
Scientific Methods to Understand Fish Population Dynamics and Support Sustainable Fisheries Management

 , ,
, , 
Abstract
:1. Introduction
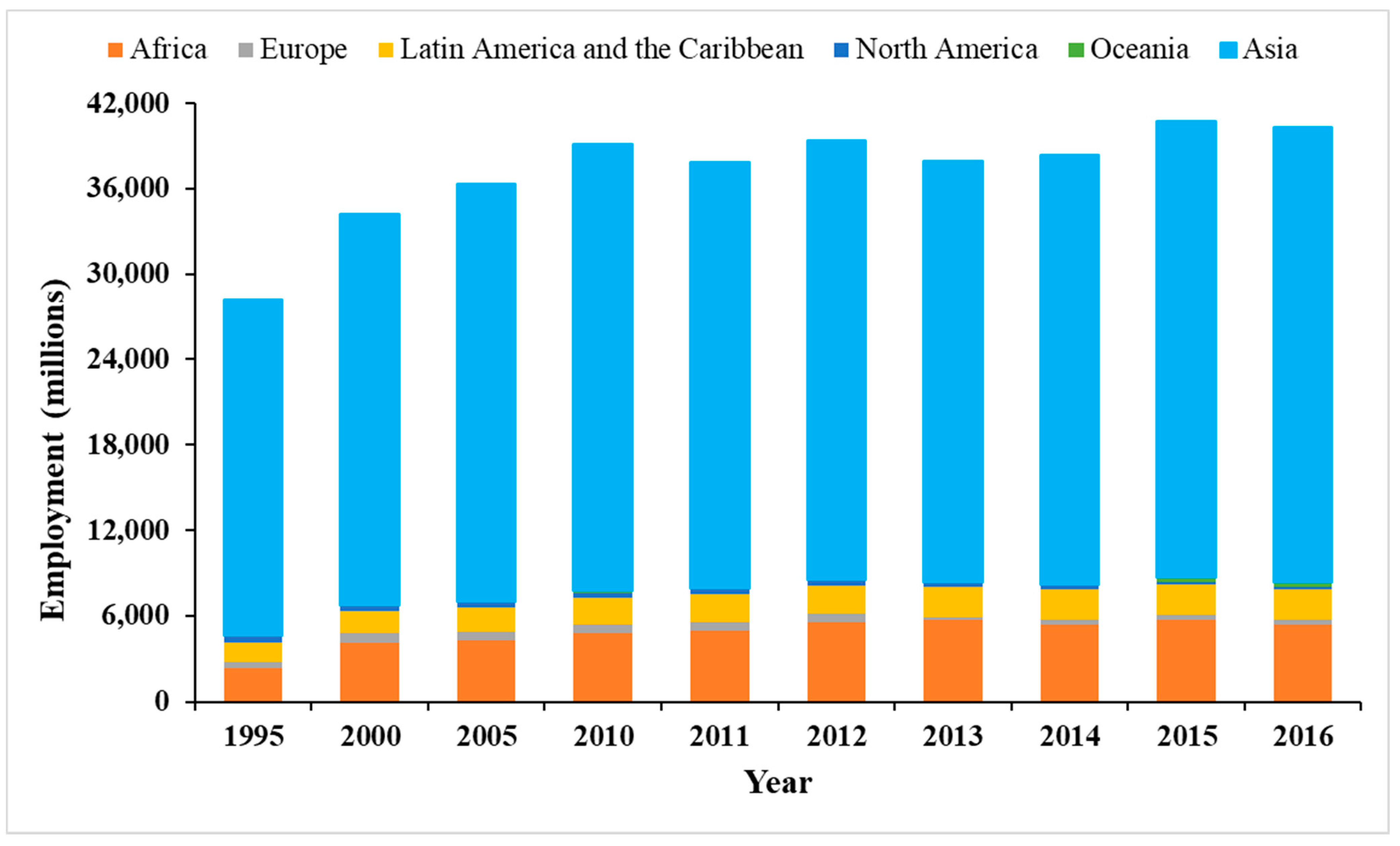
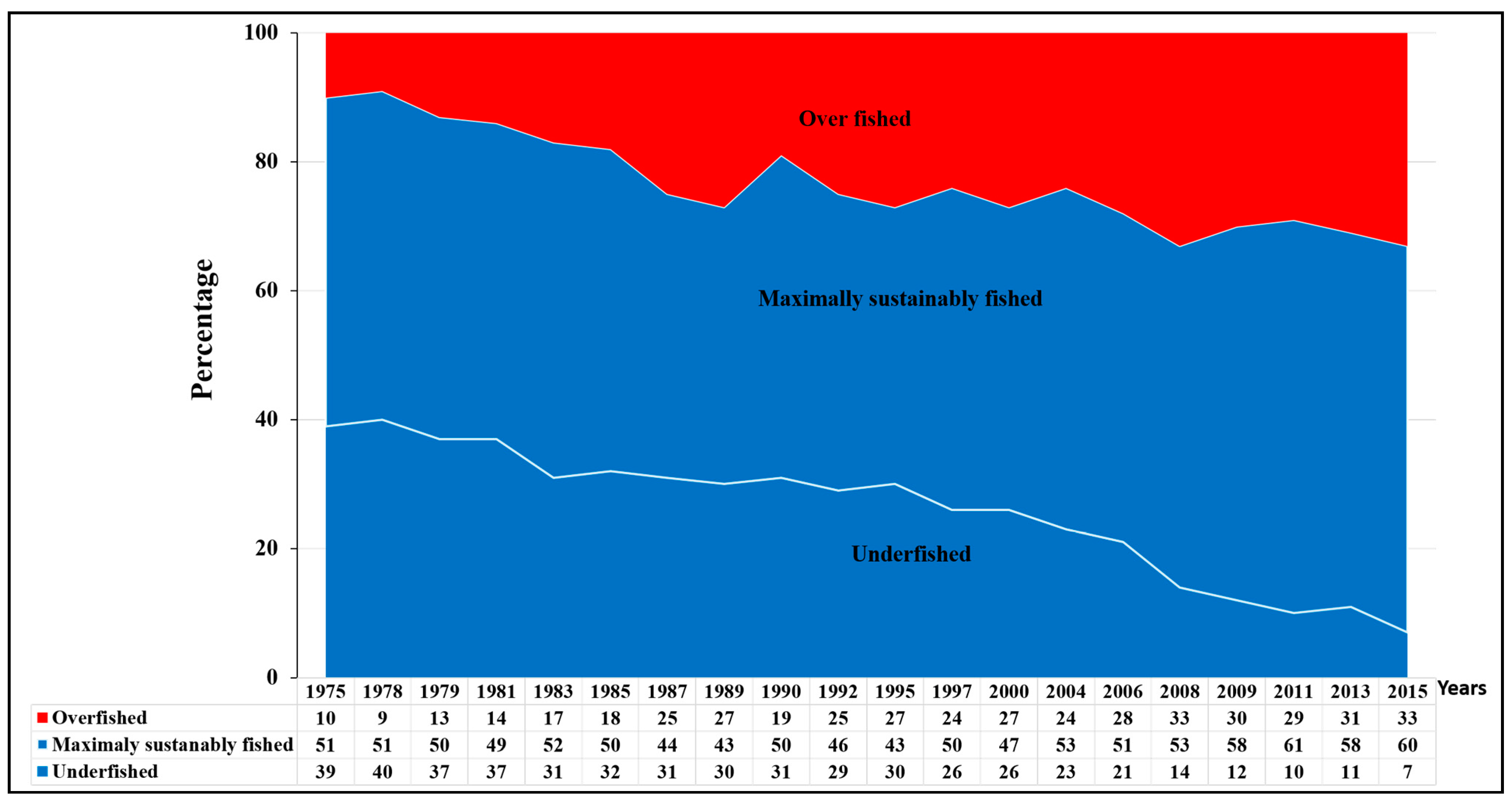
2. Global Fish Stock Status
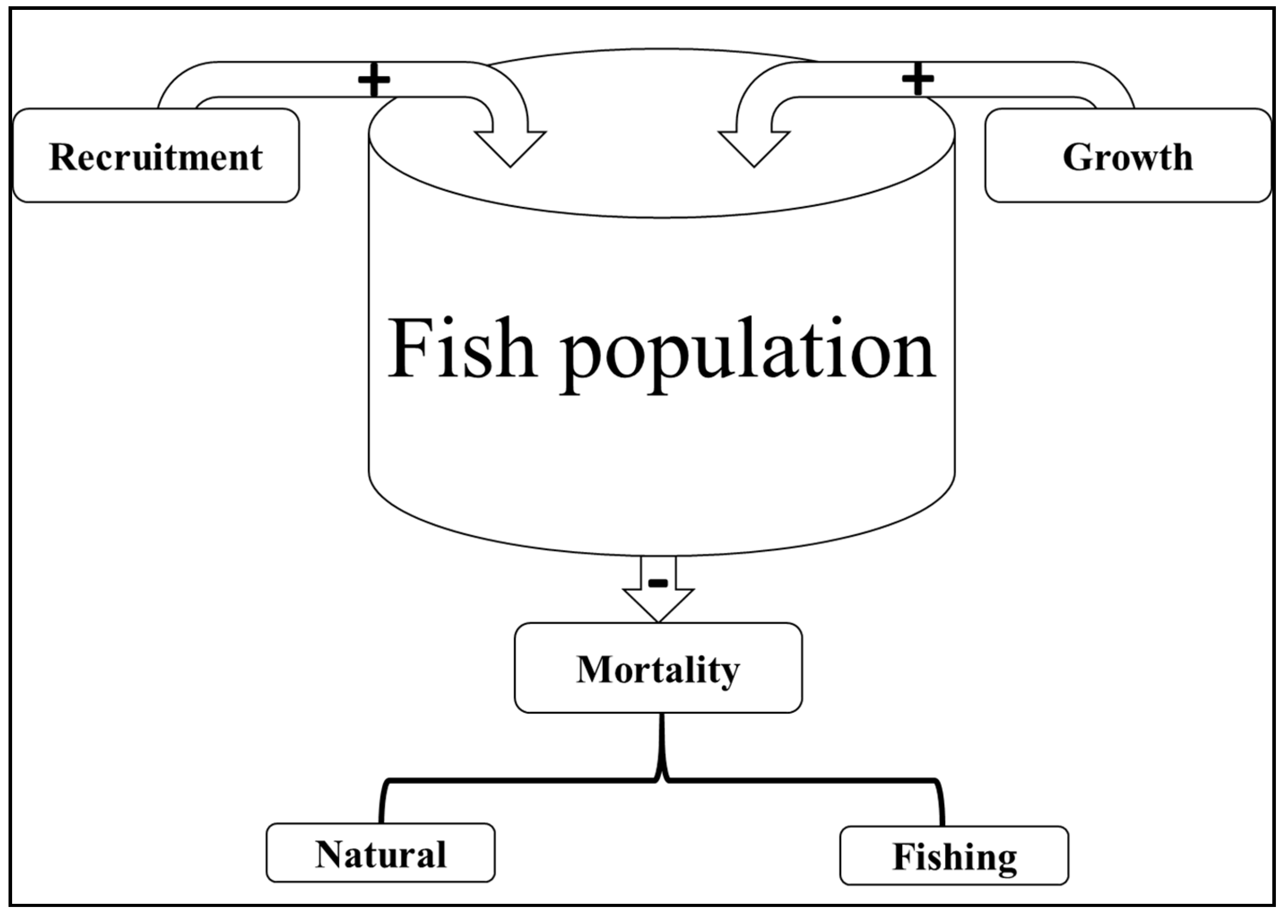
3. Dynamics in Fish Population Size and Life History Traits
3.1. Factors Affecting Fish Population Imbalance
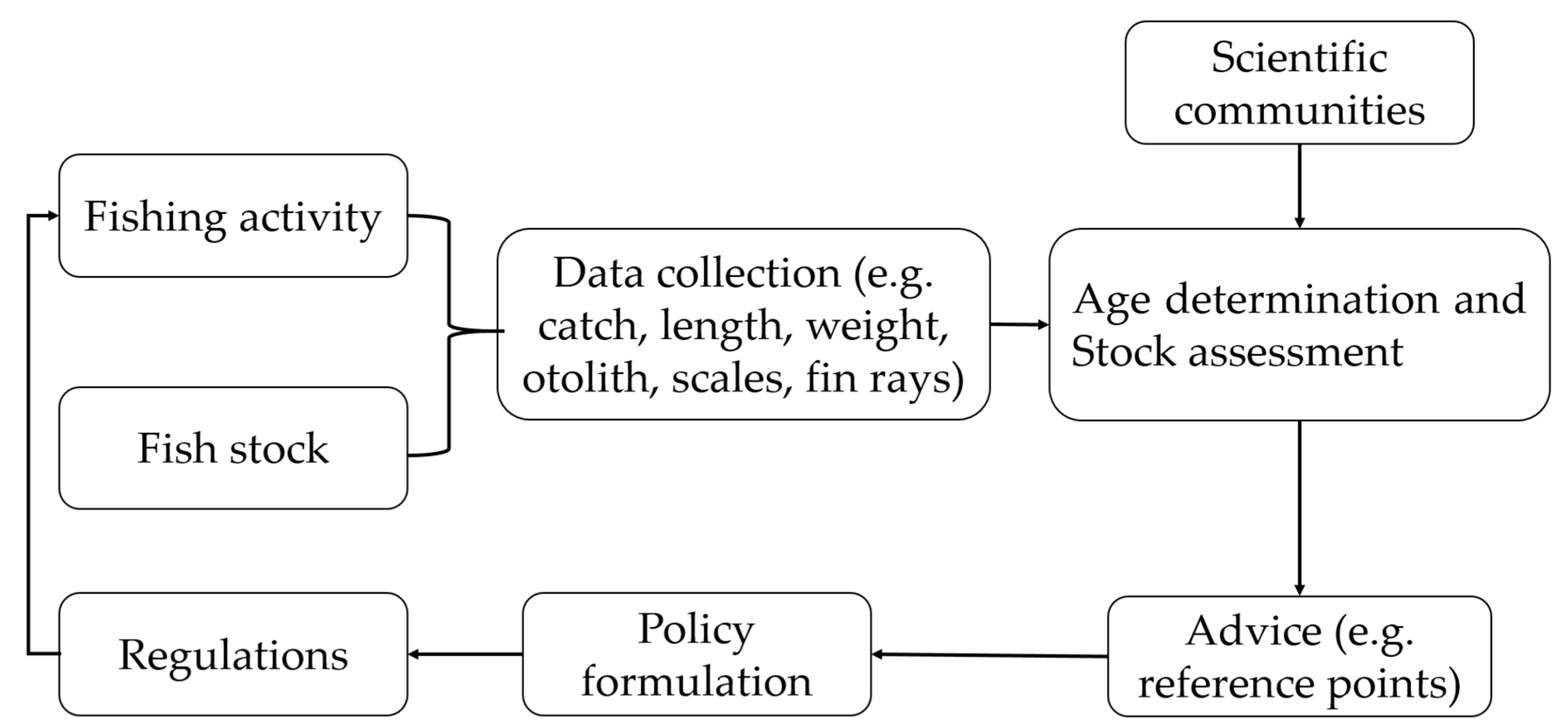
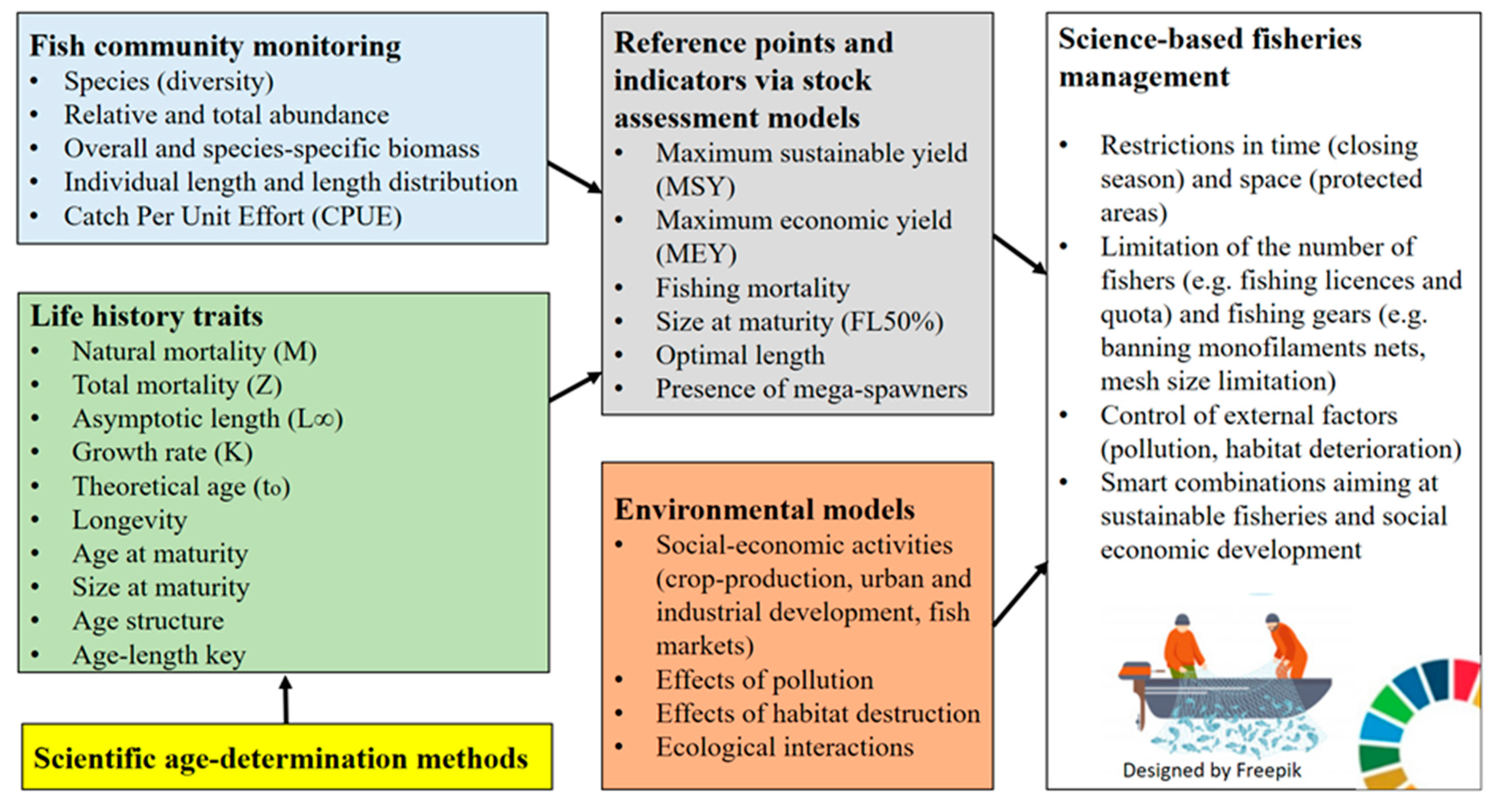
3.2. Life History Traits as a Basis for Stock Assessment and Fisheries Management
3.3. Advantages and Limitations of the Hard Structures Used for Age Estimation
3.3.1. Otoliths
3.3.2. Scales
3.3.3. Fin Rays
4. Precision and Accuracy of Otolith Preparation Methods
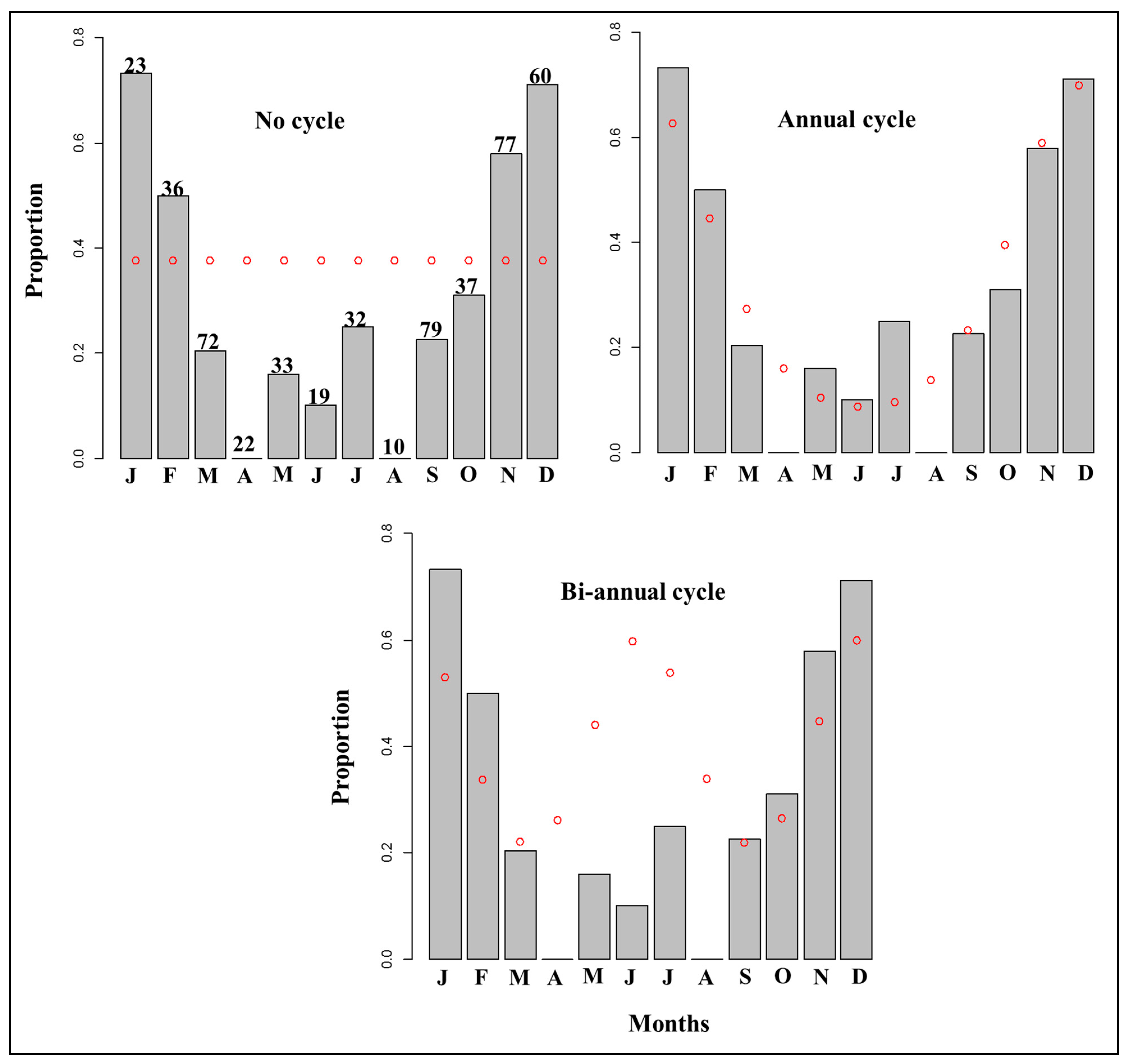
5. Validation of the Rate of Growth Zone Deposition
5.1. Mark-Recapture of Chemically Tagged Fish
5.2. Marginal Increment Analysis
5.3. Edge Analysis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Welcomme, R.L.; Cowx, I.G.; Coates, D.; Béné, C.; Funge-Smith, S.; Halls, A.; Lorenzen, K. Inland capture fisheries. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2881–2896. [Google Scholar] [CrossRef] [PubMed]
- Lynch, A.J.; Cooke, S.J.; Deines, A.M.; Bower, S.D.; Bunnell, D.B.; Cowx, I.G.; Nguyen, V.M.; Nohner, J.; Phouthavong, K.; Riley, B. The social, economic, and environmental importance of inland fish and fisheries. Environ. Rev. 2016, 24, 115–121. [Google Scholar] [CrossRef] [Green Version]
- FAO. The State of World Fisheriess and Aquaculture 2018-Meeting the Sustainable Development Goals, Room, Italy. Licence: CC BY-NC-SA, 3.0 IGO; FAO: Rome, Italy, 2018. [Google Scholar]
- Béné, C.; Macfadyen, G.; Allison, E.H. Increasing the Contribution of Small-Scale Fisheries to Poverty Alleviation and Food Security; 9251056641; FAO Fisheries Technical Paper, No. 481; FAO: Rome, Italy, 2007; p. 125. [Google Scholar]
- FAO and World Fish Center. Small-Scale Capture Fisheries: A Global Overview with Emphasis on Developing Countries; World Bank: Washington, DC, USA, 2008; p. 64. [Google Scholar]
- Coulthard, S.; Johnson, D.; McGregor, J.A. Poverty, sustainability and human wellbeing: A social wellbeing approach to the global fisheries crisis. Glob. Environ. Chang. 2011, 21, 453–463. [Google Scholar] [CrossRef]
- Finegold, C. Fisheries, Sustainability and Development; The Royal Swedish Academy of Agriculture and Forestry: Stockholm, Sweden, 2009; pp. 353–364. [Google Scholar]
- Monfort, M.C. The Role of Women in the Seafood Industry; The GLOBEFISH Research Programme: Rome, Italy, 2015; p. 119. [Google Scholar]
- Lem, A. The WTO Doha Round and Fisheries: What’s at Stake; FAO Fact Sheet for WTO Ministerial Conference in Cancun: Cancun, Mexico, 2003. [Google Scholar]
- Ignatius, S.; Delaney, A.; Haapasaari, P. Socio-cultural values as a dimension of fisheries governance: The cases of Baltic salmon and herring. Environ. Sci. Policy 2019, 94, 1–8. [Google Scholar] [CrossRef]
- Ives, C.D.; Kendal, D. The role of social values in the management of ecological systems. J. Environ. Manag. 2014, 144, 67–72. [Google Scholar] [CrossRef] [PubMed]
- FAO. FAO The State of World Fisheries and Aquaculture 2010, Rome, Italy; 9251066752; Food and Agriculture Organization of the United Nations: Rome, Italy, 2010. [Google Scholar]
- Barrett, C.B. Measuring food insecurity. Science 2010, 327, 825–828. [Google Scholar] [CrossRef] [PubMed]
- Frid, C.L.J.; Paramor, O.A.L. Feeding the qorld: What role for fisheries? ICES J. Mar. Sci. 2012, 69, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Roos, N.; Wahab, M.A.; Chamnan, C.; Thilsted, S.H. The role of fish in food-based strategies to combat vitamin A and mineral deficiencies in developing countries. J. Nutr. 2007, 137, 1106–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youn, S.-J.; Taylor, W.W.; Lynch, A.J.; Cowx, I.G.; Beard Jr, T.D.; Bartley, D.; Wu, F. Inland capture fishery contributions to global food security and threats to their future. Glob. Food Secur. 2014, 3, 142–148. [Google Scholar] [CrossRef]
- Kapetsky, J. Review of the State of World Fishery Resources: Inland Fisheries; No. 942, Rev.1; FAO Fisheries Circular: Rome, Italy, 2003; p. 60. [Google Scholar]
- Bartley, D.; De Graaf, G.; Valbo-Jørgensen, J.; Marmulla, G. Inland capture fisheries: Status and data issues. Fish. Manag. Ecol. 2015, 22, 71–77. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.-I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.-H.; Soto, D.; Stiassny, M.L. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Baron, J.S.; Poff, N.L.; Angermeier, P.L.; Dahm, C.N.; Gleick, P.H.; Hairston Jr, N.G.; Jackson, R.B.; Johnston, C.A.; Richter, B.D.; Steinman, A.D. Meeting ecological and societal needs for freshwater. Ecol. Appl. 2002, 12, 1247–1260. [Google Scholar] [CrossRef]
- Navarro-Ortega, A.; Acuña, V.; Bellin, A.; Burek, P.; Cassiani, G.; Choukr-Allah, R.; Dolédec, S.; Elosegi, A.; Ferrari, F.; Ginebreda, A. Managing the effects of multiple stressors on aquatic ecosystems under water scarcity. The GLOBAQUA project. Sci. Total Environ. 2015, 503, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Søndergaard, M.; Jeppesen, E. Anthropogenic impacts on lake and stream ecosystems, and approaches to restoration. J. Appl. Ecol. 2007, 44, 1089–1094. [Google Scholar] [CrossRef]
- WEF. The Global Risks Report 2017, 12th ed.; 91–93 Route de la Capite CH-1223 Cologny: Geneva, Switzerland, 2017. [Google Scholar]
- de Graaf, M.; van Zwieten, P.A.M.; Machiels, M.A.M.; Lemma, E.; Wudneh, T.; Dejen, E.; Sibbing, F.A. Vulnerability to a small-scale commercial fishery of Lake Tana’s (Ethiopia) endemic Labeobarbus compared with African catfish and Nile tilapia: An example of recruitment-overfishing? Fish. Res. 2006, 82, 304–318. [Google Scholar] [CrossRef]
- Tanaka, S.K.; Zhu, T.; Lund, J.R.; Howitt, R.E.; Jenkins, M.W.; Pulido, M.A.; Tauber, M.; Ritzema, R.S.; Ferreira, I.C. Climate warming and water management adaptation for California. Clim. Chang. 2006, 76, 361–387. [Google Scholar] [CrossRef]
- Vörösmarty, C.J.; McIntyre, P.B.; Gessner, M.O.; Dudgeon, D.; Prusevich, A.; Green, P.; Glidden, S.; Bunn, S.E.; Sullivan, C.A.; Liermann, C.R. Global threats to human water security and river biodiversity. Nature 2010, 467, 555. [Google Scholar] [CrossRef] [PubMed]
- Beard, J.T.D.; Arlinghaus, R.; Cooke, S.J.; McIntyre, P.B.; De Silva, S.; Bartley, D.; Cowx, I.G. Ecosystem approach to inland fisheries: Research needs and implementation strategies. Biol. Lett. 2011, 7, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Davidson, E.A.; de Araújo, A.C.; Artaxo, P.; Balch, J.K.; Brown, I.F.; Bustamante, M.M.; Coe, M.T.; DeFries, R.S.; Keller, M.; Longo, M. The Amazon basin in transition. Nature 2012, 481, 321. [Google Scholar] [CrossRef]
- Ziv, G.; Baran, E.; Nam, S.; Rodríguez-Iturbe, I.; Levin, S.A. Trading-off fish biodiversity, food security, and hydropower in the Mekong River Basin. Proc. Natl. Acad. Sci. USA 2012, 109, 5609–5614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dejen, E.; Anteneh, W.; Vijverberg, J. The decline of the Lake Tana (Ethiopia) fisheries: Causes and possible solutions. Land Degrad. Dev. 2017, 28, 1842–1851. [Google Scholar] [CrossRef] [Green Version]
- Gebremedhin, S.; Getahun, A.; Anteneh, W.; Gedif, B.; Gashu, B.; Tefera, B.; Berhanie, Z.; Alemaw, D. Effect of large weirs on abundance and diversity of migratory Labeobarbus species in tributaries of Lake Tana, Ethiopia. Afr. J. Aquat. Sci. 2017, 42, 367–373. [Google Scholar]
- Gebremedhin, S.; Getahun, A.; Anteneh, W.; Bruneel, S.; Goethals, P. A drivers-pressure-state-impact-responses framework to support the sustainability of fish and fisheries in Lake Tana, Ethiopia. Sustainability 2018, 10, 2957. [Google Scholar] [CrossRef] [Green Version]
- WWF. Living Planet Report 2016. In Risk and Resilience in a New Era; WWF International: Gland, Switzerland, 2016. [Google Scholar]
- Ogutu-Ohwayo, R. The decline of the native fishes of lakes Victoria and Kyoga (East Africa) and the impact of introduced species, especially the Nile perch, Lates niloticus, and the Nile tilapia, Oreochromis niloticus. Environ. Biol. Fishes 1990, 27, 81–96. [Google Scholar] [CrossRef]
- Lowe-McConnell, R. Recent research in the African Great Lakes: Fisheries, biodiversity and cichlid evolution. In Proceedings of the Freshwater Forum, Freshwater Biological Association, Dushanbe, Tajikistan, 29 August–1 September 2003; Volume 20, pp. 4–64. [Google Scholar]
- Ogutu-Ohwayo, R.; Balirwa, J.S. Management challenges of freshwater fisheries in Africa. Lakes Reserv. Res. Manag. 2006, 11, 215–226. [Google Scholar] [CrossRef]
- de Graaf, M.; Machiels, M.A.M.; Wudneh, T.; Sibbing, F.A. Declining stocks of Lake Tana’s endemic Barbus species flock (Pisces, Cyprinidae): Natural variation or human impact? Biol. Conserv. 2004, 116, 277–287. [Google Scholar] [CrossRef]
- Wey, O.L.F. Lake Malawi is Home to Unique Fish Species. Nearly 10% are Endangered. Available online: https://theconversation.com/lake-malawi-is-home-to-unique-fish-species-nearly-10-are-endangered-110294 (accessed on 10 December 2019).
- Turner, G.F.; Tweddle, D.; Makwinja, R.D. Changes in demersal cichlid communities as a result of trawling in southern Lake Malaŵi. In The Impact of Species Changes in African Lakes; Pitcher, T.J., Hart, P.J.B., Eds.; Springer: Dordrecht, The Netherlands, 1995; pp. 397–412. [Google Scholar]
- Turner, G.F. Management, conservation and species changes of exploited fish stocks in Lake Malaŵi. In The Impact of Species Changes in African Lakes; Pitcher, T.J., Hart, P.J.B., Eds.; Springer: Dordrecht, The Netherlands, 1995; pp. 365–395. [Google Scholar]
- Turner, J. Some effects of demersal trawling in Lake Malawi (Lake Nyasa) from 1968 to 1974. J. Fish Biol. 1977, 10, 261–271. [Google Scholar] [CrossRef]
- Turner, J. Changes in the size structure of cichlid populations of Lake Malaŵi resulting from bottom trawling. J. Fish. Board Can. 1977, 34, 232–238. [Google Scholar] [CrossRef]
- Weyl, O.; Nyasulu, T.; Rusuwa, B. Assessment of catch, effort and species changes in the pair-trawl fishery of southern Lake Malawi, Malawi, Africa. Fish. Manag. Ecol. 2005, 12, 395–402. [Google Scholar] [CrossRef]
- FAO. Fisheries Management in the Southeast Arm of Lake Malawi, the Upper Shire River and Lake Malombe, with Particular Reference to the fisheries on Chambo (Oreochromis spp.); CIFA Technical Paper, No. 21.; FAO: Rome, Italy, 1993. [Google Scholar]
- Banda, M.; Jamu, D.; Njaya, F.; Makuliwa, M.; Maluwa, A. In The chambo restoration strategic plan. In Proceedings of the National Workshop, 71, Boadzulu Lakeshore Resort, Mangochi, Malawi, 13–16 May 2003. WorldFish Center. [Google Scholar]
- Wey, O.L.F.; Steve, D.; Mafaniso; Hara, F.N.; Denis, T. Fish and fisheries in Lake Malawi: An assessment of four decades of management interventions. In Proceedings of the African Great Lakes Conference, Entebbe, Uganda, 2–5 May 2017; 5 May 2017. [Google Scholar]
- Weyl, O.L.; Ribbink, A.J.; Tweddle, D. Lake Malawi: Fishes, fisheries, biodiversity, health and habitat. Aquat. Ecosyst. Health Manag. 2010, 13, 241–254. [Google Scholar] [CrossRef]
- Brad, C. Dynamics of fish stocks and commercial fisheries in Lake Victoria, East Africa: Implications for management. In Proceedings of the African Great Lakes Conference, Entebbe, Uganda, 2–5 May 2017. [Google Scholar]
- Sayer, C.; Máiz-Tomé, L.; Darwall, W. Freshwater Biodiversity in the Lake Victoria Basin: Guidance for Species Conservation, Site Protection, Climate Resilience and Sustainable Livelihoods; IUCN, Gland: Cambridge, UK, 2018; p. 226. [Google Scholar]
- Gownaris, N.J.; Pikitch, E.K.; Ojwang, W.O.; Michener, R.; Kaufman, L. Predicting species’ vulnerability in a massively perturbed system: The fishes of Lake Turkana, Kenya. PLoS ONE 2015, 10, e0127027. [Google Scholar] [CrossRef] [Green Version]
- Tweddle, D.; Cowx, I.; Peel, R.; Weyl, O. Challenges in fisheries management in the Zambezi, one of the great rivers of Africa. Fish. Manag. Ecol. 2015, 22, 99–111. [Google Scholar] [CrossRef]
- Vijverberg, J.; Dejen, E.; Getahun, A.; Nagelkerke, L.A.J. The composition of fish communities of nine Ethiopian lakes along a north-south gradient: Threats and possible solutions. Anim. Biol. 2012, 62, 315–335. [Google Scholar] [CrossRef] [Green Version]
- Tesfaye, G.; Wolff, M. The state of inland fisheries in Ethiopia: A synopsis with updated estimates of potential yield. Ecohydrol. Hydrobiol. 2014, 14, 200–219. [Google Scholar] [CrossRef]
- Gebremedhin, S.; Bruneel, S.; Getahun, A.; Anteneh, W.; Goethals, P. The endemic species flock of Labeobarbus spp. in L. Tana (Ethiopia) threatened by extinction: Implications for conservation management. Water 2019, 11, 2560. [Google Scholar] [CrossRef] [Green Version]
- Nagelkerke, L.A.; Mina, M.V.; Wudneh, T.; Sibbing, F.A.; Osse, J.W. In Lake Tana, a unique fish fauna needs protection. Bioscience 1995, 45, 772–775. [Google Scholar]
- Getahun, A. Labeobarbus species. In The IUCN Red List of Threatened Species 2010; International Union for Conservation of Nature and Natural Resources: Gland, Switzerland, 2010. [Google Scholar]
- Pauly, D.; Christensen, V.; Dalsgaard, J.; Froese, R.; Torres, F. Fishing down marine food webs. Science 1998, 279, 860–863. [Google Scholar] [CrossRef] [PubMed]
- Kilduf, P. Guide to Fisheries Science and Stock Assessments; The Atlantic States Marine Fisheries Commission: Arlington, VA, USA, 2009; p. 74. [Google Scholar]
- Hunter, A.; Speirs, D.C.; Heath, M.R. Fishery-induced changes to age and length dependent maturation schedules of three demersal fish species in the Firth of Clyde. Fish. Res. 2015, 170, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Lappalainen, A.; Saks, L.; Šuštar, M.; Heikinheimo, O.; Jürgens, K.; Kokkonen, E.; Kurkilahti, M.; Verliin, A.; Vetemaa, M. Length at maturity as a potential indicator of fishing pressure effects on coastal pikeperch (Sander lucioperca) stocks in the northern Baltic Sea. Fish. Res. 2016, 174, 47–57. [Google Scholar] [CrossRef]
- Nagelkerke, L.A.J.; Sibbing, F.A. The large barbs (Barbus spp. Cyprinidae, Teleostei) of Lake Tana (Ethiopia), with a description of a new species, Barbus osseensis. Neth. J. Zool. 2000, 50, 179–214. [Google Scholar] [CrossRef]
- Sibbing, F.A.; Nagelkerke, L.A.J. Resource partitioning by Lake Tana barbs predicted from fish morphometrics and prey characteristics. Rev. Fish Biol. Fish. 2001, 10, 393–437. [Google Scholar] [CrossRef]
- Ficke, A.D.; Myrick, C.A.; Hansen, L.J. Potential impacts of global climate change on freshwater fisheries. Rev. Fish Biol. Fish. 2007, 17, 581–613. [Google Scholar] [CrossRef]
- Johnson, J.L. Climate Change and Fishery Sustainability in Lake Victoria. Afr. J. Trop. Hydrobiol. Fish. 2009, 12, 31–36. [Google Scholar] [CrossRef]
- Karmakar, S.; Purkait, S.; Das, A.; Samanta, R.; Kumar, K. Climate change and Inland fisheries: Impact and mitigation strategies. J. Exp. Zool. India 2018, 21, 329–335. [Google Scholar]
- Plisnier, P.-D.; Nshombo, M.; Mgana, H.; Ntakimazi, G. Monitoring climate change and anthropogenic pressure at Lake Tanganyika. J. Great Lakes Res. 2018, 44, 1194–1208. [Google Scholar] [CrossRef]
- Solemdal, P. Maternal effects-a link between the past and the future. J. Sea Res. 1997, 37, 213–227. [Google Scholar] [CrossRef]
- Trippel, E.A. Egg size and viability and seasonal offspring production of young Atlantic cod. Trans. Am. Fish. Soc. 1998, 127, 339–359. [Google Scholar] [CrossRef]
- Froese, R. Keep it simple: Three indicators to deal with overfishing. Fish Fish. 2004, 5, 86–91. [Google Scholar] [CrossRef] [Green Version]
- Craig, J. Aging in fish. Can. J. Zool. 1985, 63, 1–8. [Google Scholar] [CrossRef]
- Beverton, R.J. Longevity in fish: Some ecological and evolutionary considerations. In Evolution of Longevity in Animals; Woodhead, A.D., Thompson, K.H., Eds.; Springer: Boston, MA, USA, 1987; pp. 161–185. [Google Scholar]
- Willis, D.W.; Murphy, B.R.; Guy, C.S. Stock density indices: Development, use, and limitations. Rev. Fish. Sci. 1993, 1, 203–222. [Google Scholar] [CrossRef]
- Neumann, R.; Allen, M. Size structure. In Analysis and Interpretation of Freshwater Fisheries Data; Guy, C.S., Brown, M.L., Eds.; American Fisheries Society: Bethesda, Maryland, 2007; pp. 375–421. [Google Scholar]
- Neumann, R.M.; Guy, C.S.; Willis, D.W. Length, weight, and associated indices. In Fisheries Techniques, 3rd ed.; Zale, A.V., Parrish, D.L., Sutton, T.M., Eds.; American Fisheries Society: Bethesda, Maryland, 2012; pp. 637–676. [Google Scholar]
- Isermann, D.A.; Paukert, C.P. Regulating harvest. In Inland Fisheries Management in North America, 3rd ed.; Wayne, A.H., Michael, C.Q., Eds.; American Fisheries Society: Bethesda, Maryland, 2010; pp. 185–212. [Google Scholar]
- Pope, K.L.; Lochmann, S.E.; Young, M.K. Methods for assessing fish populations. In Inland Fisheries Management in North America, 3rd ed.; Hubert, W.A., Quist, M.C., Eds.; American Fisheries Society: Bethesda, MD, USA, 2010; pp. 325–351. [Google Scholar]
- Ricker, W.E. Computation and Interpretation of Biological Statistics of Fish Populations; Bulletin of the Fisheries Research Board of Canada: Ottawa, ON, Canada, 1975; p. 382. [Google Scholar]
- Campana, S. Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J. Fish Biol. 2001, 59, 197–242. [Google Scholar] [CrossRef]
- Moltschaniwskyj, N.; Cappo, M. Alternatives to sectioned otoliths: The use of other structures and chemical techniques to estimate age and growth for marine vertebrates and invertebrates. In Tropical Fish Otoliths: Information for Assessment, Management and Ecology; Green, B.S., Mapstone, B.D., Carlos, G., Begg, G.A., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 133–173. [Google Scholar]
- Green, B.; Mapstone, B.; Carlos, G.; Begg, G. Introduction to otoliths and fisheries in the tropics. In Tropical Fish Otoliths: Information for Assessment, Management and Ecology, Reviews: Methods and Technologies in Fish Biology and Fisheries, 11th ed.; Green, B., Mapstone, B., Carlos, G., Begg, G., Eds.; Springer: Dordrecht, The Netherlands, 2009. [Google Scholar]
- Göçer, M.; Ekingen, G. Comparisons of various bony structures for the age determination of Liza ramada (Risso, 1826) population from the Mersin Bay. J. Fish. Aquat. Sci. 2005, 22, 211–213. [Google Scholar]
- Begg, G.A.; Campana, S.E.; Fowler, A.J.; Suthers, I.M. Otolith research and application: Current directions in innovation and implementation. Mar. Freshw. Res. 2005, 56, 477–483. [Google Scholar] [CrossRef] [Green Version]
- Chilton, D.E.; Beamish, R.J. Age determination methods for fishes studied by the groundfish program at the Pacific Biological Station. Can. Spec. Publ. Fish. Aquat. Sci. 1982, 60, 102. [Google Scholar]
- Admassu, D. Age and Growth Determination of Tilapia, Oreochromis Niloticus L. (Pisces: Cichlidae) in Some Lakes in Ethiopia. Ph.D. Thesis, Addis Ababa University, Addis Ababa, Ethiopia, 1998; p. 115. [Google Scholar]
- Admassu, D.; Casselman, J.M. Otolith age determination for adult tilapia, Oreochromis niloticus L. from Lake Awassa (Ethiopian Rift Valley) by interpreting biannuli and differentiating biannual recruitment. Hydrobiologia 2000, 418, 15–24. [Google Scholar] [CrossRef]
- Brown, P.; Green, C.; Sivakumaran, K.; Stoessel, D.; Giles, A. Validating otolith annuli for annual age determination of common carp. Trans. Am. Fish. Soc. 2004, 133, 190–196. [Google Scholar] [CrossRef]
- Phelps, Q.E.; Edwards, K.R.; Willis, D.W. Precision of five structures for estimating age of common carp. N. Am. J. Fish. Manag. 2007, 27, 103–105. [Google Scholar] [CrossRef]
- Weyl, O.L.; Stadtlander, T.; Booth, A.J. Establishment of translocated populations of smallmouth yellowfish, Labeobarbus aeneus (Pisces: Cyprinidae), in lentic and lotic habitats in the Great Fish River system, South Africa. Afr. Zool. 2009, 44, 93–105. [Google Scholar] [CrossRef]
- Butler, J.L.; Folkvord, A. Section 2: Estimation of fish age and growth (Editorial). Fish. Res. 2000, 46, 51–52. [Google Scholar] [CrossRef]
- Casselman, J.M. Growth and relative size of calcified structures of fish. Trans. Am. Fish. Soc. 1990, 119, 673–688. [Google Scholar] [CrossRef]
- Cardinale, M.; Arrhenius, F. Using otolith weight to estimate the age of haddock (Melanogrammus aeglefinus): A tree model application. J. Appl. Ichthyol. 2004, 20, 470–475. [Google Scholar] [CrossRef]
- Worthington, D.; Doherty, P.; Fowler, A. Variation in the relationship between otolith weight and age: Implications for the estimation of age of two tropical damselfish (Pomacentrus moluccensis and P. wardi). Can. J. Fish. Aquat. Sci. 1995, 52, 233–242. [Google Scholar] [CrossRef]
- Pilling, G.; Grandcourt, E.; Kirkwood, G. The utility of otolith weight as a predictor of age in the emperor Lethrinus mahsena and other tropical fish species. Fish. Res. 2003, 60, 493–506. [Google Scholar] [CrossRef]
- Quist, M.C.; Pegg, M.A.; DeVries, D.R. Age and growth. In Fisheries Techniques, 3rd ed.; Zale, A.V., Parrish, D.L., Sutton, T.M., Eds.; American Fisheries Society: Bethesda, Maryland, 2012; pp. 677–731. [Google Scholar]
- Braaten, P.J.; Doeringsfeld, M.R.; Guy, C.S. Comparison of age and growth estimates for river carpsuckers using scales and dorsal fin ray sections. North Am. J. Fish. Manag. 1999, 19, 786–792. [Google Scholar] [CrossRef]
- Maceina, M.; Sammons, S. An evaluation of different structures to age freshwater fish from a northeastern US river. Fish. Manag. Ecol. 2006, 13, 237–242. [Google Scholar] [CrossRef]
- Kanwal, B.; Pathani, S. Age-growth, length-weight and condition factor of a hill stream fish, Gara lamta (Hamilton Buchanan) of Kumaun Himalaya. Nat. Sci. 2011, 9, 199–206. [Google Scholar]
- Khan, S.; Khan, M.A.; Miyan, K. Comparison of age estimates from otoliths, vertebrae, and pectoral spines in African sharptooth catfish, Clarias gariepinus (Burchell). Est. J. Ecol. 2011, 60, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Rick, W.; Deanna, L.; Kimberly, H.; Susan, M.; Theresa, J.C.; Ross, F.T. Comparison of Scales, Pectoral Fin Rays, and Otoliths for Estimating Age, Growth, and Mortality of Lake Whitefish, Coregonus Clupeaformis, in Great Slave Lake; Canadian Science Advisory Secretariat: Ottawa, ON, Canada, 2017. [Google Scholar]
- Beamish, R.J. Use of fin-ray sections to age walleye pollock, Pacific cod, and albacore, and the importance of this method. Trans. Am. Fish. Soc. 1981, 110, 287–299. [Google Scholar] [CrossRef]
- Cass, A.J.; Beamish, R.J. First evidence of validity of the fin-ray method of age determination for marine fishes. North Am. J. Fish. Manag. 1983, 3, 182–188. [Google Scholar] [CrossRef]
- Chilton, D.; Bilton, H. New method for ageing Chinook Salmon (Onchorhynchus tshawytscha) using dorsal fin rays, and evidence if its validity. Can. J. Fish. Aquat. Sci. 1986, 43, 1588–1594. [Google Scholar] [CrossRef]
- Michaletz, P.H.; Nicks, D.M.; Buckner Jr, E.W. Accuracy and precision of estimates of back-calculated channel catfish lengths and growth increments using pectoral spines and otoliths. North Am. J. Fish. Manag. 2009, 29, 1664–1675. [Google Scholar] [CrossRef]
- Sikstrom, C. Otolith, pectoral fin ray, and scale age determinations for Arctic grayling. Progress. Fish Cult. 1983, 45, 220–223. [Google Scholar] [CrossRef]
- Williamson, C.; Macdonald, J.S. The Use of Three Ageing Techniques to Estimate the Growth Rates for Rainbow Trout, Oncorhynchus Mykiss, and Bull Trout, Salvelinus Confluentus, from Selected Locations Near Takla Lake, British Columbia; Canadian Technical Report of Fisheries and Aquatic Sciences; Fisheries & Oceans Canada, Pacific Region, Science Branch: West Vancouver, BC, Canada, 1997; p. 20. [Google Scholar]
- Mills, K.H.; Chalanchuk, S.M. The Fin-Ray Method of Aging Lake Whitefish. Ann. Zool. Fennici 2004, 215–223. [Google Scholar]
- Zymonas, N.; McMahon, T. Comparison of pelvic fin rays, scales and otoliths for estimating age and growth of bull trout, Salvelinus confluentus. Fish. Manag. Ecol. 2009, 16, 155–164. [Google Scholar] [CrossRef]
- Beamish, R.; Harvey, H. Age determination in the white sucker. J. Fish. Board Can. 1969, 26, 633–638. [Google Scholar] [CrossRef]
- Mills, K.; Beamish, R. Comparison of fin-ray and scale age determinations for lake whitefish (Coregonus clupeaformis) and their implications for estimates of growth and annual survival. Can. J. Fish. Aquat. Sci. 1980, 37, 534–544. [Google Scholar] [CrossRef]
- Maraldo, D.C.; MacCrimmon, H.R. Comparison of ageing methods and growth rates for largemouth bass, Micropterus salmoides Lacepede, from northern latitudes. Environ. Biol. Fishes 1979, 4, 263–271. [Google Scholar] [CrossRef]
- Barber, W.E.; McFarlane, G.A. Evaluation of three techniques to age Arctic char from Alaskan and Canadian waters. Trans. Am. Fish. Soc. 1987, 116, 874–881. [Google Scholar] [CrossRef]
- Baxter, H.W.A.; Harrington, G.T.M. Comparison of age determinations based on scales, otoliths and fin rays for cutthroat trout from Yellowstone Lake. Northwest Sci. 1987, 61, 32–36. [Google Scholar]
- Steward, C.A.; DeMaria, K.D.; Shenker, J.M. Using otolith morphometrics to quickly and inexpensively predict age in the gray angelfish (Pomacanthus arcuatus). Fish. Res. 2009, 99, 123–129. [Google Scholar] [CrossRef]
- Campana, S.E.; Annand, M.C.; McMillan, J.I. Graphical and statistical methods for determining the consistency of age determinations. Trans. Am. Fish. Soc. 1995, 124, 131–138. [Google Scholar] [CrossRef]
- Marriott, R.J.; Mapstone, B.D. Consequences of inappropriate criteria for accepting age estimates from otoliths, with a case study for a long-lived tropical reef fish. Can. J. Fish. Aquat. Sci. 2006, 63, 2259–2274. [Google Scholar] [CrossRef]
- Fowler, A. Age in years from otoliths of adult tropical fish. In Tropical Fish Otoliths: Information for Assessment, Management and Ecology; Springer: Dordrecht, The Netherland, 2009; pp. 55–92. [Google Scholar]
- Campana, S.E. Comparison of age determination methods for the starry flounder. Trans. Am. Fish. Soc. 1984, 113, 365–369. [Google Scholar] [CrossRef]
- Marriott, R.J.; Mapstone, B.D.; Ballagh, A.C.; Currey, L.M.; Penny, A.; Williams, A.J.; Jackson, G.; Lou, D.; Mapleston, A.J.; Jarvis, N.D. Accepting final counts from repeat readings of otoliths: Should a common criterion apply to the age estimation of fish? Mar. Freshw. Res. 2010, 61, 1171–1184. [Google Scholar] [CrossRef] [Green Version]
- Sylvester, R.M.; Berry, C.R., Jr. Comparison of white sucker age estimates from scales, pectoral fin rays, and otoliths. N. Am. J. Fish. Manag. 2006, 26, 24–31. [Google Scholar] [CrossRef]
- Beamish, R.; Fournier, D. A method for comparing the precision of a set of age determinations. Can. J. Fish. Aquat. Sci. 1981, 38, 982–983. [Google Scholar] [CrossRef]
- Chang, W.Y. A statistical method for evaluating the reproducibility of age determination. Can. J. Fish. Aquat. Sci. 1982, 39, 1208–1210. [Google Scholar] [CrossRef]
- Hoenig, J.; Morgan, M.; Brown, C. Analysing differences between two age determination methods by tests of symmetry. Can. J. Fish. Aquat. Sci. 1995, 52, 364–368. [Google Scholar] [CrossRef]
- Ogle, D.H. Modified AgeBias Plot; FishR: Pittsburgh, WI, USA; Available online: http://derekogle.com/fishR/2017-04-14-ModifiedAgeBiasPlot (accessed on 3 April 2019).
- Campana, S.E. Chemistry and composition of fish otoliths: Pathways, mechanisms and applications. Mar. Ecol. Prog. Ser. 1999, 188, 263–297. [Google Scholar] [CrossRef] [Green Version]
- Secor, D.H.; Dean, J.M. Comparison of otolith-based back-calculation methods to determine individual growth histories of larval striped bass, Morone saxatilis. Can. J. Fish. Aquat. Sci. 1992, 49, 1439–1454. [Google Scholar] [CrossRef]
- Secor, D.; Houde, E.; Monteleone, D. A mark-release experiment on larval striped bass Morone saxatilis in a Chesapeake Bay tributary. ICES J. Mar. Sci. 1995, 52, 87–101. [Google Scholar] [CrossRef]
- Campana, S.E. Photographic Atlas of Fish Otoliths of the Northwest Atlantic Ocean Canadian Special Publication of Fisheries and Aquatic Sciences No. 133; NRC Research Press: Ottawa, ON, USA, 2004. [Google Scholar]
- Panfili, J.; de Pontual, H.; Troadec, H.; Casselman, J.; Moksness, E.; Wright, P. Manual of Fish Sclerochronology; IRD: Brest, France, 2002; p. 464. [Google Scholar]
- Andrews, A.; Ashford, J.; Brooks, C.; Krusic-Golub, K.; Duhamel, G.; Belchier, M.; Lundstrom, C.; Cailliet, G. Lead–radium dating provides a framework for coordinating the age estimation of Patagonian toothfish (Dissostichus eleginoides) between fishing areas. Mar. Freshw. Res. 2011, 62, 781–789. [Google Scholar] [CrossRef]
- Andrews, A.H.; Coale, K.H.; Nowicki, J.L.; Lundstrom, C.; Palacz, Z.; Burton, E.J.; Cailliet, G.M. Application of an ion-exchange separation technique and thermal ionization mass spectrometry to 226Ra determination in otoliths for radiometric age determination of long-lived fishes. Can. J. Fish. Aquat. Sci. 1999, 56, 1329–1338. [Google Scholar] [CrossRef]
- Andrews, A.H.; Kalish, J.M.; Newman, S.J.; Johnston, J.M. Bomb radiocarbon dating of three important reef-fish species using Indo-Pacific Δ14C chronologies. Mar. Freshw. Res. 2011, 62, 1259–1269. [Google Scholar] [CrossRef]
- Andrews, A.H.; Kerr, L.A.; Cailliet, G.M.; Brown, T.A.; Lundstrom, C.C.; Stanley, R.D. Age validation of canary rockfish (Sebastes pinniger) using two independent otolith techniques: Lead-radium and bomb radiocarbon dating. Mar. Freshw. Res. 2007, 58, 531–541. [Google Scholar] [CrossRef] [Green Version]
- Burton, M.L.; Potts, J.C.; Page, J.; Poholek, A. Age, growth, mortality and reproductive seasonality of jolthead porgy, Calamus bajonado, from Florida waters. PeerJ 2017, 5, e3774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cailliet, G.; Andrews, A.; Burton, E.; Watters, D.; Kline, D.; Ferry-Graham, L. Age determination and validation studies of marine fishes: Do deep-dwellers live longer? Exp. Gerontol. 2001, 36, 739–764. [Google Scholar] [CrossRef]
- Cailliet, G.M.; Yudin, K.G.; Tanaka, S.; Taniuchi, T. Growth characteristics of two populations of Mustelus manazo from Japan based upon cross-readings of vertebral bands. Noaa Tech. Rep. Nmfs 1990, 90, 167–176. [Google Scholar]
- Admassu, D. A Study on the Age and Growth of Adult Oreochromis Niloticus (Pisces: Cichlidae) in Lake Awassa, Ethiopia. Master’s Thesis, Addis Ababa University, Addis Ababa, Ethiopia, 1989. [Google Scholar]
- Alemu, K. Age and Growth of Oreochromis Niloticus (Pisces: Cichlidae) in Lake Hayq, Ethiopia. Master’s Thesis, Addis Ababa University, Addis Ababa, Ethiopia, 1995. [Google Scholar]
- Gebremedhin, S.; Bekaert, K.; Getahun, A.; Bruneel, S.; Anteneh, W.; Goethals, P.; Torreele, E. Comparison of otolith readability and reproducibility of counts of translucent zones using different otolith preparation methods for four endemic Labeobarbus species in Lake Tana, Ethiopia. Water 2019, 11, 1336. [Google Scholar] [CrossRef] [Green Version]
- Gebremedhin, S.; Bekaert, K.; Getahun, A.; Bruneel, S.; Anteneh, W.; Torreele, E.; Goethals, P. Validation of the periodicity of growth zone deposition in otolith of two large endemic Labeobarbus species in Lake Tana, Ethiopia. Afr. Zool. 2019, 54, 231–238. [Google Scholar] [CrossRef]
- Taylor, G.C.; Peel, R.A.; Hay, C.J.; Weyl, O.L. Validation of the periodicity of growth zone formation in the otoliths of four fish species from the Upper Zambezi ecoregion, southern Africa. Afr. Zool. 2016, 51, 153–159. [Google Scholar] [CrossRef]
- Tekle-Giorgis, Y. Comparative Age and Growth Assessment of the African Catfish, Clarias gariepinus Burchell (Clariidae) and, Nile Perch, Latesniloticus, Linn (Centropomidae) in the Three Southern Rift Valley Lakes of Ethiopia (Lakes Awassa, Abaya and Chamo). Ph. D. Thesis, Addis Ababa University, Addis Ababa, Ethiopia, 2002; p. 160. [Google Scholar]
- Winker, H.; Ellender, B.R.; Weyl, O.L.; Booth, A.J. Validation of growth zone deposition in otoliths of two large endemic cyprinids in Lake Gariep, South Africa. Afr. Zool. 2010, 45, 133–138. [Google Scholar] [CrossRef]
- Winker, H.; Weyl, O.L.; Booth, A.J.; Ellender, B.R. Validating and corroborating the deposition of two annual growth zones in asteriscus otoliths of common carp Cyprinus carpio from South Africa’s largest impoundment. J. Fish Biol. 2010, 77, 2210–2228. [Google Scholar] [CrossRef] [PubMed]
- Winker, H.; Weyl, O.L.; Booth, A.J.; Ellender, B.R. Life history strategy and population characteristics of an unexploited riverine cyprinid, Labeo capensis, in the largest impoundment in the Orange River Basin. Afr. Zool. 2012, 47, 85–99. [Google Scholar] [CrossRef]
- Yosef, T.; Casselman, J. A procedure for increasing the precision of otolith age determination of tropical fish by differentiating biannual recruitment. Recent Dev. Fish Otolith Res. 1995, 19, 247. [Google Scholar]
- Piddocke, T.P.; Butler, G.L.; Butcher, P.A.; Purcell, S.W.; Bucher, D.J.; Christidis, L. Age validation in the Lutjanidae: A review. Fish. Res. 2015, 167, 48–63. [Google Scholar] [CrossRef]
- Geffen, A. Validation of otolith increment deposition rate. In Otolith Microstructure Examination and Analysis; D.K, S., Campana, S.E., Eds.; Cananadian Special Publication of Fisheries and Aquatic Sciences: Ottawa, ON, Canada, 1992; Volume 117, pp. 101–113. [Google Scholar]
- Foreman, T. A method of simultaneously tagging large oceanic fish and injecting them with tetracycline. Fish. Bull. 1987, 85, 645–647. [Google Scholar]
- Oliveira, K. Field validation of annular growth rings in the American eel, Anguilla rostrate, using tetracycline-marked otoliths. Fish. Bull. 1996, 94, 186–189. [Google Scholar]
- Beamish, R.J.; Chilton, D.E. Preliminary evaluation of a method to determine the age of sablefish (Anoplopoma fimbria). Can. J. Fish. Aquat. Sci. 1982, 39, 277–287. [Google Scholar] [CrossRef]
- Brown, C.A.; Gruber, S.H. Age assessment of the lemon shark, Negaprion brevirostris, using tetracycline validated vertebral centra. Copeia 1988, 747–753. [Google Scholar] [CrossRef]
- Fowler, A.J. Validation of annual growth increments in the otoliths of a small, tropical coral reef fish. Mar. Ecol. Prog. Ser. Oldendorf 1990, 64, 25–38. [Google Scholar] [CrossRef]
- Ellender, B.R.; Weyl, O.L.; Winker, H. Age and growth and maturity of southern Africa’s largest cyprinid fish, the largemouth yellowfish Labeobarbus kimberleyensis. J. Fish Biol. 2012, 81, 1271–1284. [Google Scholar] [CrossRef] [PubMed]
- Beamish, R.; McFarlane, G. Reevaluation of the interpretation of annuli from otoliths of a long-lived fish, Anoplopoma fimbria. Fish. Res. 2000, 46, 105–111. [Google Scholar] [CrossRef]
- Fowler, A.; Short, D. Validation of age determination from otoliths of the King George whiting Sillaginodes punctata (Perciformes). Mar. Biol. 1998, 130, 577–587. [Google Scholar] [CrossRef]
- Carlson, J.K.; Cortés, E.; Johnson, A.G. Age and growth of the blacknose shark, Carcharhinus acronotus, in the eastern Gulf of Mexico. Copeia 1999, 684–691. [Google Scholar] [CrossRef]
- Goldman, K.J. Age and growth of elasmobranch fishes. Fao Fish. Tech. Pap. 2005, 474, 76. [Google Scholar]
- Cailliet, G.M.; Smith, W.D.; Mollet, H.F.; Goldman, K.J. Age and growth studies of chondrichthyan fishes: The need for consistency in terminology, verification, validation, and growth function fitting. Environ. Biol. Fishes 2006, 77, 211–228. [Google Scholar] [CrossRef]
- Lessa, R.; Santana, F.M.; Duarte-Neto, P. A critical appraisal of marginal increment analysis for assessing temporal periodicity in band formation among tropical sharks. Environ. Biol. Fishes 2006, 77, 309–315. [Google Scholar] [CrossRef]
- Okamura, H.; Punt, A.E.; Semba, Y.; Ichinokawa, M. Marginal increment analysis: A new statistical approach of testing for temporal periodicity in fish age verification. J. Fish Biol. 2013, 82, 1239–1249. [Google Scholar] [CrossRef]
- Lowerre-Barbieri, S.K.; Chittenden Jr, M.E.; Jones, C.M. A comparison of a validated otolith method to age weakfish, Cynoscion regalis, with the traditional scale method. Fish. Bull. 1994, 92, 555–568. [Google Scholar]
- Okamura, H.; Semba, Y. A novel statistical method for validating the periodicity of vertebral growth band formation in elasmobranch fishes. Can. J. Fish. Aquat. Sci. 2009, 66, 771–780. [Google Scholar] [CrossRef]
- Beverton, R.J.; Holt, S.J. On the Dynamics of Exploited Fish Populations; Fisheries Investigation Series 2; UK Ministry of Agriculture: London, UK, 1957; Volume 19, p. 540.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Life History Traits | Assessment and Management Use |
|---|---|
| Age data (i.e., individual age) | Used to determine the age at first maturity and to estimate growth parameters |
| Age and length at first maturity | Used to establish a minimum size limit to ensure that individual fish can reproduce at least once before being harvested |
| Maximum age (i.e., longevity) | Used to estimate the natural mortality rate and set the maximum age in the stock assessment models |
| Asymptotic length (L∞) | Used to set size limits (i.e., restrict mesh sizes and prohibit fishing gears) |
| Growth rate (K) | Indicates how fast the fish will reach the fisher’s minimum size limit |
| Mortality rates (i.e., natural and fishing) and spawning behaviour | Highlights the area and time of increased vulnerability to harvest and can be used for seasonal or area closures to protect the spawning biomass |
| Sex ratio | Used to understand the relationship between individuals, the reproductive potential of the species and the state of the population |
| Habitat preference | Helps to identify areas for protection |
| Methods | Advantages | Disadvantages | Precision | Sample Size | Cost |
|---|---|---|---|---|---|
| Mark-recapture | Validates the frequency of post-tagging growth increments. | Low or non-existent of recaptured fish at liberty (>1 year). Difficulty to identify a post-mark annulus | ±1 year | >1 | Minimal excluding cost of tagging cruise |
| Marginal increment analysis and Edge analysis | Validates frequency of growth increments formation. | Applicable to fast-growing or young fish. A year-round sample is required. | ±1 year | >100 | Minimal other than a fish collection |
| Radiometric | Validates absolute age. Applicable to any recently collected data. Suitable to long-lived fish | Can only distinguish between widely divergent age estimates | ±25–50% | 10–50 | ~$1000 per age category |
| Radiocarbon | Validate both absolute age and frequency of growth increments. Suitable for long lived fish. Does not require recently collected sample. | Some of the fish in the sample must be hatched before 1965. | ±1–3 years | 20–30 | $700–$1000 per otolith |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gebremedhin, S.; Bruneel, S.; Getahun, A.; Anteneh, W.; Goethals, P. Scientific Methods to Understand Fish Population Dynamics and Support Sustainable Fisheries Management. Water 2021, 13, 574. https://doi.org/10.3390/w13040574
Gebremedhin S, Bruneel S, Getahun A, Anteneh W, Goethals P. Scientific Methods to Understand Fish Population Dynamics and Support Sustainable Fisheries Management. Water. 2021; 13(4):574. https://doi.org/10.3390/w13040574
Chicago/Turabian StyleGebremedhin, Shewit, Stijn Bruneel, Abebe Getahun, Wassie Anteneh, and Peter Goethals. 2021. "Scientific Methods to Understand Fish Population Dynamics and Support Sustainable Fisheries Management" Water 13, no. 4: 574. https://doi.org/10.3390/w13040574
APA StyleGebremedhin, S., Bruneel, S., Getahun, A., Anteneh, W., & Goethals, P. (2021). Scientific Methods to Understand Fish Population Dynamics and Support Sustainable Fisheries Management. Water, 13(4), 574. https://doi.org/10.3390/w13040574