
Population Dynamics, Fishery, and Exploitation Status of Norway Lobster (Nephrops norvegicus) in Eastern Mediterranean



Abstract
:1. Introduction
2. Materials and Methods
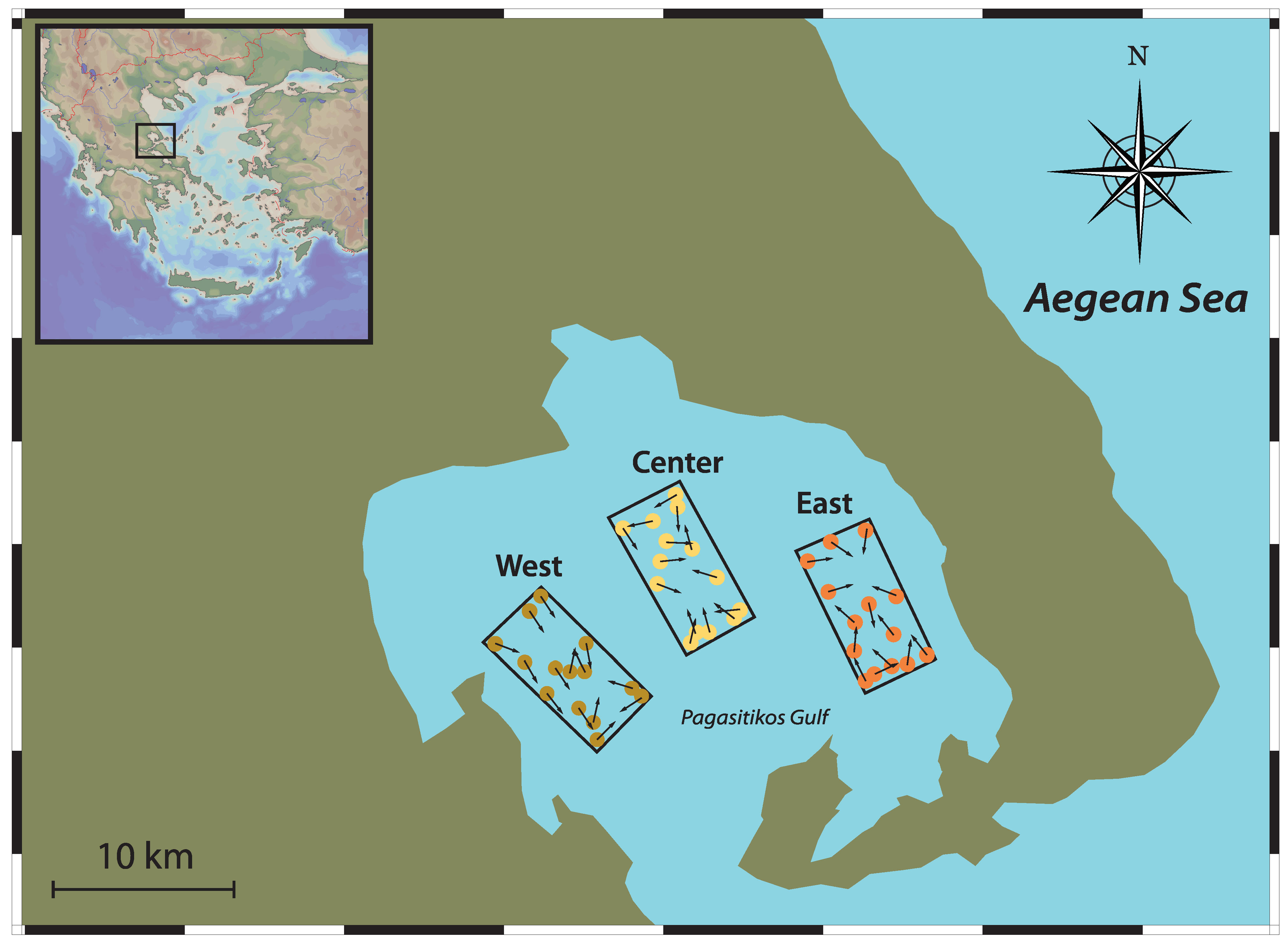
2.1. Sampling Area
2.2. Trawl Survey
2.3. Measurements
2.4. Data Analysis
3. Results
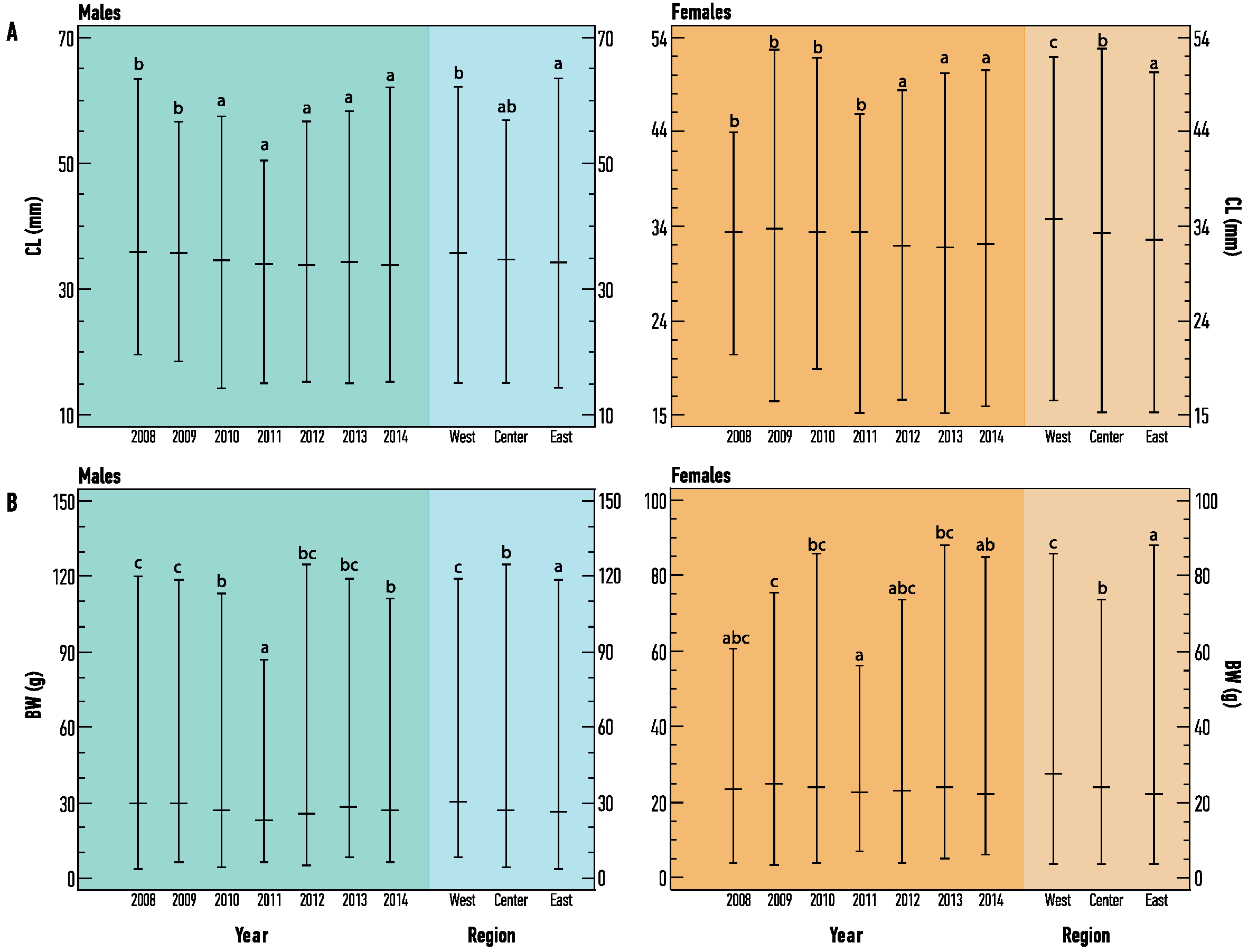
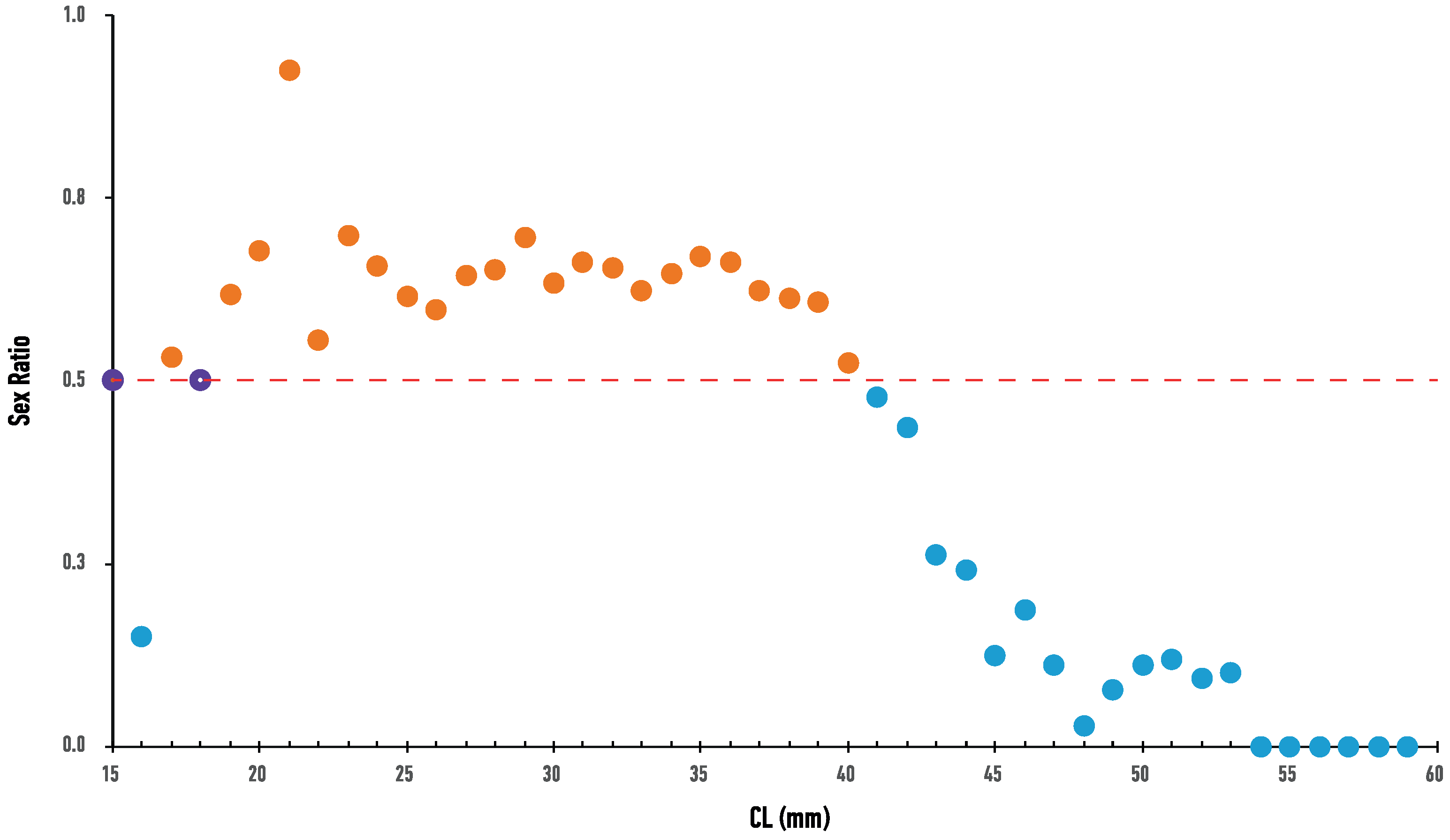
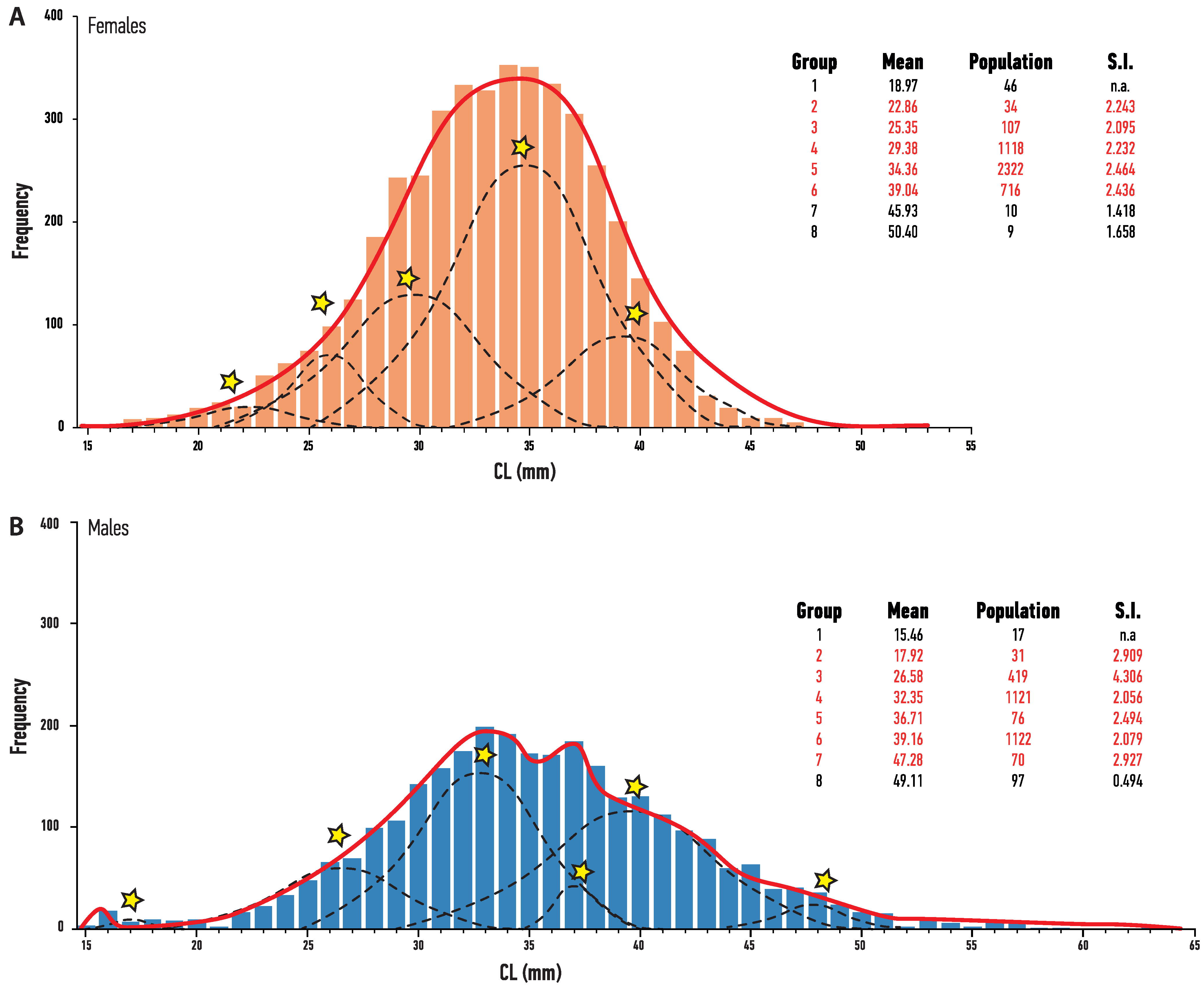
3.1. Population Structure
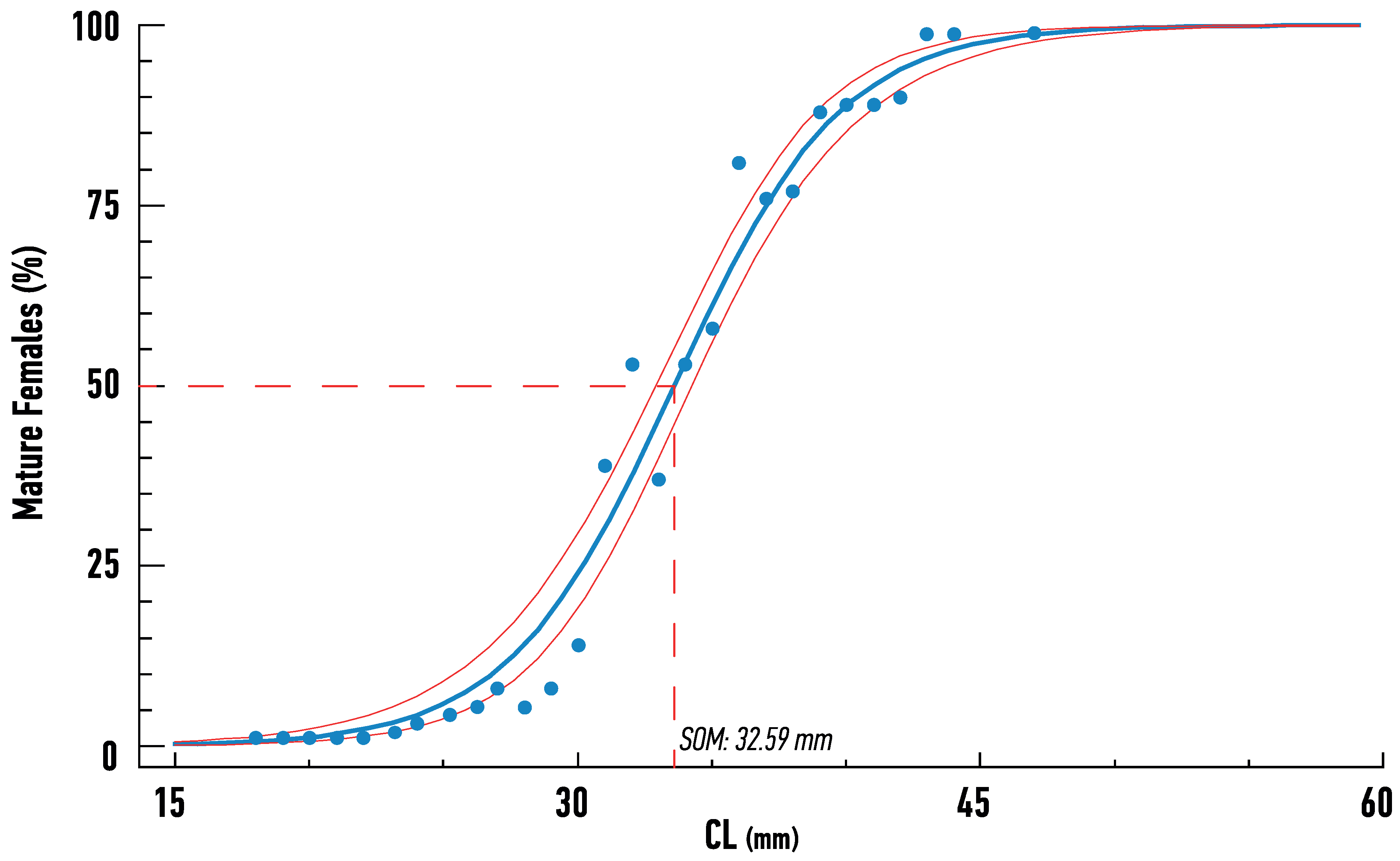
3.2. Age Determination and Maturity
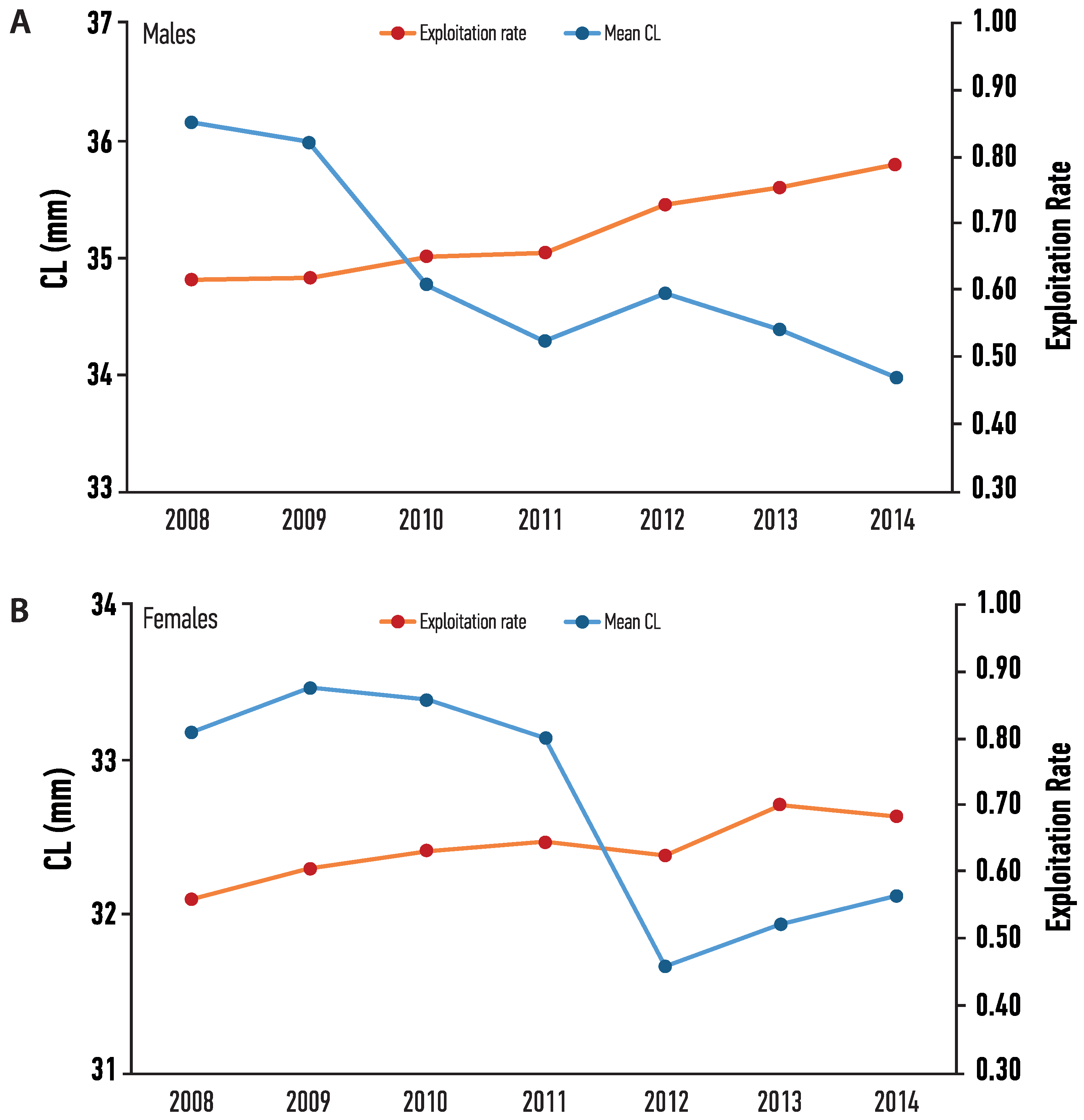
3.3. Growth Parameters and Mortality
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Johnson, M.P.; Lordan, C.; Power, A.M. Habitat and Ecology of Nephrops norvegicus. In Advances in Marine Biology; Johnson, M.L., Johnson, M.P., Eds.; Academic Press: Cambridge, MA, USA, 2013; Volume 64, pp. 27–63. [Google Scholar]
- Bell, M.; Tuck, I.D.; Dobby, H. Nephrops Species. In Lobsters: Biology, Management, Aquaculture and Fisheries; Phillips, B.F., Ed.; Wiley: Hoboken, NJ, USA, 2013; pp. 357–413. [Google Scholar]
- Ungfors, A.; Bell, E.; Johnson, M.L.; Cowing, D.; Dobson, N.C.; Bublitz, R.; Sandell, J. Nephrops Fisheries in European Waters. In Advances in Marine Biology; Johnson, M.L., Johnson, M.P., Eds.; Academic Press: Cambridge, MA, USA, 2013; Volume 64, pp. 247–314. [Google Scholar]
- FAO. Fishfinder-Web Site. FAO FishFinder Contacts. FI Institutional Websites; FAO Fisheries and Aquaculture Department: Rome, Italy, 2021; Available online: www.fao.org/fishery/ (accessed on 13 January 2021).
- Carpenter, G.; Kleinjans, R.; Villasante, S.; O’Leary, B.C. Landing the blame: The influence of EU Member States on quota setting. Mar. Policy 2016, 64, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Colloca, F.; Cardinale, M.; Maynou, F.; Giannoulaki, M.; Scarcella, G.; Jenko, K.; Bellido, J.M.; Fiorentino, F. Rebuilding Mediterranean fisheries: A new paradigm for ecological sustainability. Fish Fish. 2011, 14, 89–109. [Google Scholar] [CrossRef]
- Sardà, F.; Lleonart, J.; Cartes, J.E. An analysis of the population dynamics of Nephrops norvegicus (L.) in the Mediterranean Sea. Sci. Mar. 1998, 62, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Ligas, A.; Sartor, P.; Colloca, F. Trends in population dynamics and fishery of Parapenaeus longirostris and Nephrops norvegicus in the Tyrrhenian Sea (NW Mediterranean): The relative importance of fishery and environmental variables. Mar. Ecol. 2011, 32, 25–35. [Google Scholar] [CrossRef]
- Cosgrove, R.; Browne, D.; Minto, C.; Tyndall, P.; Oliver, M.; Montgomerie, M.; McHugh, M. A game of two halves: Bycatch reduction in Nephrops mixed fisheries. Fish. Res. 2019, 210, 31–40. [Google Scholar] [CrossRef]
- Farmer, A.S.D. Reproduction in Nephrops norvegicus (Decapoda: Nephropidae). J. Zoöl. 1974, 174, 161–183. [Google Scholar] [CrossRef]
- Sardà, F. Reproduction and Moult Synchronism in Nephrops norvegicus (L.) (Decapoda, Nephropidae) in the Western Mediterranean: Is Spawning Annual or Biennial? Crustaceana 1991, 60, 186–199. [Google Scholar] [CrossRef]
- Tuck, I.; Chapman, C.J.; Atkinson, R.J.A. Population biology of the Norway lobster, Nephrops norvegicus(L.) in the Firth of Clyde, Scotland—I: Growth and density. ICES J. Mar. Sci. 1997, 54, 125–135. [Google Scholar] [CrossRef]
- Mytilineou, C.; Castro, M.; Gancho, P.; Fourtouni, A. Growth studies on Norway lobster, Nephrops norvegicus (L.), in different areas of the Mediterranean Sea and the adjacent Atlantic. Sci. Mar. 1998, 62, 43–60. [Google Scholar] [CrossRef]
- Ulmestrand, M.; Eggert, H. Growth of Norway lobster, Nephrops norvegicus (Linnaeus 1758), in the Skagerrak, estimated from tagging experiments and length frequency data. ICES J. Mar. Sci. 2001, 58, 1326–1334. [Google Scholar] [CrossRef] [Green Version]
- Stamatis, C.; Triantafyllidis, A.; Moutou, K.A.; Mamuris, Z. Mitochondrial DNA variation in Northeast Atlantic and Mediterranean populations of Norway lobster, Nephrops norvegicus. Mol. Ecol. 2004, 13, 1377–1390. [Google Scholar] [CrossRef] [PubMed]
- McQuaid, N.; Briggs, R.; Roberts, D. Estimation of the size of onset of sexual maturity in Nephrops norvegicus (L.). Fish. Res. 2006, 81, 26–36. [Google Scholar] [CrossRef]
- Ridgway, I.; Taylor, A.; Atkinson, R.; Chang, E.; Neil, D. Impact of capture method and trawl duration on the health status of the Norway lobster, Nephrops norvegicus. J. Exp. Mar. Biol. Ecol. 2006, 339, 135–147. [Google Scholar] [CrossRef]
- Bell, M.; Elson, J.; Addison, J.; Revill, A.; Bevan, D. Trawl catch composition in relation to Norway lobster (Nephrops norvegicus L.) abundance on the Farn Deeps grounds, NE England. Fish. Res. 2008, 90, 128–137. [Google Scholar] [CrossRef]
- Ziegler, F.; Valentinsson, D. Environmental life cycle assessment of Norway lobster (Nephrops norvegicus) caught along the Swedish west coast by creels and conventional trawls—LCA methodology with case study. Int. J. Life Cycle Assess. 2008, 13, 487–497. [Google Scholar] [CrossRef]
- Ayza, O.; Tuset, V.M.; González, J.A. Estimation of size at onset of sexual maturity and growth parameters in Norway lobster (Nephrops norvegicus) off the Portuguese coast. Fish. Res. 2011, 108, 205–208. [Google Scholar] [CrossRef]
- Murray, F.; Cowie, P.R. Plastic contamination in the decapod crustacean Nephrops norvegicus (Linnaeus, 1758). Mar. Pollut. Bull. 2011, 62, 1207–1217. [Google Scholar] [CrossRef]
- Leocádio, A.M.; Whitmarsh, D.; Castro, M. Comparing Trawl and Creel Fishing for Norway Lobster (Nephrops norvegicus): Biological and Economic Considerations. PLoS ONE 2012, 7, e39567. [Google Scholar] [CrossRef]
- Eiríksson, H. Reproductive Biology of Female Norway Lobster, Nephrops norvegicus (Linnaeus, 1758) Leach, in Icelandic Waters During the Period 1960–2010: Comparative Overview of Distribution Areas in the Northeast Atlantic and the Mediterranean. In Advances in Marine Biology; Lesser, M., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 68, pp. 65–210. [Google Scholar]
- Wood, H.; Eriksson, S.; Nordborg, M.; Styf, H. The effect of environmental stressors on the early development of the Norway lobster Nephrops norvegicus (L.). J. Exp. Mar. Biol. Ecol. 2015, 473, 35–42. [Google Scholar] [CrossRef]
- Haynes, P.S.; Browne, P.; Fullbrook, L.; Graham, C.T.; Hancox, L.; Johnson, M.P.; Lauria, V.; Power, A.M. Growth in Nephrops norvegicus from a tag-recapture experiment. Sci. Rep. 2016, 6, 35143. [Google Scholar] [CrossRef] [Green Version]
- Méhault, S.; Morandeau, F.; Kopp, D. Survival of discarded Nephrops norvegicus after trawling in the Bay of Biscay. Fish. Res. 2016, 183, 396–400. [Google Scholar] [CrossRef] [Green Version]
- Becker, C.; Cunningham, E.M.; Dick, J.T.A.; Eagling, L.E.; Sigwart, J.D. A unified scale for female reproductive stages in the Norway lobster (Nephrops norvegicus): Evidence from macroscopic and microscopic characterization. J. Morphol. 2018, 279, 1700–1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Power, A.M.; Merder, J.; Browne, P.; Freund, J.A.; Fullbrook, L.; Graham, C.; Kennedy, R.J.; O’Carroll, J.P.J.; Wieczorek, A.M.; Johnson, M.P. Field-recorded data on habitat, density, growth and movement of Nephrops norvegicus. Sci. Data 2019, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, J.; Finarelli, J.A.; Jonasson, J.P.; Carlsson, J. Mitochondrial D-loop DNA analyses of Norway lobster (Nephrops norvegicus) reveals genetic isolation between Atlantic and East Mediterranean populations. J. Mar. Biol. Assoc. UK 2019, 99, 933–940. [Google Scholar] [CrossRef]
- Mytilineou, C.; Sardà, F. Age and growth of Nephrops norvegicus in the Catalan Sea, using length-frequency analysis. Fish. Res. 1995, 23, 283–299. [Google Scholar] [CrossRef]
- Maynou, F.; Sardà, F. Nephrops norvegicus population and morphometrical characteristics in relation to substrate heterogeneity. Fish. Res. 1997, 30, 139–149. [Google Scholar] [CrossRef]
- Bianchini, M.L.; Di Stefano, L.; Ragonese, S. Size and age at onset of sexual maturity of female Norway lobster Nephrops norvegicus L. (Crustacea: Nephropidae) in the Strait of Sicily (Central Mediterranean Sea). Sci. Mar. 1998, 62, 151–159. [Google Scholar] [CrossRef]
- Mori, M.; Biagi, F.; De Ranieri, S. Fecundity and egg loss during incubation in Norway lobster (Nephrops norvegicus) in the North Tyrrhenian Sea. J. Nat. Hist. 1998, 32, 1641–1650. [Google Scholar] [CrossRef]
- Sardà, F. Growth parameters of deep-water decapod crustaceans in the Northwestern Mediterranean Sea: A comparative approach. Mar. Biol. 2000, 136, 79–90. [Google Scholar] [CrossRef]
- Abelló, P.; Abella, Á.; Adamidou, A.; Jukic-Peladic, S.; Maiorano, P.; Spedicato, M.T. Geographical patterns in abundance and population structure of Nephrops norvegicus and Parapenaeus longirostris (Crustacea: Decapoda) along the European Mediterranean coasts. Sci. Mar. 2002, 66, 125–141. [Google Scholar] [CrossRef] [Green Version]
- Morello, E.B.; Antolini, B.; Gramitto, M.E.; Atkinson, R.J.A.; Froglia, C. The fishery for Nephrops norvegicus (Linnaeus, 1758) in the central Adriatic Sea (Italy): Preliminary observations comparing bottom trawl and baited creels. Fish. Res. 2009, 95, 325–331. [Google Scholar] [CrossRef]
- Brčić, J.; Herrmann, B.; Mašanović, M.; Baranović, M.; Šifner, S.K.; Škeljo, F. Size selection of Nephrops norvegicus (L.) in commercial creel fishery in the Mediterranean Sea. Fish. Res. 2018, 200, 25–32. [Google Scholar] [CrossRef] [Green Version]
- García-De-Vinuesa, A.; Breen, M.; Benoît, H.P.; Maynou, F.; Marquès, M. Seasonal variation in the survival of discarded Nephrops norvegicus in a NW Mediterranean bottom-trawl fishery. Fish. Res. 2020, 230, 105671. [Google Scholar] [CrossRef]
- Bertrand, J.A.; Gil de Sola, L.; Papaconstantinou, C.; Relini, G.; Souplet, A. The general specifications of the MEDITS surveys. Sci. Mar. 2002, 66, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.J.; Papadopoulou, K.-N. Burrow density and stock size fluctuations of Nephrops norvegicus in a semi-enclosed bay. ICES J. Mar. Sci. 2003, 60, 798–805. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.J.; Marrs, S.; Atkinson, R.; Papadopoulou, K.; Hills, J. Underwater television for fisheries-independent stock assessment of Nephrops norvegicus from the Aegean (eastern Mediterranean) Sea. Mar. Ecol. Prog. Ser. 2003, 256, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Mente, E.; Karapanagiotidis, I.T.; Logothetis, P.; Vafidis, D.; Malandrakis, E.; Neofitou, N.; Exadactylos, A.; Stratakos, A. The reproductive cycle of Norway lobster. J. Zoöl. 2009, 278, 324–332. [Google Scholar] [CrossRef]
- Aydin, I.; Aydin, C. Length-Length and Length-Weight Relationships in Nephrops norvegicus from the Aegean Sea (Linnaeus, 1758). Mediterr. Mar. Sci. 2011, 12, 121. [Google Scholar] [CrossRef] [Green Version]
- Stathopoulou, E.; Apostologamvrou, C.; Vafidis, D.; Mente, E.; Berillis, P. Sex differentiation in the histology of the digestive gland of Nephrops norvegicus from Pagasitikos Gulf during spring. Aquat. Living Resour. 2017, 30, 28. [Google Scholar] [CrossRef]
- Kevrekidis, K.; Thessalou-Legaki, M. Catch rates, size structure and sex ratio of Melicertus kerathurus (Decapoda: Penaeidae) from an Aegean Sea trawl fishery. Fish. Res. 2006, 80, 270–279. [Google Scholar] [CrossRef]
- Vafidis, D.; Leontarakis, P.K.; Dailianis, T.; Kallianiotis, A. Population characteristics of four deep-water pandalid shrimps (Decapoda: Caridea) in the northern Aegean Sea (NE Mediterranean). J. Nat. Hist. 2008, 42, 2079–2093. [Google Scholar] [CrossRef]
- Kevrekidis, K.; Thessalou-Legaki, M. Population dynamics of Melicertus kerathurus (Decapoda: Penaeidae) in Thermaikos Gulf (N. Aegean Sea). Fish. Res. 2011, 107, 46–58. [Google Scholar] [CrossRef]
- Lolas, A.; Vafidis, D. Population dynamics of two caprellid species (Crustaceae: Amphipoda: Caprellidae) from shallow hard bottom assemblages. Mar. Biodivers. 2013, 43, 227–236. [Google Scholar] [CrossRef]
- Konstantinidis, I.; Gkafas, G.; Karamitros, G.; Lolas, A.; Antoniadou, C.; Vafidis, D.; Exadactylos, A. Population structure of two benthic species with different larval stages in the eastern Mediterranean Sea. J. Environ. Prot. Ecol. 2017, 18, 930–939. [Google Scholar]
- Lolas, A.; Antoniadou, C.; Vafidis, D. Spatial variation of molluscan fauna associated with Cystoseira assemblages from a semi-enclosed gulf in the Aegean Sea. Reg. Stud. Mar. Sci. 2018, 19, 17–24. [Google Scholar] [CrossRef]
- Klaoudatos, D.; Kotsiri, Z.; Neofitou, N.; Lolas, A.; Vafidis, D. Population Characteristics of the Mid-Littoral Chthamalid Barnacle C. stellatus (Poli, 1791) in Eastern Mediterranean (Central Greece). Water 2020, 12, 3304. [Google Scholar] [CrossRef]
- Tuck, I.; Atkinson, R.J.A.; Chapman, C.J. Population biology of the Norway lobster, Nephrops norvegicus (L.) in the Firth of Clyde, Scotland II: Fecundity and size at onset of sexual maturity. ICES J. Mar. Sci. 2000, 57, 1227–1239. [Google Scholar] [CrossRef]
- Powell, A.; Eriksson, S.P. Reproduction: Life Cycle, Larvae and Larviculture. In Advances in Marine Biology; Johnson, M.L., Johnson, M.P., Eds.; Academic Press: Cambridge, MA, USA, 2013; Volume 64, pp. 201–245. [Google Scholar]
- Sánchez-Lizaso, J.L.; Goñi, R.; Reñones, O.; Charton, J.G.; Galzin, R.; Bayle, J.; Sanchez-Jerez, P.; Pérez-Ruzafa, A.; Ramos, A. Density dependence in marine protected populations: A review. Environ. Conserv. 2000, 27, 144–158. [Google Scholar] [CrossRef] [Green Version]
- Relini, L.O.; Zamboni, A.; Fiorentino, F.; Massi, D. La reproducción de la cigala (Nephrops norvegicus, Crustacea Decapoda Nephropidae) en diferentes zonas del Mediterráneo. Sci. Mar. 1998, 62, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Castro, M. A Methodology for Obtaining Information on the Age Structure and Growth Rates of the Norway Lobster, Nephrops Nor Vegicus (L.) (Decapoda, Nephropoidea). Crustaceana 1992, 63, 29–43. [Google Scholar] [CrossRef]
- Angelini, S.; Martinelli, M.; Santojanni, A.; Colella, S. Biological evidence of the presence of different subpopulations of Norway lobster (Nephrops norvegicus) in the Adriatic Sea (Central Mediterranean Sea). Fish. Res. 2020, 221, 105365. [Google Scholar] [CrossRef]
- Vogt, G. Ageing and longevity in the Decapoda (Crustacea): A review. Zoöl. Anz. A J. Comp. Zoöl. 2012, 251, 1–25. [Google Scholar] [CrossRef]
- Pauly, D. Beyond duplicity and ignorance in global fisheries. Sci. Mar. 2009, 73, 215–224. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
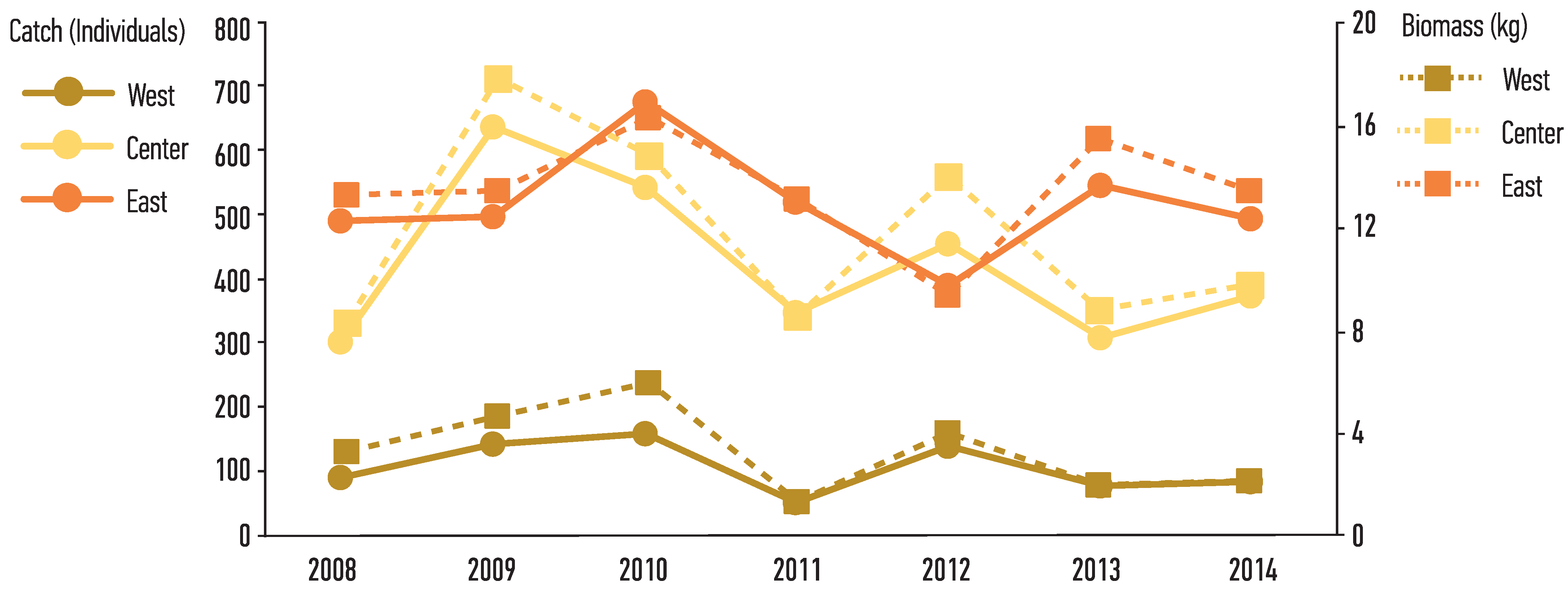{kind=link}
{kind=link}
{kind=link}
{kind=link}
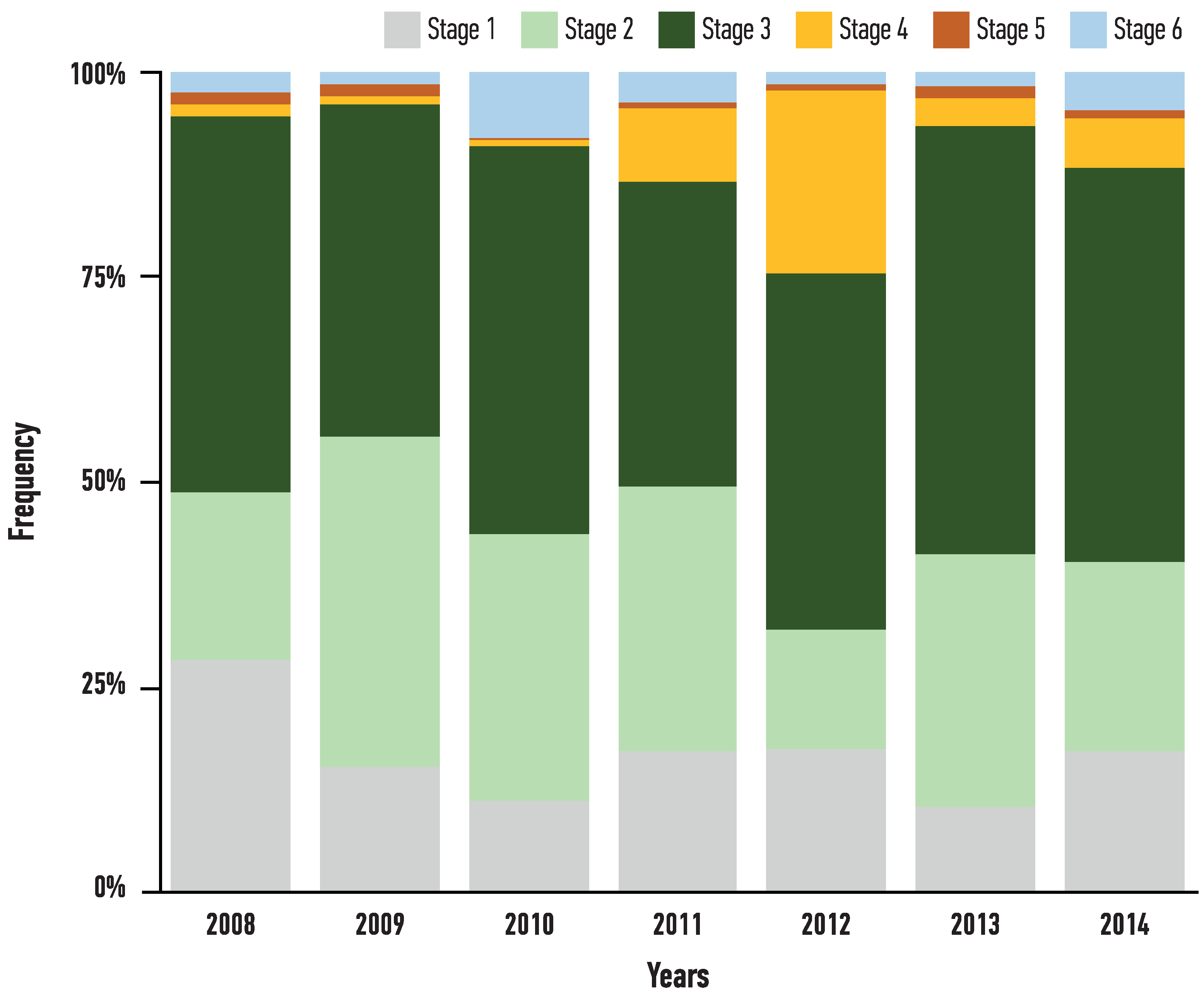{kind=link}
{kind=link}
{kind=link}
| Year | a | b | r | F | p |
|---|---|---|---|---|---|
| Males | |||||
| 2008 | −3.023 ± 0.12 | 2.893 ± 0.07 | 0.898 | 0.83 | 0.7743 |
| 2009 | −3.348 ± 0.08 | 3.100 ± 0.05 | 0.941 | 0.61 | 0.9873 |
| 2010 | −3.105 ± 0.08 | 2.939 ± 0.05 | 0.924 | 2.12 | 0.0003 * |
| 2011 | −2.089 ± 0.12 | 2.269 ± 0.08 | 0.801 | 1.44 | 0.1064 |
| 2012 | −2.565 ± 0.09 | 2.604 ± 0.06 | 0.901 | 2.82 | 0.1002 |
| 2013 | −1.530 ± 0.10 | 1.956 ± 0.07 | 0.827 | 1.21 | 0.2648 |
| 2014 | −1.865 ± 0.10 | 2.158 ± 0.06 | 0.857 | 1.66 | 0.0505 |
| Females | |||||
| 2008 | −3.252 ± 0.08 | 3.101 ± 0.05 | 0.933 | 1.35 | 0.0533 |
| 2009 | −2.975 ± 0.07 | 2.851 ± 0.05 | 0.901 | 1.90 | <0.0001 * |
| 2010 | −3.342 ± 0.05 | 3.089 ± 0.03 | 0.956 | 1.09 | 0.2307 |
| 2011 | −1.555 ± 0.11 | 1.913 ± 0.07 | 0.779 | 1.49 | 0.0200 * |
| 2012 | −2.193 ± 0.09 | 2.365 ± 0.06 | 0.858 | 2.36 | <0.0001 * |
| 2013 | −1.988 ± 0.08 | 2.234 ± 0.06 | 0.866 | 1.49 | 0.0143 * |
| 2014 | −2.094 ± 0.09 | 2.285 ± 0.06 | 0.861 | 1.27 | 0.0935 |
| Year | CLinf (mm) | K (year−1) | t0 (year) | Z (year−1) | M (year−1) | F (year−1) | E |
|---|---|---|---|---|---|---|---|
| Males | |||||||
| 2008 | 83.62 | 0.11 | −2.84 | 0.798 | 0.306 | 0.492 | 0.62 |
| 2009 | 79.38 | 0.10 | −2.57 | 0.771 | 0.293 | 0.478 | 0.62 |
| 2010 | 77.14 | 0.09 | −1.66 | 0.690 | 0.241 | 0.449 | 0.65 |
| 2011 | 74.23 | 0.11 | −1.18 | 0.698 | 0.239 | 0.460 | 0.66 |
| 2012 | 78.99 | 0.11 | −0.59 | 0.723 | 0.196 | 0.527 | 0.73 |
| 2013 | 80.95 | 0.09 | −0.77 | 0.704 | 0.174 | 0.531 | 0.75 |
| 2014 | 79.90 | 0.13 | −0.13 | 0.768 | 0.161 | 0.606 | 0.79 |
| Females | |||||||
| 2008 | 66.91 | 0.14 | −1.93 | 0.783 | 0.346 | 0.437 | 0.56 |
| 2009 | 69.55 | 0.18 | −2.19 | 0.854 | 0.337 | 0.518 | 0.61 |
| 2010 | 68.19 | 0.12 | −1.46 | 0.726 | 0.269 | 0.457 | 0.63 |
| 2011 | 64.83 | 0.18 | −2.72 | 0.767 | 0.273 | 0.494 | 0.64 |
| 2012 | 67.48 | 0.14 | −2.98 | 0.813 | 0.308 | 0.506 | 0.62 |
| 2013 | 69.12 | 0.11 | −1.25 | 0.792 | 0.238 | 0.554 | 0.70 |
| 2014 | 68.76 | 0.13 | −0.51 | 0.721 | 0.228 | 0.493 | 0.68 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lolas, A.; Vafidis, D. Population Dynamics, Fishery, and Exploitation Status of Norway Lobster (Nephrops norvegicus) in Eastern Mediterranean. Water 2021, 13, 289. https://doi.org/10.3390/w13030289
Lolas A, Vafidis D. Population Dynamics, Fishery, and Exploitation Status of Norway Lobster (Nephrops norvegicus) in Eastern Mediterranean. Water. 2021; 13(3):289. https://doi.org/10.3390/w13030289
Chicago/Turabian StyleLolas, Alexios, and Dimitris Vafidis. 2021. "Population Dynamics, Fishery, and Exploitation Status of Norway Lobster (Nephrops norvegicus) in Eastern Mediterranean" Water 13, no. 3: 289. https://doi.org/10.3390/w13030289
APA StyleLolas, A., & Vafidis, D. (2021). Population Dynamics, Fishery, and Exploitation Status of Norway Lobster (Nephrops norvegicus) in Eastern Mediterranean. Water, 13(3), 289. https://doi.org/10.3390/w13030289