Squalius carolitertii (Doadrio, 1988). J. Appl. Ichthyol. 2012, 28, 26–30. [Google Scholar] [CrossRef]
- Liao, J.C. A review of fish swimming mechanics and behavior in altered flows. Philos. Trans. R. Soc. B 2007, 362, 1973–1993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, C. Fatigue and exercise tests with fish. Comp. Biochem. Physiol. A 1995, 112, 1–20. [Google Scholar] [CrossRef]
- Mallen-Cooper, M.; Zampatti, B.; Stuart, I.; Baumgartner, L. Innovative fishways—Manipulating turbulence in the vertical-slot design to improve performance and reduce cost. In Report to the Murray-Darling Basin Commission by Fishway Consulting Services; Murray Darling Basin Authority: Sydney, Australia, 2008. [Google Scholar]
- Field, C.B.; Barros, V.R.; Mach, K.; Mastrandrea, M. Climate Change 2014: Impacts, Adaptation, and Vulnerability. In Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Q (L·s−1) | Pv (W·m−3) | Cd (-) | L (m) | B (m) | b (m) | ΔH (m) | hm (m) | i (%) | |
|---|---|---|---|---|---|---|---|---|---|
| VSF 1 | 112 | 114 | 0.71 | 1.85 | 1.00 | 0.10 | 0.16 | 0.8 | 8.5 |
| VSF 2 | 80 | 83 | 0.52 | ||||||
| MSF 1 | 58 | 59 | 0.53 | ||||||
| MSF 2 | 37 | 38 | 0.34 |
| Fishway Configuration | VSF 1 | VSF 2 | MSF 1 | MSF 2 |
|---|---|---|---|---|
| Q (L·s−1) | 112 | 80 | 58 | 37 |
| U average (m·s−1) | 0.58 | 0.35 | 0.26 | 0.19 |
| U max (m·s−1) | 2.2 | 1.9 | 1.4 | 1.4 |
| TKE average (m2·s−2) | 0.054 | 0.042 | 0.026 | 0.016 |
| TKE max (m2·s−2) | 0.34 | 0.35 | 0.12 | 0.18 |
| RSS average (Pa) | 10.3 | 8.2 | 5.1 | 3.4 |
| RSS max (Pa) | 145 | 147 | 52 | 55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romão, F.; Quaresma, A.L.; Santos, J.M.; Amaral, S.D.; Branco, P.; Pinheiro, A.N. Multislot Fishway Improves Entrance Performance and Fish Transit Time over Vertical Slots. Water 2021, 13, 275. https://doi.org/10.3390/w13030275
Romão F, Quaresma AL, Santos JM, Amaral SD, Branco P, Pinheiro AN. Multislot Fishway Improves Entrance Performance and Fish Transit Time over Vertical Slots. Water. 2021; 13(3):275. https://doi.org/10.3390/w13030275
Chicago/Turabian StyleRomão, Filipe, Ana L. Quaresma, José M. Santos, Susana D. Amaral, Paulo Branco, and António N. Pinheiro. 2021. "Multislot Fishway Improves Entrance Performance and Fish Transit Time over Vertical Slots" Water 13, no. 3: 275. https://doi.org/10.3390/w13030275
APA StyleRomão, F., Quaresma, A. L., Santos, J. M., Amaral, S. D., Branco, P., & Pinheiro, A. N. (2021). Multislot Fishway Improves Entrance Performance and Fish Transit Time over Vertical Slots. Water, 13(3), 275. https://doi.org/10.3390/w13030275