
A Review of 50 Years of Study of Hydrology, Wetland Dynamics, Aquatic Metabolism, Water Quality and Trophic Status, and Nutrient Biogeochemistry in the Barataria Basin, Mississippi Delta—System Functioning, Human Impacts and Restoration Approaches


 ,
,  ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
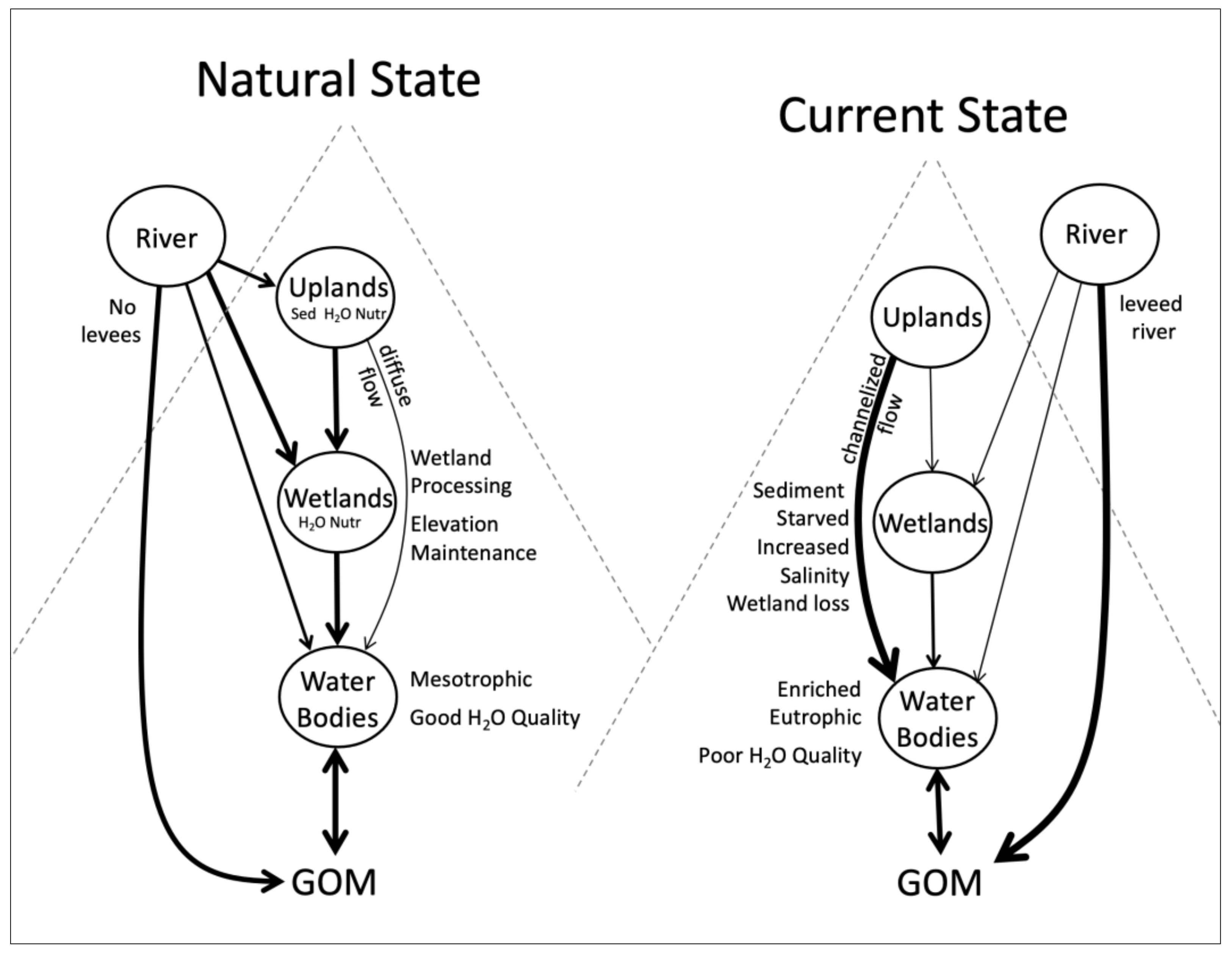
2. Overview Conceptual Model of the Barataria Basin
- Human impacts in the basin include high rates of relative sea-level rise, loss of riverine input, and pervasive alteration of hydrology and nutrient enrichment.
- The hydrology of the basin has been pervasively altered with elimination of riverine surface input, clearing of natural levees and excavation of a dense network of drainage canals in agricultural fields, and extensive channelization of wetlands so that most upland runoff flows directly to open water bodies. The wet-dry season and tidal flooding of wetlands have been altered, especially with spoil banks created during channelization.
- Nutrient processing has changed dramatically with fertilizer application and rapid nutrient runoff, especially nitrate, that is discharged directly to open water bodies. The role of wetlands in buffering nutrient concentrations and stoichiometry has been greatly reduced.
- Because of high nutrient input due to wetland bypassing, water bodies in the upper and mid basins receiving high nutrient inputs have become very productive, more heterotrophic due to high organic input from wetlands and uplands, and eutrophic to hyper-eutrophic. Water bodies with low agricultural input are less productive and slightly autotrophic, and more mesotrophic. Water quality in the upper basin is highly degraded while that in the lower basin is less so.
- Wetlands in the basin have become degraded and there has been extensive wetland loss in the lower basin due to increasing water levels, lack of riverine input, saltwater intrusion, and pervasive alteration of hydrology. Marsh productivity has declined due to these human impacts. The impacts of oil and gas activity have contributed significantly to marsh loss. The upper basin is dominated by bald cypress-water tupelo swamps and some freshwater marshes, but there has been relatively little loss of wetland area. However, forested wetlands are now mostly permanently flooded so that productivity is declining, and recruitment is limited so that there is a slow deterioration of these wetlands. Low to no natural recruitment because of increased flooding has made logging unsustainable.
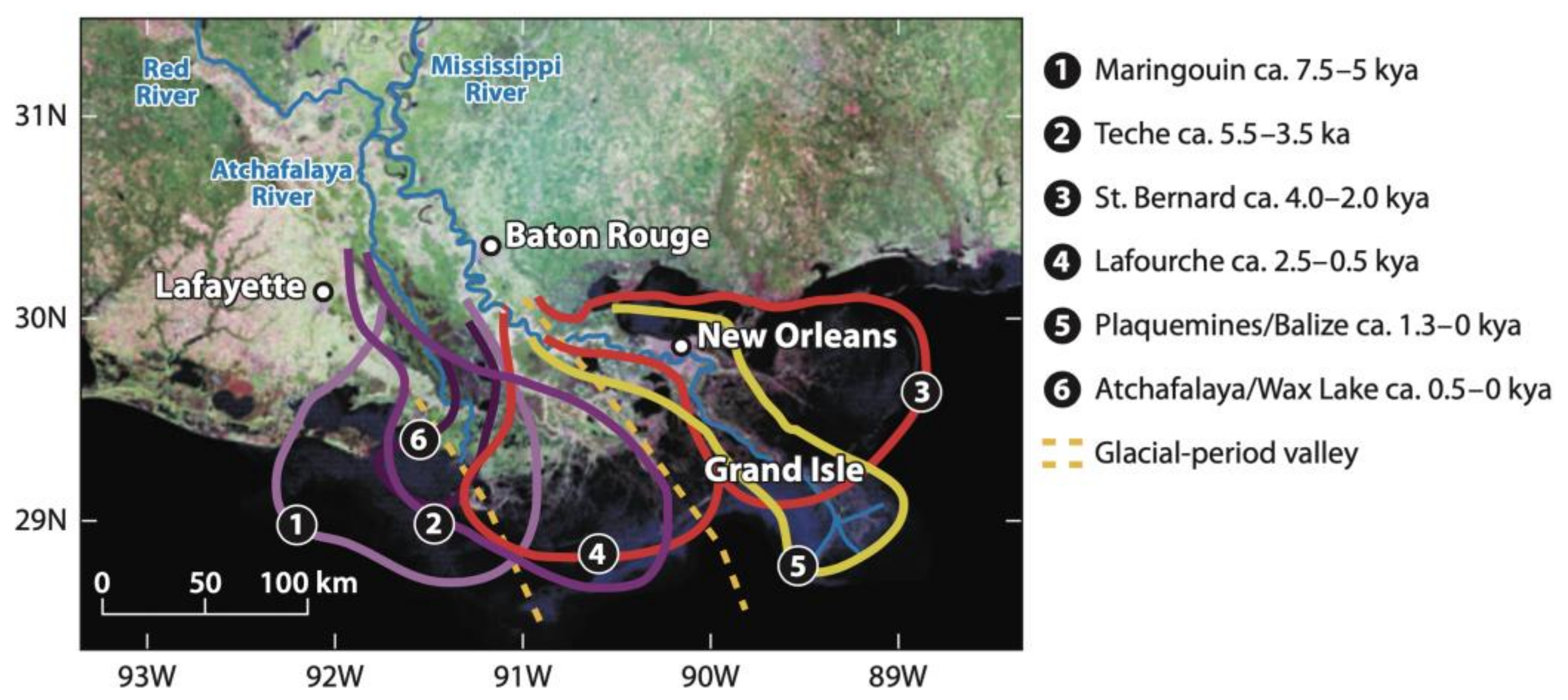
3. Description of the Barataria Basin
Wetland Change in the Barataria Basin
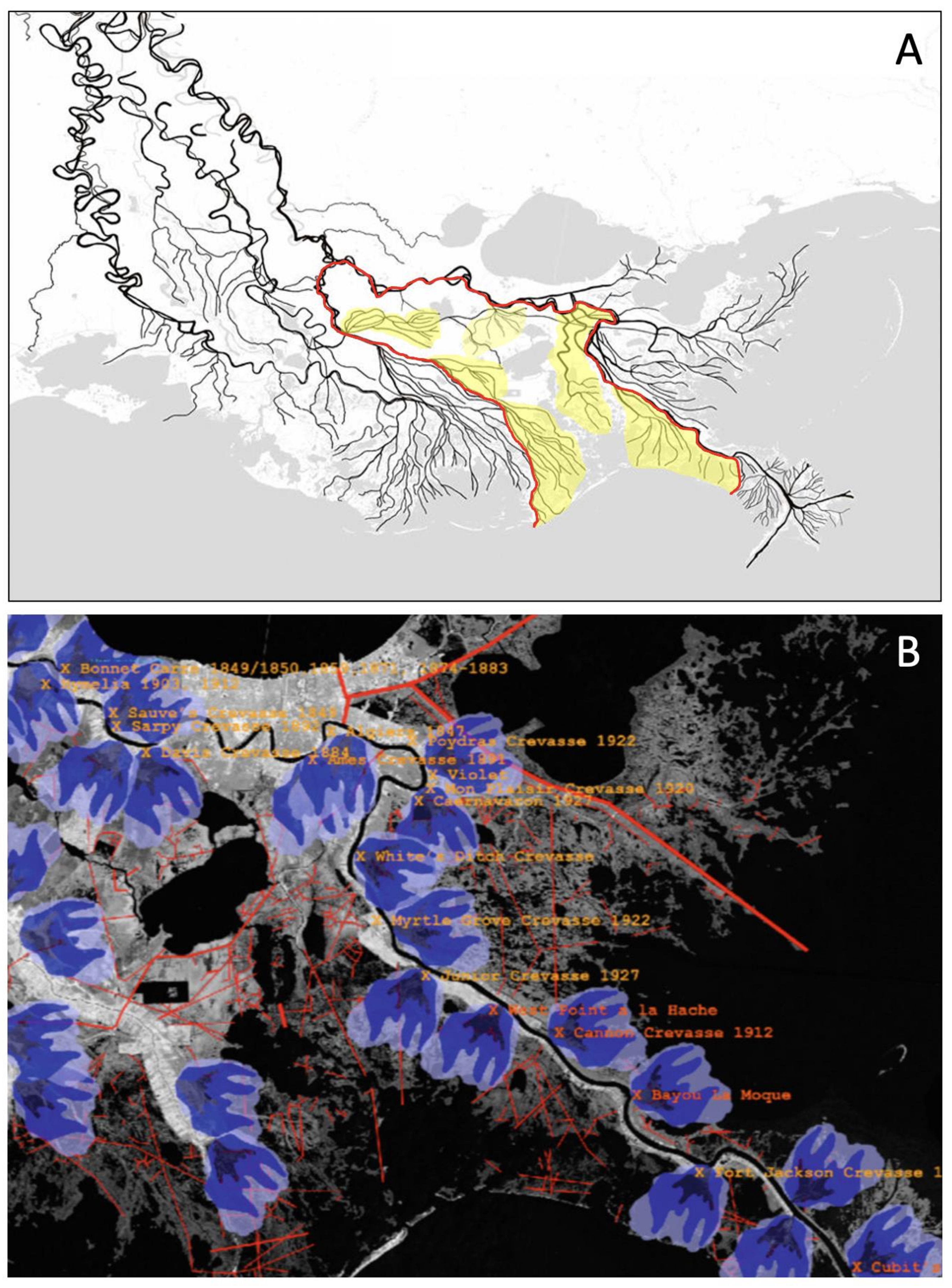
4. Pervasive Changes in Hydrology
5. Water Quality, Biogeochemistry, and Changes in the Trophic Status of Water Bodies of the Barataria Basin from the 1970s to 2015
5.1. Temporal Changes in the Water Quality of the Basin Over Four Decades
5.2. Wetland-Nutrient Interactions in Upper Barataria Basin
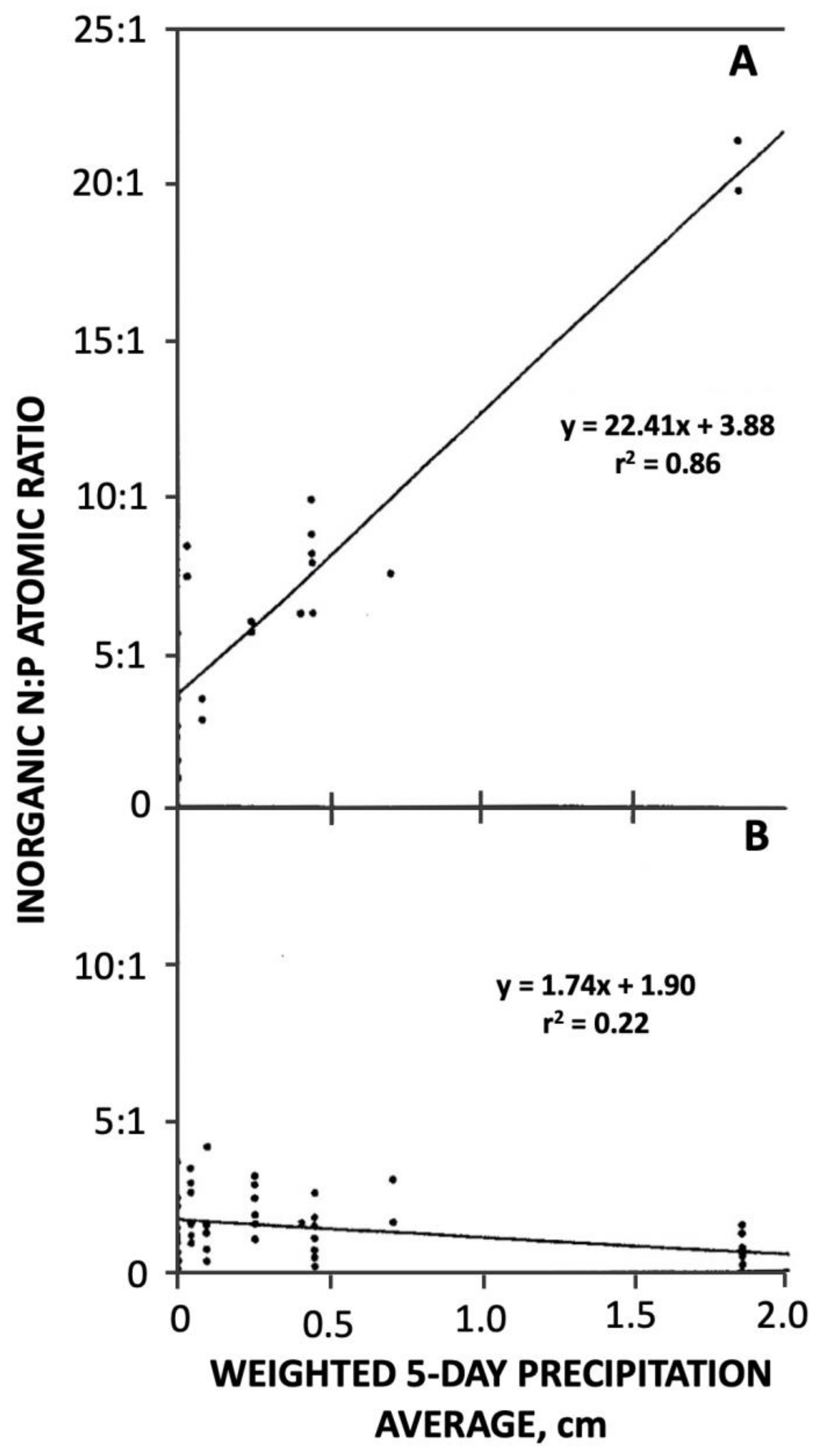
5.3. Non-Conservative Nutrient Dynamics in Barataria Basin
5.4. Changes in the Trophic Status of the Barataria Basin over Four Decades
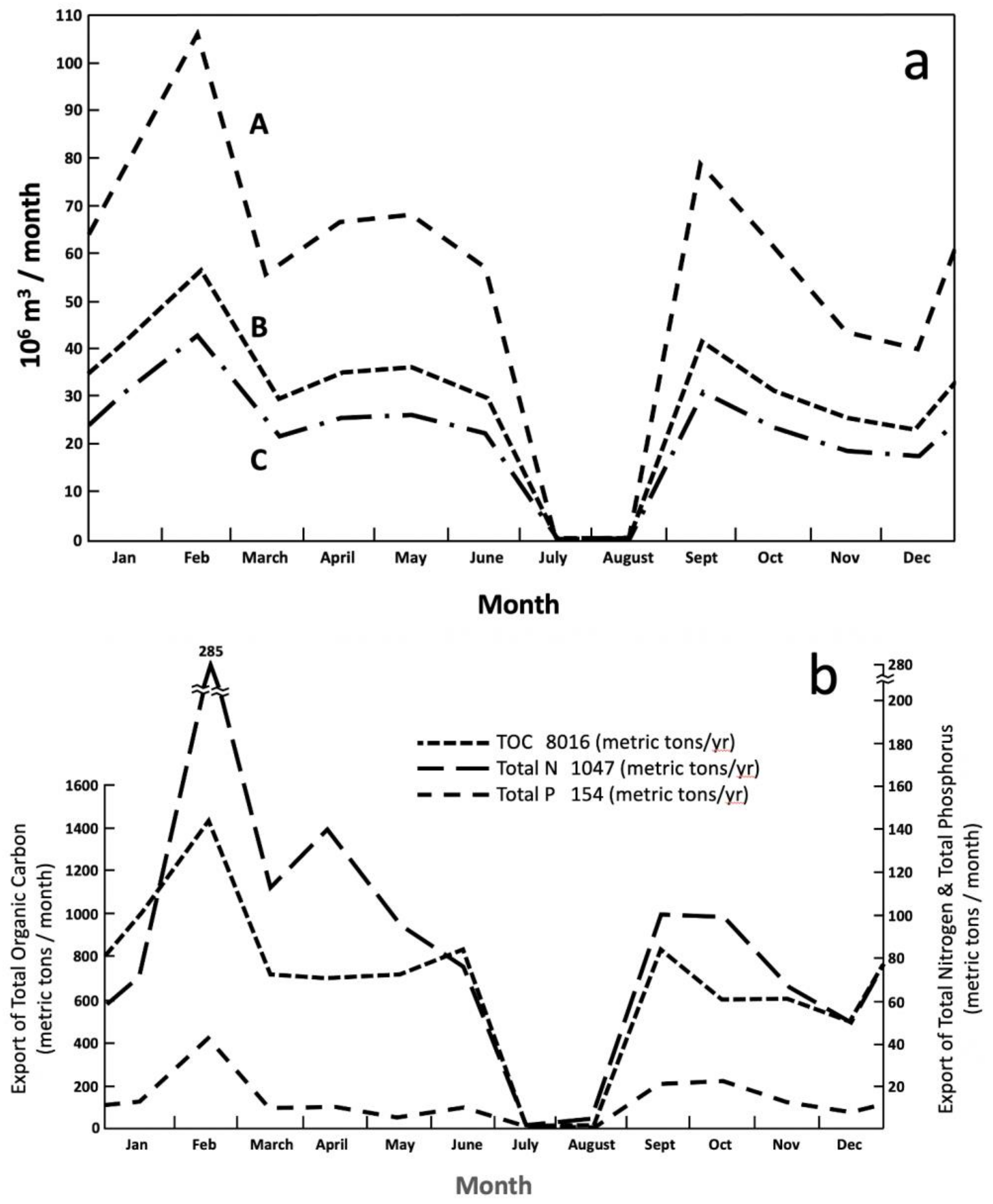
6. Aquatic Metabolism and Materials Export in Barataria Basin
7. Structure and Productivity of Wetlands in Barataria Basin
7.1. Forested Wetlands
7.2. Herbaceous Wetlands
8. Synthesis of System Functioning and Change
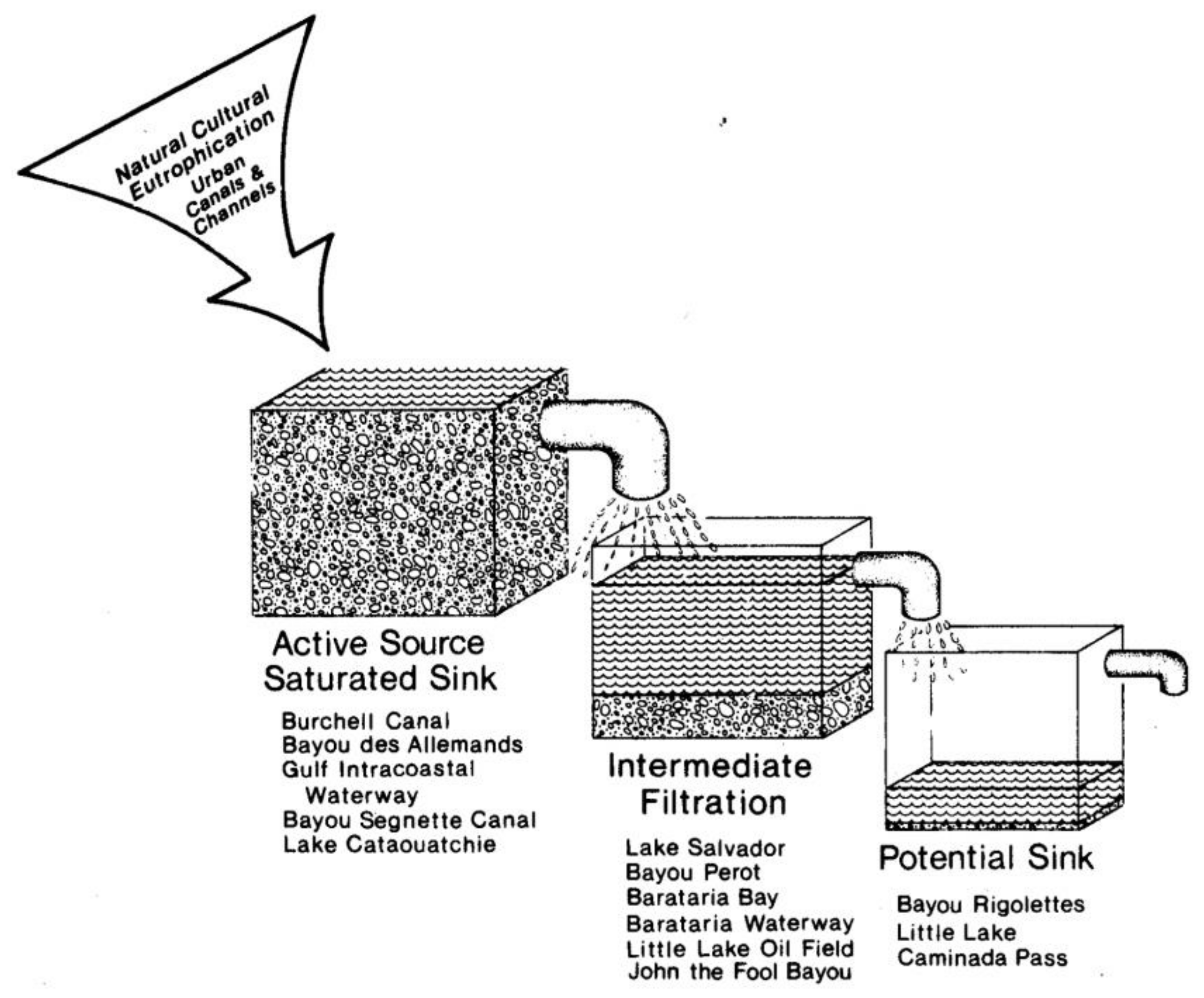
8.1. Source–Sink Model of Nutrient Dynamics and Trophic Status
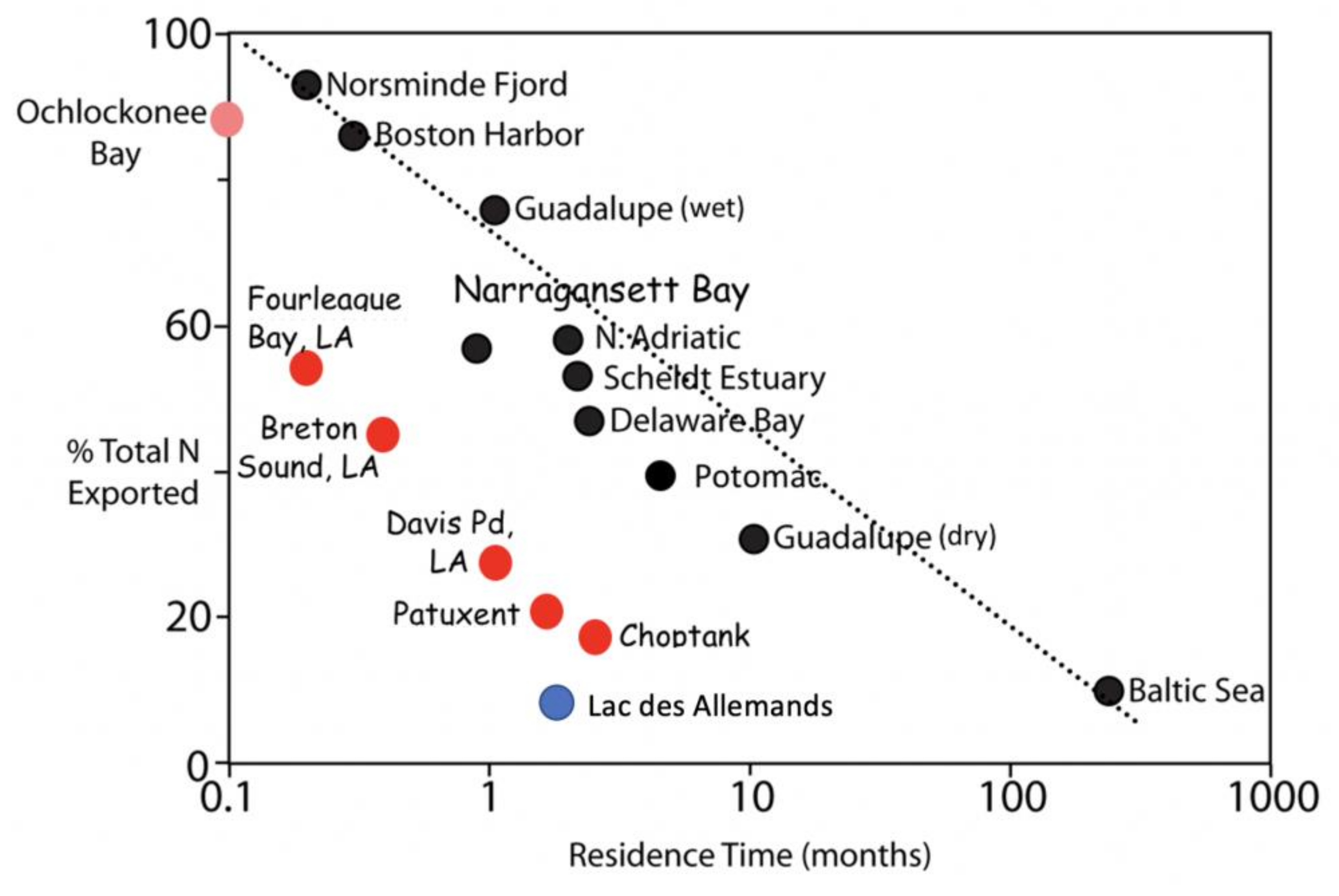
8.2. Freshwater Residence Time and Nitrogen Export
8.3. The Impact of the Davis Pond Diversion on Wetlands and Waters of Mid Barataria Basin
9. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kemp, P. Mud Deposition at the Shoreface: Wave and Sediment Dynamics on the Chenier Plain of Louisiana. Ph.D. Dissertation, Louisiana State University, Baton Rouge, LA, USA, May 1987. [Google Scholar]
- Roberts, H.H. Dynamic changes of the holocene Mississippi River delta plain: The delta cycle. J. Coast. Res. 1997, 13, 605–627. [Google Scholar]
- Hijma, M.; Shen, Z.; Törnqvist, T.; Mauz, B. Late Holocene evolution of a coupled, mud-dominated delta plain-chenier plain systems, coastal Louisiana, USA. Earth Surf. Dynam. 2018, 5, 689–710. [Google Scholar] [CrossRef] [Green Version]
- Couvillion, B.R.; Fischer, M.R.; Beck, H.J.; Sleavin, W.J. Spatial Configuration Trends in Coastal Louisiana from 1985 to 2010. Wetlands 2016, 36, 347–359. [Google Scholar] [CrossRef]
- Stagg, C.L.; Osland, M.J.; Moon, J.A.; Hall, C.T.; Feher, L.C.; Jones, W.R.; Couvillion, B.R.; Hartley, S.B.; Vervaeke, W.C. Quantifying hydrologic controls on local-and landscape-scale indicators of coastal wetland loss. Ann. Bot. 2020, 125, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Day, J.W.; Smith, W.; Stowe, W.; Wagner, P. Community Structure and Carbon Budget of a Salt Marsh and Shallow Bay Estuarine System in Louisiana; Publ. No. LSU-SG-72-04; Center for Wetland Resources, Louisiana State University: Baton Rouge, LA, USA, 1973; p. 79. [Google Scholar]
- Conner, W.H.; Day, J.W. (Eds.) The Ecology of Barataria Basin, Louisiana: An Estuarine Profile; Biological Report; National Wetlands Research Center, U.S. Fish and Wildlife Service: Washington, DC, USA, 1987; Volume 85, 165p.
- Blum, M.D.; Roberts, H.H. The Mississippi delta region: Past, present, and future. Annu. Rev. Earth Planet. Sci. 2012, 40, 655–683. [Google Scholar] [CrossRef]
- Condrey, R.E.; Hoffman, P.E.; Evers, D.E. The last naturally active delta complexes of the Mississippi River (LNDM): Discovery and implications. In Perspectives on the Restoration of the Mississippi Delta; Day, J.W., Kemp, G.P., Freemen, A.M., Muth, D.P., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 33–50. [Google Scholar]
- Swarzenski, C.M. Surface-Water Hydrology of the Gulf Intracoastal Waterway in South-Central Louisiana, 1996–1999; Professional Paper 1672; U.S. Geological Survey: Reston, VA, USA, 2003; p. 51.
- USACOE (U.S. Army Corps of Engineers). Freshwater Diversions; USACE: New Orleans, LA, USA, 2006. [Google Scholar]
- Coleman, D.J.; Kolker, A.S.; Johannesson, K.H. Submarine groundwater discharge and alkaline earth element dynamics in a deltaic coastal setting. Hydrol. Res. 2017, 48, 1169–1176. [Google Scholar] [CrossRef]
- Hopkinson, C.S., Jr.; Day, J.W., Jr.; Kjerfve, B. Ecological significance of summer storms on shallow coastal lake and estuarine systems. Cont. Mar. Sci. 1985, 28, 69–77. [Google Scholar]
- Wiseman, W.J.; Swenson, E.M.; Power, J. Salinity trends in Louisiana estuaries. Estuaries 1990, 13, 265–271. [Google Scholar] [CrossRef]
- Wissel, B.; Gaçe, A.; Fry, B. Tracing river influences on phytoplankton dynamics in two Louisiana estuaries. Ecology 2005, 86, 2751–2762. [Google Scholar] [CrossRef]
- Turner, R.E.; Swenson, E.M.; Milan, C.S.; Lee, J.M. Spatial variations in Chlorophyll a, C, N, and P in a Louisiana estuary from 1994 to 2016. Hydrobiologia 2019, 834, 131–144. [Google Scholar] [CrossRef] [Green Version]
- Couvillion, B.R.; Beck, H.; Schoolmaster, D.; Fischer, M. Land Area Change in Coastal Louisiana 1932 to 2016; Scientific Investigations Map 3381; U.S. Geological Survey: Reston, VA, USA, 2017; p. 16.
- Day, J.W.; Li, B.; Marx, B.D.; Zhao, D.; Lane, R.R. Multivariate analyses of water quality dynamics over four decades in the Barataria Basin, Mississippi Delta. Water 2020, 12, 3143. [Google Scholar] [CrossRef]
- Britsch, L.D.; Dunbar, J.B. Land loss rates: Louisiana Coastal Plain. J. Coast. Res. 1993, 9, 324–338. [Google Scholar]
- Britsch, L.D.; Dunbar, J.B. Land Loss in Coastal Louisiana 1932 to 2001. In A Series of 7 Large Format Maps; Technical Report ERDC/GSL TR-05-13; Engineer Research and Development Center: Vicksburg, MS, USA, 2006. [Google Scholar]
- Couvillion, B.R.; Barras, J.A.; Steyer, G.D.; Sleavin, W.; Fischer, M.; Beck, H.; Trahan, N.; Griffin, B.; Heckman, D. Land Area Change in Coastal Louisiana from 1932 to 2010. Scientific Investigations, Map 3164, Scale 1:265,000; U.S. Geological Survey: Reston, VA, USA, 2011; p. 12.
- Day, J.W.; Britsch, L.D.; Hawes, S.R.; Shaffer, G.P.; Reed, D.J.; Cahoon, D. Pattern and process of land loss in the Mississippi delta: A spatial and temporal analysis of wetland habitat change. Estuaries 2000, 23, 425–438. [Google Scholar] [CrossRef]
- Day, J.W.; Boesch, D.F.; Clairain, E.J.; Kemp, G.P.; Laska, S.B.; Mitsch, W.J.; Orth, K.; Mashriqui, H.; Reed, D.J.; Shabman, L.; et al. Restoration of the Mississippi Delta: Lessons from hurricanes Katrina and Rita. Science 2007, 315, 1679–1684. [Google Scholar] [CrossRef] [Green Version]
- Day, J.W.; Shaffer, G.P.; Cahoon, D.R.; DeLaune, R.D. Canals, backfilling and wetland loss in the Mississippi Delta. Estuar. Coast. Shelf Sci. 2019, 227, 106325. [Google Scholar] [CrossRef]
- Day, J.W.; Kemp, G.P.; Reed, D.J.; Cahoon, D.R.; Boumans, R.M.; Suhayda, J.M.; Gambrell, R. Vegetation death and rapid loss of surface elevation in two contrasting Mississippi delta salt marshes: The role of sedimentation, autocompaction and sea-level rise. Ecol. Eng. 2011, 37, 229–240. [Google Scholar] [CrossRef]
- Elsey-Quirk, T.; Graham, S.A.; Mendelssohn, I.A.; Snedden, G.; Day, J.W.; Twilley, R.R.; Shaffer, G.; Sharp, L.A.; Pahl, J.; Lane, R.R. Mississippi river sediment diversions and coastal wetland sustainability: Synthesis of responses to freshwater, sediment, and nutrient inputs. Estuar. Coast. Shelf Sci. 2019, 221, 170–183. [Google Scholar] [CrossRef]
- Fisk, H.N. Geological Investigation of the Alluvial Valley of the Lower Mississippi River. Illus; U.S. Army Corps of Engineers: Washington, DC, USA, 1944; p. 78. [Google Scholar]
- Davis, D.W. Crevasses on the lower course of the Mississippi River. Coastal Zone’93. In Proceedings of the Eighth Symposium on Coastal and Ocean Management, New Orleans, LA, USA, 19–23 July 1993; American Society of Civil Engineers: Reston, VA, USA, 1993; pp. 360–378. [Google Scholar]
- Davis, D.W. Historical perspective on crevasses, levees, and the Mississippi River. In Transforming New Orleans and Its Environs: Centuries of Change; Colten, C.E., Ed.; University of Pittsburgh Press: Pittsburgh, PA, USA, 2000; pp. 84–106. [Google Scholar]
- Day, J.W.; Cable, J.E.; Lane, R.R.; Kemp, G.P. Sediment deposition at the Caernarvon crevasse during the great Mississippi flood of 1927: Implications for coastal restoration. Water 2016, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- Day, J.W.; Agboola, J.; Chen, Z.; D’Elia, C.; Forbes, D.L.; Giosan, L.; Kemp, P.; Kuenzer, C.; Lane, R.R.; Ramachandran, R.; et al. Approaches to defining deltaic sustainability in the 21st century. Estuar. Coast. Shelf Sci. 2016, 183, 275–291. [Google Scholar] [CrossRef]
- Day, J.W., Jr.; Butler, T.J.; Conner, W.H. Productivity and nutrient export studies in a cypress swamp and lake system in Louisiana. In Estuarine Processes: Circulation, Sediments, and Transfer of Material in the Estuary; Academic Press: New York, NY, USA, 1977; Volume 2, pp. 255–269. [Google Scholar]
- Rosenzweig, J. Reinventing the Mississippi River Delta: A Flood of Creativity. Master’s Thesis, Tulane University School of Architecture, New Orleans, LA, USA, 2007. [Google Scholar]
- Lane, R.R.; Kemp, G.P.; Day, J.W. A Brief History of Delta Formation and Deterioration. In Multifunctional Wetlands; Nagabhatla, N., Metcalfe, C.D., Eds.; Springer: Cham, Switzerland, 2018; pp. 11–27. [Google Scholar]
- Visser, J.M.; Sasser, C.E.; Chabreck, R.H.; Linscombe, R.G. The impact of a severe drought on the vegetation of a subtropical estuary. Estuaries 2002, 25, 1184–1195. [Google Scholar] [CrossRef]
- Day, J.; Hunter, R.; Keim, R.F.; DeLaune, R.; Shaffer, G.; Evers, E.; Reed, D.; Brantley, C.; Kemp, P.; Day, J.; et al. Ecological response of forested wetlands with and without large-scale Mississippi River input: Implications for management. Ecol. Eng. 2012, 46, 57–67. [Google Scholar] [CrossRef]
- Edwards, B.L.; Allen, S.T.; Braud, D.H.; Keim, R.F. Stand density and carbon storage in cypress-tupelo wetland forests of the Mississippi River delta. For. Ecol. Manag. 2019, 441, 106–114. [Google Scholar] [CrossRef]
- Hsueh, Y.H.; Chambers, J.L.; Krauss, K.W.; Allen, S.T.; Keim, R.F. Hydrologic exchanges and baldcypress water use on deltaic hummocks, Louisiana, USA. Ecohydrology 2016, 9, 1452–1463. [Google Scholar] [CrossRef]
- Needham, H.F.; Keim, B.D.; Sathiaraj, D. A review of tropical cyclone-generated storm surges: Global data sources, observations, and impacts. Rev. Geophys. 2015, 53, 545–591. [Google Scholar] [CrossRef]
- Doyle, T.W.; Conner, W.H.; Day, R.H.; Krauss, K.W.; Swarzenski, C.M. Wind damage and salinity effects of Hurricanes Katrina and Rita on coastal baldcypress forests of Louisiana. Plant Environ. Sci. 2007, 12, 163–168. [Google Scholar]
- Keim, R.F.; Lemon, M.G.T.; Oakman, E.C. Post hurricane salinity in an impounded coastal wetland (Bayou Sauvage, Louisiana, USA). J. Coast. Res. 2019, 35, 1003–1009. [Google Scholar] [CrossRef]
- Gael, B.T.; Hopkinson, C.S. Drainage density, land-use and eutrophication in Barataria Basin, Louisiana. In Proceedings of the Third Marsh and Estuary Management Symposium, Baton Rouge, LA, USA, 6–7 March 1978; Day, J., Culley, D., Turner, R., Mumphrey, A., Eds.; Louisiana State University Division of Continuing Education: Baton Rouge, LA, USA, 1979; pp. 147–163. [Google Scholar]
- Brezonik, P.; Shannon, E. Trophic state of lakes in north central Florida. In Florida Water Resources Research Center; Publ. No. 13; University of Florida: Gainesville, FL, USA, 1971. [Google Scholar]
- Day, J.W.; Hopkinson, C.S.; Conner, W.H. An analysis of environmental factors regulating community metabolism and fisheries production in a Louisiana estuary. In Estuarine Comparisons; Kennedy, V., Ed.; Academic Press: New York, NY, USA, 1982; pp. 121–136. [Google Scholar]
- Seaton, A. Nutrient Chemistry in the Barataria Basin—A Multivariant Approach. Master’s Thesis, Louisiana State University, Baton Rouge, LA, USA, 1979; 123p. [Google Scholar]
- Seaton, A.; Day, J. The Development of a Trophic State Index for the Quantification of Eutrophication in the Barataria Basin. In Proceedings of the Third Marsh and Estuary Management Symposium, Baton Rouge, LA, USA, 6–7 March 1978; Day, J., Culley, D., Turner, R., Mumphrey, A., Eds.; Louisiana State University Division of Continuing Education: Baton Rouge, LA, USA, 1979; pp. 113–125. [Google Scholar]
- Witzig, A.; Day, J. A Tropic State Index for the Louisiana Coastal Zone; Technical Completion Report to the LA; Water Resources Research Institute, Louisiana State University: Baton Rouge, LA, USA, 1983; 35p. [Google Scholar]
- Yu, K.; DeLaune, R.D.; Tao, R.; Beine, R.L. Nonpoint source of nutrients and herbicides associated with sugarcane production and its impact on Louisiana coastal water quality. J. Environ. Qual. 2008, 37, 2275–2283. [Google Scholar] [CrossRef] [Green Version]
- DeLaune, R.; Yu, K.; Devai, I.; Tao, R. Water Quality of Upper Barataria Basin: Impact of Nonpoint Source Pollution Associated with Sugarcane Production (St. James Sugarcane Run-off Project); Final Report DEQ Contract No. CFMS597, Grant number C9-996102-08; Louisiana State University: Baton Rouge, LA, USA, 2007; 104p. [Google Scholar]
- Kemp, P.; Day, J. Nutrient dynamics in a swamp receiving agricultural runoff. In Cypress Swamps; Ewel, K., Odum, H.T., Eds.; University of Florida Press: Gainesville, FL, USA, 1984; pp. 286–293. [Google Scholar]
- Lane, R.R.; Huang, H.; Day, J.W.; Justic, D.; DeLaune, R.D. Water quality of a coastal Louisiana swamp and how dredging is undermining restoration efforts. Estuar. Coast. Shelf Sci. 2015, 152, 23–32. [Google Scholar] [CrossRef]
- Day, J.W.; Kemp, P. Long term impacts of agricultural runoff in a Louisiana swamp forest. In Ecological Considerations in Wetlands Treatment of Municipal Waste Water; Godfrey, P., Ed.; Van Norstrand Reinhold Co.: New York, NY, USA, 1985; pp. 317–326. [Google Scholar]
- Kemp, G.P.; Conner, W.H.; Day, J.W. Effects of flooding on decomposition and nutrient cycling in a Louisiana swamp forest. Wetlands 1985, 5, 35–51. [Google Scholar] [CrossRef]
- Rivera-Monroy, V.H.; Lenaker, P.; Twilley, R.R.; Delaune, R.D.; Lindau, C.W.; Nuttle, W.; Habib, E.; Fulweiler, R.W.; Castañeda-Moya, E. Denitrification in coastal Louisiana: A spatial assessment and research needs. J. Sea Res. 2010, 63, 157–172. [Google Scholar] [CrossRef]
- Rivera-Monroy, V.H.; Branoff, B.; Meselhe, E.; McCorquodale, A.; Dortch, M.; Steyer, G.D.; Visser, J.; Wang, H. Landscape-level estimation of nitrogen removal in coastal Louisiana wetlands: Potential sinks under different restoration scenarios. J. Coast. Res. 2013, 67, 75–87. [Google Scholar] [CrossRef]
- Upreti, K.; Rivera-Monroy, V.H.; Maiti, K.; Giblin, A.; Geaghan, J.P. Emerging wetlands from river diversions can sustain high denitrification rates in a coastal delta. JBR Biogeosci. 2021. (in review). [Google Scholar] [CrossRef]
- Vaccare, J.; Meselhe, E.; White, J.R. The denitrification potential of eroding wetlands in Barataria Bay, LA, USA: Implications for river reconnection. Sci. Total Environ. 2019, 686, 529–537. [Google Scholar] [CrossRef]
- Vargas-Lopez, I.A.; Rivera-Monroy, V.H.; Day, J.W.; Whitbeck, J.; Maiti, K.; Madden, C.J.; Castro, A.T. Assessing chlorophyll a spatiotemporal patterns combining in situ continuous fluorometry measurements and Landsat 8/OLI data across the Barataria Basin (Louisiana, USA). Water 2021. (in review). [Google Scholar] [CrossRef]
- Day, J.W., Jr.; Templet, P.H. Consequences of sea level rise: Implications from the Mississippi Delta. Coast. Manag. 1989, 17, 241–257. [Google Scholar] [CrossRef]
- Lane, R.R.; Mack, S.K.; Day, J.W.; Kempka, R.; Brady, L.J. Carbon sequestration at a forested wetland receiving treated municipal effluent. Wetlands 2017, 37, 861–873. [Google Scholar] [CrossRef]
- Lane, R.R.; Mack, S.K.; Day, J.W.; DeLaune, R.D.; Madison, M.J.; Precht, P.R. Fate of soil organic carbon during wetland loss. Wetlands 2016, 36, 1167–1181. [Google Scholar] [CrossRef]
- Lane, R.R.; Madden, C.J.; Day, J.W.; Solet, D.J. Hydrologic and nutrient dynamics of a coastal bay and wetland receiving discharge from the Atchafalaya River. Hydrobiologia 2011, 658, 55–66. [Google Scholar] [CrossRef]
- Randall, J.M.; Day, J.W. Effects of river discharge and vertical circulation on aquatic primary production in a turbid Louisiana (USA) estuary. Neth. J. Sea Res. 1987, 21, 231–242. [Google Scholar] [CrossRef]
- Madden, C.J. Control of Phytoplankton Production in a Shallow, Turbid Estuary. Ph.D. Dissertation, Louisiana State University, Baton Rouge, LA, USA, 1992. [Google Scholar]
- Madden, C.J.; Day, J.W., Jr.; Randall, J.M. Freshwater and marine coupling in estuaries of the Mississippi River deltaic plain 1. Limnol. Oceanogr. 1988, 33, 982–1004. [Google Scholar] [CrossRef]
- Happ, G.; Gosselink, J.G.; Day, J.W., Jr. The seasonal distribution of organic carbon in a Louisiana estuary. Estuar. Coast. Mar. Sci. 1977, 5, 695–705. [Google Scholar] [CrossRef]
- Das, A. Modeling the Impacts of Pulsed Riverine Inflows on Hydrodynamics and Water Quality in the Barataria Bay Estuary. Ph.D. Dissertation, Louisiana State University, Baton Rouge, LA, USA, 2010. [Google Scholar]
- Das, A.; Justić, D.; Swenson, E. Modeling estuarine-shelf exchanges in a deltaic estuary: Implications for coastal carbon budgets and hypoxia. Ecol. Model. 2010, 221, 978–985. [Google Scholar] [CrossRef]
- Norgress, R.E. The History of the Cypress Lumber Industry in Louisiana. Master’s Thesis, Louisiana State University, Baton Rouge, LA, USA, 1936. [Google Scholar]
- Mancil, E.M. An Historical Geography of Industrial Cypress Lumbering. Ph.D. Dissertation, Louisiana State University, Baton Rouge, LA, USA, 1972. [Google Scholar]
- Anderson, R.C.; White, J. A cypress swamp outlier in southern Illinois. Ill. State Acad. Sci. 1970, 63, 6–13. [Google Scholar]
- Conner, W.H.; Day, J.W. Productivity and composition of a baldcypress-water tupelo site and a bottomland hardwood site in a Louisiana swamp. Am. J. Bot. 1976, 63, 1354–1364. [Google Scholar] [CrossRef]
- Conner, W.H.; Gosselink, J.G.; Parrondo, R.T. Comparison of the vegetation of three Louisiana swamp sites with different flooding regimes. Am. J. Bot. 1981, 68, 320–331. [Google Scholar] [CrossRef]
- Conner, W.H.; Day, J.W. Water level variability and litterfall productivity of forested freshwater wetlands in Louisiana. Am. Midl. Nat. 1992, 128, 237–245. [Google Scholar] [CrossRef]
- Conner, W.H.; Mihalia, I.; Wolfe, J. Tree community structure and changes from 1987 to 1999 in three Louisiana and three South Carolina forested wetlands. Wetlands 2002, 22, 58–70. [Google Scholar] [CrossRef]
- Nienhuis, J.H.; Törnqvist, T.E.; Jankowski, K.L.; Fernandes, A.M.; Keogh, M.E. A new subsidence map for coastal Louisiana. GSA Today 2017, 27, 58–59. [Google Scholar]
- Conner, W.H.; Day, J.W. Rising water levels in coastal Louisiana: Implications for two coastal forested wetland areas in Louisiana. J. Coast. Res. 1988, 4, 589–596. [Google Scholar]
- Conner, W.H.; Brody, M. Rising water levels and the future of southeastern Louisiana swamp forests. Estuaries 1989, 12, 318–323. [Google Scholar] [CrossRef]
- Keim, R.F.; Izdepski, C.W.; Day, J.W. Growth responses of baldcypress to wastewater nutrient additions and changing hydrologic regime. Wetlands 2012, 32, 95–103. [Google Scholar] [CrossRef]
- Allen, S.T.; Keim, R.F.; Dean, T.J. Contrasting effects of flooding on tree growth and stand density determine aboveground production, in baldcypress forests. For. Ecol. Manag. 2019, 432, 345–355. [Google Scholar] [CrossRef]
- Howard, J.J. Hurricane Katrina impact on a leveed bottomland hardwood forest in Louisiana. Am. Midl. Nat. 2012, 168, 56–69. [Google Scholar] [CrossRef]
- Conner, W.H.; Duberstein, J.A.; Day, J.W.; Hutchinson, S. Impacts of changing hydrology and hurricanes on forest structure and growth along a flooding/elevation gradient in a south Louisiana forested wetland from 1986 to 2009. Wetlands 2014, 34, 803–814. [Google Scholar] [CrossRef]
- Shaffer, G.P.; Kandalepas, D. Monitoring of the Lac des Allemands Swamp (BA-34-2) Hydrologic Restoration Project; Final Report; Wetland Resources, LLC: Ponchatoula, LA, USA, 2016. [Google Scholar]
- Shaffer, G.P.; Kandalepas, D. Monitoring of the Lac des Allemands Swamp (BA-34-2) Hydrologic Restoration Project; Final Report; Wetland Resources, LLC: Ponchatoula, LA, USA, 2019. [Google Scholar]
- Day, J.W.; Ko, J.Y.; Rybczyk, J.; Sabins, D.; Bean, R.; Berthelot, G.; Brantley, C.; Cardoch, L.; Conner, W.; Day, J.N.; et al. The use of wetlands in the Mississippi Delta for wastewater assimilation: A review. Ocean Coast. Manag. 2004, 47, 671–691. [Google Scholar] [CrossRef]
- Hesse, I.D.; Day, J.W.; Doyle, T.W. Long-term growth enhancement of baldcypress (Taxodium distichum) from municipal wastewater application. Environ. Manag. 1998, 22, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Brantley, C.G.; Day, J.W., Jr.; Lane, R.R.; Hyfield, E.; Day, J.N.; Ko, J.Y. Primary production, nutrient dynamics, and accretion of a coastal freshwater forested wetland assimilation system in Louisiana. Ecol. Eng. 2008, 34, 7–22. [Google Scholar] [CrossRef]
- Hunter, R.G.; Day, J.W.; Lane, R.R.; Lindsey, J.; Day, J.N.; Hunter, M.G. Impacts of secondarily treated municipal effluent on a freshwater forested wetland after 60 years of discharge. Wetlands 2009, 29, 363–371. [Google Scholar] [CrossRef]
- Shaffer, G.P.; Day, J.W.; Hunter, R.G.; Lane, R.R.; Lundberg, C.J.; Wood, W.B.; Hillmann, E.R.; Day, J.N.; Strickland, E.; Kandalepas, D. System response, nutria herbivory, and vegetation recovery of a wetland receiving secondarily-treated effluent in coastal Louisiana. Ecol. Eng. 2015, 79, 120–131. [Google Scholar] [CrossRef]
- Sasser, C.E.; Visser, J.M.; Mouton, E.; Linscombe, J.; Hartley, S.B. Vegetation Types in Coastal Louisiana in 2013; Open-File Report; U.S. Geological Survey: Reston, VA, USA, 2014.
- O’Neil, T. The Muskrat in the Louisiana Marshes; Louisiana Wildlife and Fisheries Commission: New Orleans, LA, USA, 1949.
- Sasser, C.E.; Gosselink, J.G.; Swenson, E.M.; Swarzenski, C.M.; Leibowitz, N.C. Vegetation, substrate and hydrology in floating marshes in the Mississippi river delta plain wetlands, USA. Vegetatio 1996, 122, 129–142. [Google Scholar] [CrossRef]
- Sasser, C.E.; Visser, J.M.; Evers, D.E.; Gosselink, J.G. The role of environmental variables on interannual variation in species composition and biomass in a subtropical minerotrophic floating marsh. Can. J. Bot. 1995, 73, 413–424. [Google Scholar] [CrossRef]
- Sasser, C.E.; Gosselink, J.G.; Swenson, E.M.; Evers, D.E. Hydrologic, vegetation, and substrate characteristics of floating marshes in sediment-rich wetlands of the Mississippi river delta plain, Louisiana, USA. Wetl. Ecol. Manag. 1995, 3, 71–187. [Google Scholar] [CrossRef]
- Sasser, C.E.S.; Swenson, E.; Evers, D.E.; Visser, J.M.; Holm, G.O.; Gosselink, J.G. Floating Marshes in the Barataria and Terrebonne Basins, Louisiana; LSU-CEI-94-02; Coastal Ecology Institute, Louisiana State University: Baton Rouge, LA, USA, 1994. [Google Scholar]
- Sasser, C.E.; Gosselink, J.G. Vegetation and primary production in a floating freshwater marsh in Louisiana. Aquat. Bot. 1984, 20, 245–255. [Google Scholar] [CrossRef]
- Sasser, C.E.; Gosselink, J.G.; Shaffer, G.P. Distribution of nitrogen and phosphorus in a Louisiana freshwater floating marsh. Aquat. Bot. 1991, 41, 317–331. [Google Scholar] [CrossRef]
- Reddy, K.R.; DeLaune, R.D. Biogeochemistry of Wetlands: Science and Applications; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Nixon, S.W.; Ammerman, J.W.; Atkinson, L.P.; Berounsky, V.M.; Billen, G.; Boicourt, W.C.; Boynton, W.R.; Church, T.M.; Ditoro, D.M.; Elmgren, R.; et al. The fate of nitrogen and phosphorus at the land-sea margin of the North Atlantic Ocean. Biogeochemistry 1996, 35, 141–180. [Google Scholar] [CrossRef]
- Dettmann, E.H. Effect of water residence time on annual export and denitrification of nitrogen in estuaries: A model analysis. Estuaries 2001, 24, 481–490. [Google Scholar] [CrossRef]
- Smith, C.J.; DeLaune, R.D.; Patrick, W.H. Fate of riverine nitrate entering an estuary: I. Denitrification and nitrogen burial. Estuaries 1985, 8, 15–21. [Google Scholar] [CrossRef]
- Penland, S.; Ramsey, K.E. Relative sea-level rise in Louisiana and the Gulf of Mexico: 1908-1988. J. Coast. Res. 1990, 6, 323–342. [Google Scholar]
- Lane, R.R.; Day, J.W.; Justic, D.; Reyes, E.; Marx, B.; Day, J.N.; Hyfield, E. Changes in stoichiometric Si, N and P ratios of Mississippi River water diverted through coastal wetlands to the Gulf of Mexico. Estuar. Coast. Shelf Sci. 2004, 60, 1–10. [Google Scholar] [CrossRef]
- Boynton, W.R.; Hagy, J.D.; Cornwell, J.C.; Kemp, W.M.; Greene, S.M.; Owens, M.S.; Baker, J.E.; Larsen, R.K. Nutrient budgets and management actions in the Patuxent River estuary, Maryland. Estuaries Coasts 2008, 31, 623–651. [Google Scholar] [CrossRef]
- Perez, B.C.; Day, J.W.; Justic, D.; Lane, R.R.; Twilley, R.R. Nutrient stoichiometry, freshwater residence time, and nutrient retention in a river-dominated estuary in the Mississippi Delta. Hydrobiologia 2011, 658, 41–54. [Google Scholar] [CrossRef]
- Bargu, S.; Justic, D.; White, J.R.; Lane, R.; Day, J.; Paerl, H.; Raynie, R. Mississippi River diversions and phytoplankton dynamics in deltaic Gulf of Mexico estuaries: A review. Estuar. Coast. Shelf Sci. 2019, 221, 39–52. [Google Scholar] [CrossRef]
- White, J.R.; DeLaune, R.D.; Justic, D.; Day, J.W.; Pahl, J.; Lane, R.R.; Boynton, W.R.; Twilley, R.R. Consequences of Mississippi River diversions on nutrient dynamics of coastal wetland soils and estuarine sediments: A review. Estuar. Coast. Shelf Sci. 2019, 224, 209–216. [Google Scholar] [CrossRef]
- Kral, F.; Corstanje, R.; White, J.R.; Veronesi, F. A geostatistical analysis of soil properties in the Davis Pond Mississippi freshwater diversion. Soil Sci. Soc. Am. J. 2012, 76, 1107–1118. [Google Scholar] [CrossRef]
- Spera, A.C.; White, J.R.; Corstanje, R. Influence of a freshwater river diversion on sedimentation and phosphorus status in a wetland receiving basin. Estuar. Coast. Shelf Sci. 2020, 238, 106728. [Google Scholar] [CrossRef]
- McAlpin, T.O.; Letter, J.V.; Martin, S.K. A Hydrodynamic Study of Davis Pond, Near New Orleans, LA; ERDC/CHL TR-08-11; Coastal and Hydraulics Laboratory, U.S. Army Engineer Research and Development Center: Visksburg, MS, USA, 2008. [Google Scholar]
- Gardner, L.M.; White, J. Denitrification enzyme activity as an indicator of nitrate movement through a diversion wetland. Soil Sci. Soc. Am. J. 2010, 74, 1037–1047. [Google Scholar] [CrossRef]
- Keogh, M.E.; Kolker, A.S.; Snedden, G.A.; Renfro, A.A. Hydrodynamic controls on sediment retention in an emerging diversion-fed delta. Geomorphology 2019, 332, 100–111. [Google Scholar] [CrossRef]
- Lane, R.R.; Day, J.W.; Marx, B.; Reves, E.; Kemp, G.P. Seasonal and spatial water quality changes in the outflow plume of the Atchafalaya River, Louisiana, USA. Estuaries 2002, 25, 30–42. [Google Scholar] [CrossRef]
- Reed, D.; Wang, Y.; Meselhe, E.; White, E. Modeling wetland transitions and loss in coastal Louisiana under scenarios of future relative sea-level rise. Geomorphology 2020, 352, 106991. [Google Scholar] [CrossRef]
- Day, J.W.; Clark, H.C.; Chang, C.; Hunter, R.; Norman, C.R. Life Cycle of Oil and Gas Fields in the Mississippi River Delta: A Review. Water 2020, 12, 1492. [Google Scholar] [CrossRef]
- Hopkinson, C.S.; Day, J.W. Modeling the relationship between development and storm water and nutrient runoff. Environ. Manag. 1980, 4, 315–324. [Google Scholar] [CrossRef]
- Hopkinson, C.S.; Day, J.W. Modeling hydrology and eutrophication in a Louisiana swamp forest ecosystem. Environ. Manag. 1980, 4, 325–335. [Google Scholar] [CrossRef]
- CPRA (Coastal Protection and Restoration Authority of Louisiana). Louisiana’s Comprehensive Master Plan for a Sustainable Coast—2017 Draft Plan Release; CPRA: Baton Rouge, LA, USA, 2017; 169p. [Google Scholar]
- Kemp, G.P.; Day, J.W.; Rogers, J.D.; Giosan, L.; Peyronnin, N. Enhancing mud supply from the Lower Missouri River to the Mississippi River Delta USA: Dam bypassing and coastal restoration. Estuar. Coast. Shelf Sci. 2016, 183, 304–313. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ecological Zone | Example | Turnovers per Year | Mean Depth (m) | Salinity (‰) | Secchi Depth (cm) | Tidal Range (cm) | Upland to Wetland + Water Ratio |
|---|---|---|---|---|---|---|---|
| Upper Basin | Lac des Allemands | 4.6 | 2.0 | 0 | 33 | 3.2 | 1: 2.3 |
| Middle Basin | Lake Cataouatche | 1.5 | 2.0 | 0–3 | 30 | na | 1: 6.7 |
| Lake Salvador | 1.0 | 2.5 | 0–6 | 72 | 8.5 | ||
| Lower Basin | Little Lake | na | 1.5 | 0–15 | 72 | 12 | 1: 33.3 |
| Lower Barataria Bay | 14.6 | 2.0 | 10–35 | 68 | 30 |
| Parameter | Agricultural Drainage Ditches | St. James Canal | Bayou Chevreuil | Swamp Forest Runoff |
|---|---|---|---|---|
| TSS | 0–(1350) | 186–275 (1500) | 62–124 (350) | 24–29 |
| Total P | 0.1–(3.5) | 0.5–0.7 (1.0) | 0.2 (0.8) | 0.1–0.2 |
| TKN | 0.2–(16.5) | 1.9–2.2 (7.0) | 2.1–2.3 (7.0) | 1.4 |
| NOx | 0.1–(15.5 ) | 0.5–2.7 (6.0) | 0.2–2.2 (6.0) | 0.2–0.3 |
| NH4 | 0–(2.3) | 0.1–0.3 | 0.1–0.4 | 0.1 |
| Chl a | NDP | NR | GP | NCP | |
|---|---|---|---|---|---|
| Bayous | 25 | 316 | 446 | 762. | −130 |
| Lac des Allemands | 65 | 1418 | 1868 | 3286 | −450 |
| Lake Cataouatche | 50 | 876 | 1205 | 2222 | −350 |
| Lake Salvador | 12 | 402 | 602 | 1058 | −198 |
| Little Lake | 10 | 639 | 753 | 1307 | −117 |
| Brackish-Saline | 10 | 940 | 910 | 1850 | 0 to 54 |
| Offshore | 7.6 | 732 | . | . | . |
| Group | DOC (mg/L) | TOC (mg/L) | Chl a (mg/m3) |
|---|---|---|---|
| Marsh | 6.7 ± 0.2 | 8.5 ± 0.2 | 10.2 ± 0.6 |
| Upper Bay | 6.2 ± 0.3 | 7.6 ± 0.3 | 10.3 ± 0.8 |
| Lower Bay | 4.3 ± 0.3 | 4.5 ± 0.3 | 8.1 ± 0.8 |
| Offshore | 2.4 ± 0.1 | 2.8 ± 0.1 | 7.6 ± 0.8 |
| g C/m2/year | |
|---|---|
| (1) Net marsh production | 590 |
| (2) Marsh consumption | 300 |
| (3) Flushed from marsh | 290 |
| (4) Flushed from bay | 240 |
| (5) Phytoplankton and benthic algae production | 450 |
| (6) Total organic carbon available to water (4) + (5) | 690 |
| (7) Consumption in water | 430 |
| (8) Net estuary production available for export or sedimentation | 260 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Day, J.W.; Conner, W.H.; DeLaune, R.D.; Hopkinson, C.S.; Hunter, R.G.; Shaffer, G.P.; Kandalepas, D.; Keim, R.F.; Kemp, G.P.; Lane, R.R.; et al. A Review of 50 Years of Study of Hydrology, Wetland Dynamics, Aquatic Metabolism, Water Quality and Trophic Status, and Nutrient Biogeochemistry in the Barataria Basin, Mississippi Delta—System Functioning, Human Impacts and Restoration Approaches. Water 2021, 13, 642. https://doi.org/10.3390/w13050642
Day JW, Conner WH, DeLaune RD, Hopkinson CS, Hunter RG, Shaffer GP, Kandalepas D, Keim RF, Kemp GP, Lane RR, et al. A Review of 50 Years of Study of Hydrology, Wetland Dynamics, Aquatic Metabolism, Water Quality and Trophic Status, and Nutrient Biogeochemistry in the Barataria Basin, Mississippi Delta—System Functioning, Human Impacts and Restoration Approaches. Water. 2021; 13(5):642. https://doi.org/10.3390/w13050642
Chicago/Turabian StyleDay, John W., William H. Conner, Ronald D. DeLaune, Charles S. Hopkinson, Rachael G. Hunter, Gary P. Shaffer, Demetra Kandalepas, Richard F. Keim, G. Paul Kemp, Robert R. Lane, and et al. 2021. "A Review of 50 Years of Study of Hydrology, Wetland Dynamics, Aquatic Metabolism, Water Quality and Trophic Status, and Nutrient Biogeochemistry in the Barataria Basin, Mississippi Delta—System Functioning, Human Impacts and Restoration Approaches" Water 13, no. 5: 642. https://doi.org/10.3390/w13050642
APA StyleDay, J. W., Conner, W. H., DeLaune, R. D., Hopkinson, C. S., Hunter, R. G., Shaffer, G. P., Kandalepas, D., Keim, R. F., Kemp, G. P., Lane, R. R., Rivera-Monroy, V. H., Sasser, C. E., R. White, J., & Vargas-Lopez, I. A. (2021). A Review of 50 Years of Study of Hydrology, Wetland Dynamics, Aquatic Metabolism, Water Quality and Trophic Status, and Nutrient Biogeochemistry in the Barataria Basin, Mississippi Delta—System Functioning, Human Impacts and Restoration Approaches. Water, 13(5), 642. https://doi.org/10.3390/w13050642