2.1. Experimental Setup, Sites and Soils
The experimental setup is part of the Germany-wide TERENO- observatory network (TERrestrial ENvironmental Observatories;
https://www.tereno.net/ accessed 1 April 2020) that intends to provide extensive data sets from high precision non-destructive measurements of the soil–plant–atmosphere continuum [
39]. The TERENO SOILCan sub-project, implements an adapted “Space for Time” (SFT) substitution approach: lysimeters are transferred across TERENO study sites, along temperature and precipitation gradients. The translocation led to an abrupt rather than gradual climate regime variation, contrary to most SFT substitution studies. Rather than following a gradual change, the aim is to compare the transferred soil with the soil at the original site. It is thus possible to account for unsuspected effects from the past [
40,
41]. The high-precision weighable SOILCan lysimeters are used to observe the effect of complex processes on the soil–plant–atmosphere relations including soil water and nitrogen dynamics [
39]. This way, effects of climatic shifts on the soil ecosystem can be observed under actual climatic conditions for Germany. For more information on the TERENO-SOILCan network and its adapted SFT substitution approach see Pütz et al. [
39] andGroh et al. [
40].
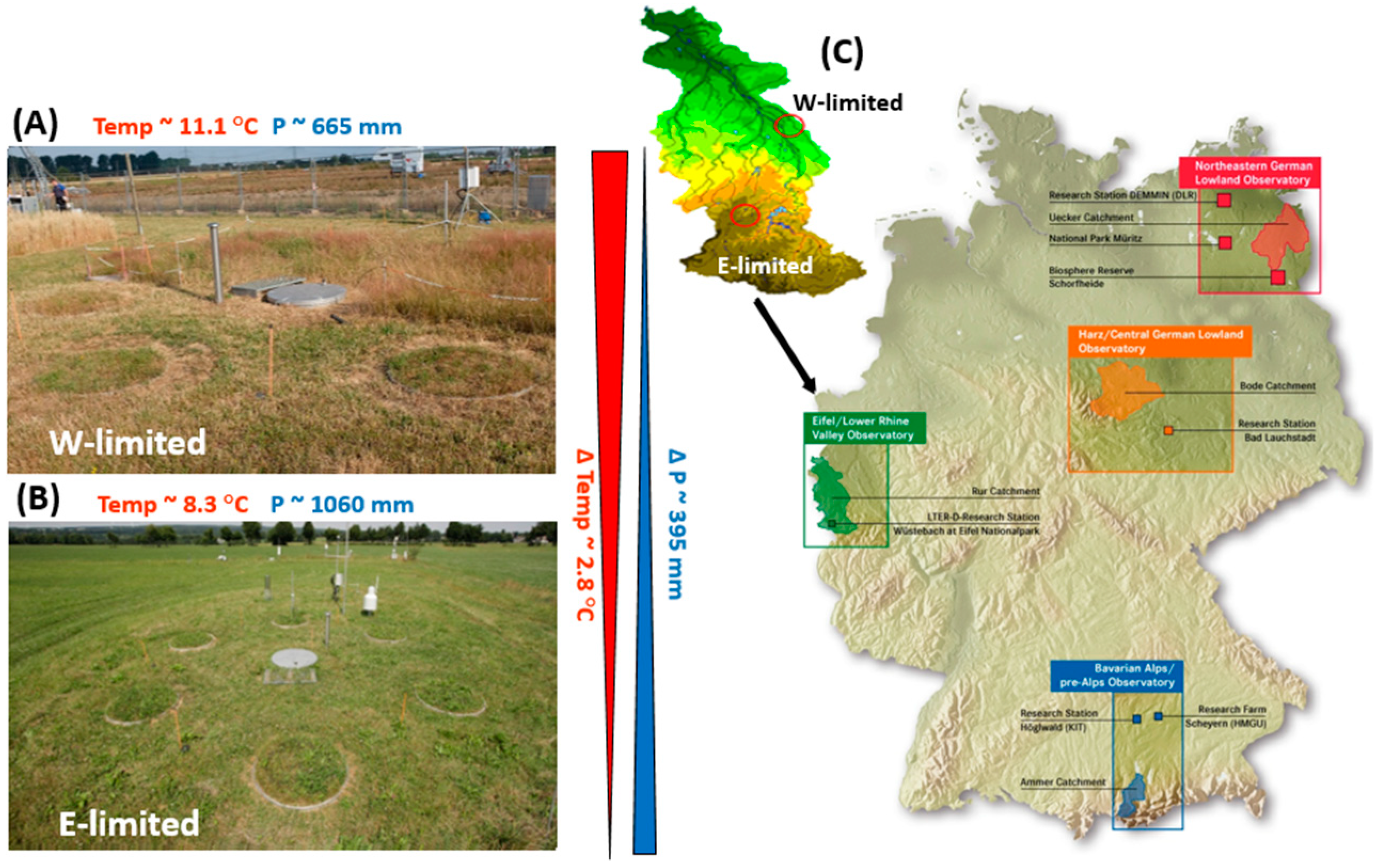
The study sites are part of the Eifel/Lower Rhine Observatory, North Rhine-Westphalia, Germany—of the TERENO SOILCan Lysimeter network [
39]. One site is located near Rollesbroich—50°37′12″ N 6°18′15″ E, 515 m asl, 1.63° slope, a grassland dominated by
Lolium perenne [
42], and the other near Selhausen—50°52′7″ N 6°26′58″ E, 104 m asl, 0.3° slope [
43]. The annual average T
air in Rollesbroich measured for the period 2012–2018 was 8.3 °C, while it was 11.1 °C in Selhausen. Mean annual Prec for the period 2012–2018, as measured with lysimeters, was 1060 mm year
−1 in Rollesbroich and 665 mm year
−1 in Selhausen. A graphical presentation of the TERENO network and the two study sites can be found in
Figure A1 (see
Appendix A).
In 2010, nine lysimeters were filled with undisturbed soil monoliths at Rollesbroich, six of them—see TERENO-portal
www.tereno.net/ddp/, accessed 1 April 2020, lysimeter ID: Ro_Y_011 to Ro_Y_016—were installed in the close vicinity of the monolith excavation site in Rollesbroich, and three—lysimeter ID: Se_Y_021, Se_Y_025, and Se_Y_026—were transferred to the site Selhausen. From these lysimeters, data from 2012 to 2018 was used as explained below. Data from 2011 was not considered because the lysimeters required a period of at least one hydrological year to adapt to the sites’ conditions [
37].
Each lysimeter consists of an outer cylindrical stainless steel shell of 1.50 m height and 1 m² surface area. A silicon porous suction cup rake (SIC40, UMS GmbH, Munich, Germany) at the bottom of the soil monolith lysimeters at approximately 145 cm depth is connected to a bi-directional pump and a seepage water tank. This pump allows for the transport of water into or out of the lysimeter to reduce any difference in matric potentials between the surrounding field soil and the lysimeter soil at 1.4 m depth. Through this pump control, the upward- or downward-directed water flux at the lysimeter bottom boundary is supposed to mimic the real water movement in the surrounding field soil. The lysimeters and the seepage water tanks are equipped with weight cells of high resolution—0.01 kg for the lysimeters, 0.001 kg for the tanks. Aliquots of the seepage water—only downward water flux—are collected in a bottle attached to the water tank. A detailed description of the lysimeters and setup is given in Pütz et al. [
39] and Groh et al. [
40].
Water samples from the tanks, aliquots and deposition sampler were collected every two weeks and analyzed for DIN, NO3−, and NH4+. The DIN was measured via a TOC—total organic carbon—Analyzer (TOC-VCPH, Shimadzu Corporation, Kyoto, Japan) with a total nitrogen measuring unit (TNM-1, Shimadzu Corporation, Kyoto, Japan). The NO3− concentration was determined with an ion chromatography system (Dionex-ICS-4000 and Dionex-AS-23/-24, Thermo Fisher Scientific Inc., Waltham, MA, USA). NH4+ content was measured with a mass spectrometer (Perkin Elmer SCIEX ELAN 6000, PerkinElmer, Inc., Waltham, MA, USA) until 2016 and afterwards via a continuous flow analysis (CFA, Skalar Analytic GmbH, Erkelenz, Deutschland).
The gaseous N
2O emissions were determined by collecting air samples each week for all lysimeter at Selhausen and for three of the six lysimeters at Rollesbroich—Ro_Y_011, Ro_Y_014, Ro_Y_015—during the period from September 2013 until December 2018. Additional measurements were done within three days after fertilizer application. The samples were manually collected with closed static chamber measurements [
39]. Each measurement lasted 32 min, during which four samples of the air in the chamber were taken after 2, 12, 22, and 32 min. The samplings were carried out at around noon. This time of the day was proposed to allow for a good estimation of the mean daily emission rate [
44]. The N
2O concentration in the gas samples from the static chambers was analyzed by gas chromatography (Clarus 580 with ECD, PerkinElmer, Inc., Waltham, MA, USA).
The management of the vegetation on the lysimeters—date for the grass cuts, fertilizer application—follows the extensive management of the surrounding grassland in Rollesbroich—i.e., three to four grass cuts per year, low N-fertilizer input by liquid manure. Only for 2018, the date of the organic fertilizer input was missed, and instead of liquid manure, calcium ammonium nitrate with a N-concentration of 27% (CAN27) was added at the end of the year (see
Table A1 in the
Appendix A). Samples of the mowed grass and applied fertilizer were weighed with a precision scale (EMS 6K0.1 model, KERN & SOHN GmbH, Balingen, Germany, range: 0–6 kg, resolution: 0.1 g) and dried at 60 °C for 24 h and then at 105 °C to evaluate the percentage of dry matter. The dry mater samples were also analyzed with a CHN—carbon hydrogen nitrogen—elemental analyzer (VarioElCube, Elementar Analysensysteme GmbH, Langenselbold, Germany) and a thermal conductivity detector to measure the N-content.
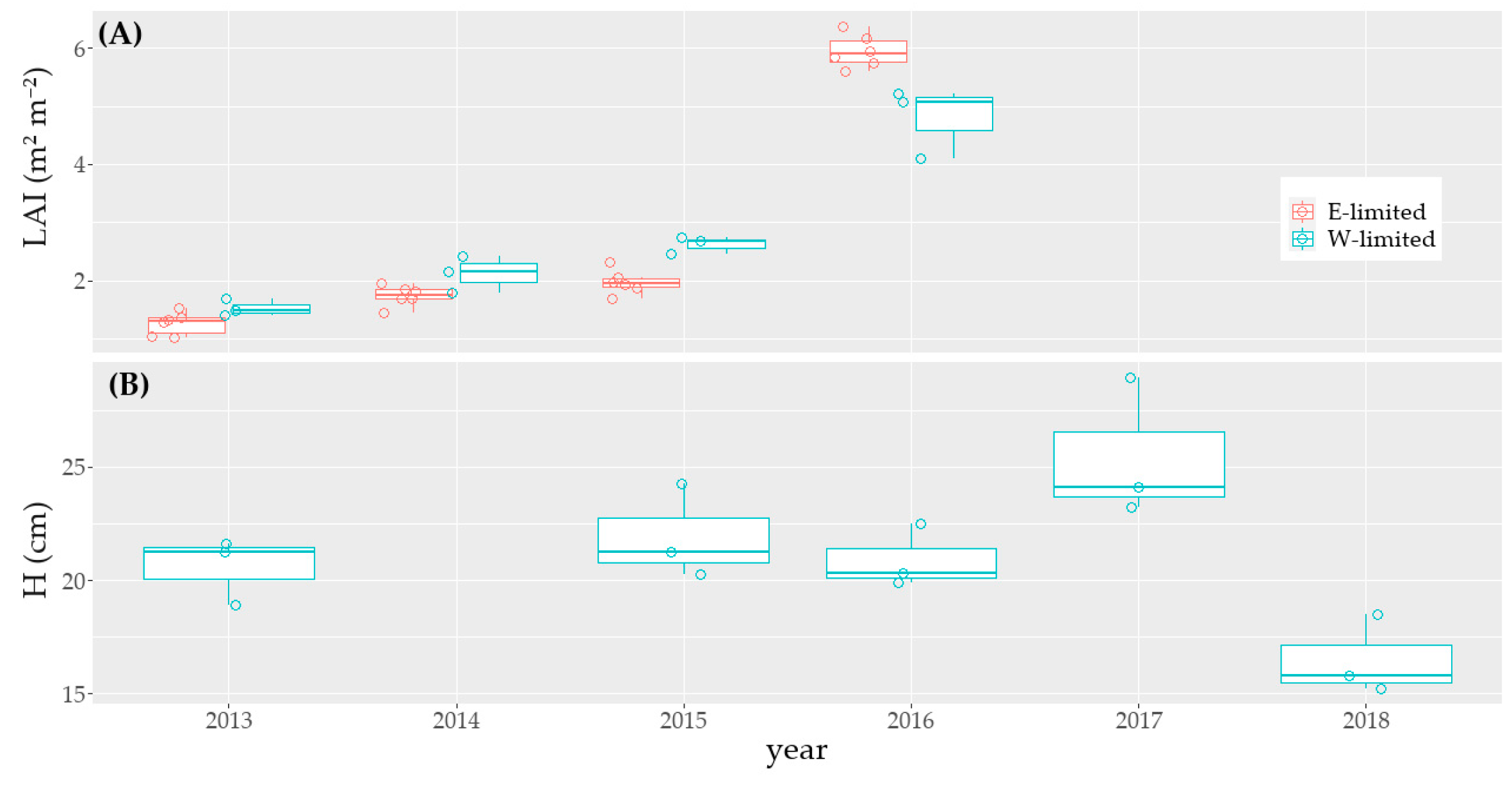
Changes in soil organism dynamics were not monitored as this would involve destructive soil sampling. Plant growth dynamics were monitored via height measurements. A presentation of the gap-filled height data is presented in
Figure A2 (see
Appendix A).
Climatic variables and data gathered from the lysimeter balances and sensors had previously been processed and checked for plausibility [
45,
46]. To reduce the noise of the data, the smoothing filter AWAT—Adaptive Window and Adaptive Threshold filter [
47,
48]—was used. For more information regarding the data preparation, see Pütz et al. [
39] and Rahmati et al. [
37].
For this study, the data qualified as “good” was downloaded from the TERENO online database—
http://teodoor.icg.kfa-juelich.de/ibg3searchportal2/index.jsp, accessed 1 April 2020. When data regarding climatic variables were missing for the reference climatic station of the lysimeters—TERENO-portal, Rollesbroich: Ro_BKY_010; Selhausen: Se_BDK_002— we gap-filled the time-series using data from other stations near the lysimeters from the TERENO-portal and did a step-wise gap-filling: (a) a linear regression was done with each station for each variable; (b) the regression with the highest coefficient of determination (R²) was used first to fill the gaps. The data was processed with R, a programming language and free software environment for statistical computing [
49].
2.2. Water Balance
The water balance at the water- and energy-limited site was described according toPütz et al. [
39]:
with Prec (mm), ETa (mm), Dnet (mm) is the net water flux across the lysimeter bottom, Dr the seepage water (mm), CR the water injected at the bottom of the lysimeter (mm), mimicking the capillary rise of water, and ΔWS the change in soil water storage (mm). Prec also includes water from non-rainfall events like dew or fog formation, which can account annual up to 8% of the total Prec at the sites (see Groh et al. [
50] andBrunke et al. [
51]).
Gaps in the time series’ of daily ETa (or Prec) data were filled in a step-wise procedure based on linear regressions with the ETa (or Prec) of the other lysimeters of the same study site, as explained in detail in Groh et al. [
40]. For filling all remaining gaps of ETa time series, a linear regression between the data from the lysimeter with those of the grass height-adjusted reference evapotranspiration (ETcrop) was used. ETcrop was calculated on a 10 min basis according to Allen et al. [
52] using the full form Penman–Monteith model (see Rahmati et al. [
37]).
To fill gaps in the time series’ of daily grass height, we applied the following method: (a) we calculated the mean of all the available measurements taken at the first day of the growing period (respectively after the grass cut) and used it as default value for the first day of the growing period (respectively after the grass cut) when no data was available; (b) between two cuts during the growing period, we did a linear interpolation between the available measurements; (c) during the growing period, when there were a data gap between the last grass measurement and the day of the grass cut, we did an extrapolation—based on the last available values. Rahmati et al. [
37] did directly an interpolation between the grass height measurements. For this reason, our grass height and ETcrop data differ from theirs.
At the catchment scale, climate–vegetation interactions can be defined according to the Budyko framework [
53]. Briefly, plant transpiration and soil evaporation are dependent on the potential evapotranspiration to water supply ratio (i.e., AI, the aridity index, Equation (3)). Indeed, if AI < 1, evapotranspiration is limited by its demand for energy, whereas for AI > 1, water is the limiting factor for evapotranspiration. The other Budyko factor (EI, the annual evaporation index, defined as the ratio of ETa to water supply, Equation (4)) represents the water partitioning between evapotranspiration and run-off In their study, Rahmati et al. [
37] observed a mean annual AI < 1 for Rollesbroich—thereafter called E-limited—and AI > 1 for Selhausen—thereafter called W-limited. They, however, defined water supply as the annual precipitation. Thus, their analysis did not account for the effects of ground and surface water interaction, which can lead to strongly biased results [
54]. To analyze the climatic conditions at both sites for each year, we therefore used the Budyko framework, as adapted by Condon and Maxwell [
54] (Equation (5)), with ETa and ETcrop (mm), and Prec (mm) for the year n. To capture all available water for the soil for the year n, groundwater contribution to water input was added to the denominators: CR (mm) for the year n and ΔW when the ΔWS of the year n-1 was positive. The Budyko curve was computed using the equation of Fu [
55]. The value of Fu’s parameter (ω) was taken fromRahmati et al. [
37]—ω = 2.6.
2.3. Nitrogen Balance
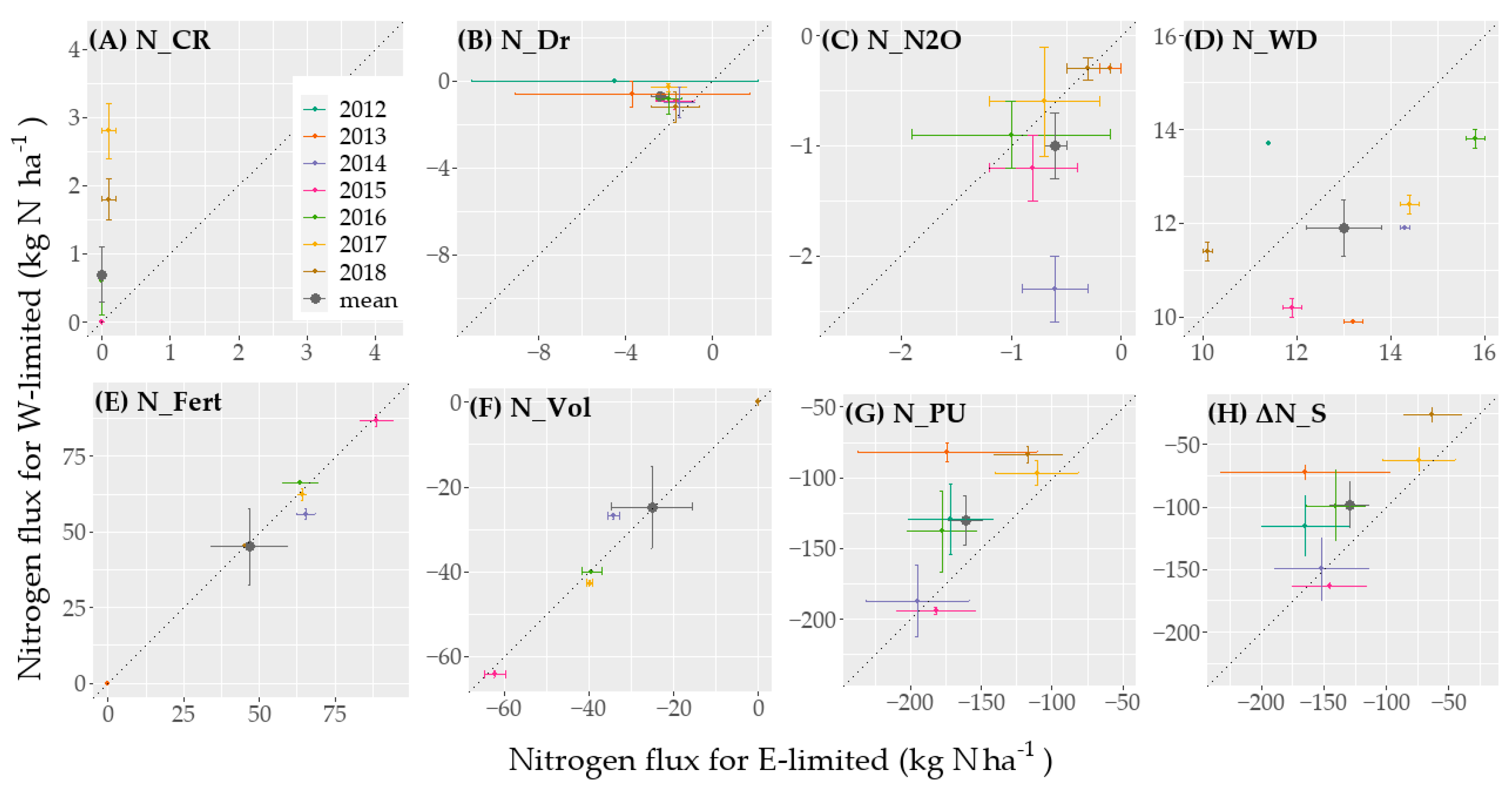
The soil N-balance for the lysimeter soils was described according to Klammler and Fank [
56] by the sum of all measurable input and output fluxes that are balanced by the storage change as
with N_WD the N from the wet deposition (kg ha
−1), N_Fert the fertilizer N (kg ha
−1), and N_Vol the ammonia volatilization of the fertilized N (kg ha
−1), N_PU (kg ha
−1); N_Dr is the leached N in the drainage water (kg ha
−1) determined from the N-concentration in the bottle where seepage water aliquot is collected, N_CR is the N in the solution injected at the bottom of the lysimeter (kg ha
−1) determined from the N-concentration in the lysimeter tank, N_N
2O is the gaseous flux of N
2O (kg ha
−1), and ΔN_S the change in soil N-storage (kg ha
−1). For periods without available N
2O data, the N
2O flux was assumed to be zero because it is mostly negligibly small compared to other N-fluxes in extensively managed grasslands [
22,
30]. The N-fluxes from N_Dr, N_CR, and N_WD data were processed and gap-filled following the method presented by Knauer [
57]. During dry periods, it was not possible to collect enough water to analyze the water samples because the water flow across the lysimeter bottom was small, which led to larger gaps in the concentration time series. There is therefore no observation for 2012 for W-limited. The data time series contain other gaps that correspond to periods with negligible water and nitrogen fluxes.
With the N
2O-concentration data—from the gas-chambers of the W-limited lysimeters and the E-limited lysimeters Ro_Y_011, Ro_Y_014, and Ro_Y_015, the N2O flux rate (f0) was computed according to the Hutchinson–Mosier Regression (HMR) approach [
58] using the HMR package of R [
59]. Either a non-linear HMR or a linear regression was selected according to the criteria described in Deppe [
22]. From f0 (ppm min
−1), the N_N
2O flux rate (N_N
2O-rate, in µg N m
−2 h
−1) was calculated (Equation (8)).
with M_N
2O the molar mass of N
2O (g mol
−1), V_N
2O its molar volume (m³ mol
−1), M_N the molar mass of N (g mol
−1), and the numbers are for unit conversions. Daily N_N
2O emissions were calculated from the hourly N_N
2O emission rates. Then, the N_N
2O emissions between two sampling dates—for intervals inferior to one month—were filled using a linear interpolation by averaging the fluxes of two consecutive sampling dates, multiplied by the number of days in this period. With this technique, we obtained the N_N
2O emission data for 120 days in 2013, 365 days in 2014, 2015, and 2016, 339 days in 2017, and 142 days in 2018.
To obtain an approximation of N_Vol, the equation of Menzi et al. [
60] was used:
with N_Vol in kg N ha
−1, TAN the total ammonia-N content of the fertilizer (g N ha
−1), SD the mean saturation deficit of the air (mbar) and AR the application rate of the fertilizer—the amount of each fertilizer application (kg ha
−1). The environmental variables correspond to the mean value for the day of the fertilizer input. We assumed that TAN made up about 65% of the fertilizer’s N [
61].
For the CAN27, the approach suggested by the COMIFER Association—the French Committee for the Study and Development of sustainable Fertilization—[
62] was used to evaluate the volatilization. Both methods serve in the first instance as a rough estimate rather than a precise method to evaluate N_Vol. We still assumed that this approximation would allow for a more exact calculation of ΔN_S than disregarding N_Vol.
2.4. Statistical Analysis
The shapiro.test and leveneTest functions from the stats [
49] and car [
63] packages in R were used to evaluate the dependent variables’ normal-like distribution and the homogeneity of their variance. A
p-value < 0.01 was considered as significant. When necessary, the variables were transformed to obtain a normal distribution using the BoxCoxTrans function from the caret package [
64].
The parametric Pearson (respectively non-parametric Kendall) correlation coefficient was used to compare the influence of plant aboveground net primary production (ANPP) and plant N-concentration on N_PU (respectively Dr and N-concentration in Dr on N_Dr). The tests were done with the function cor.test from the package stats.
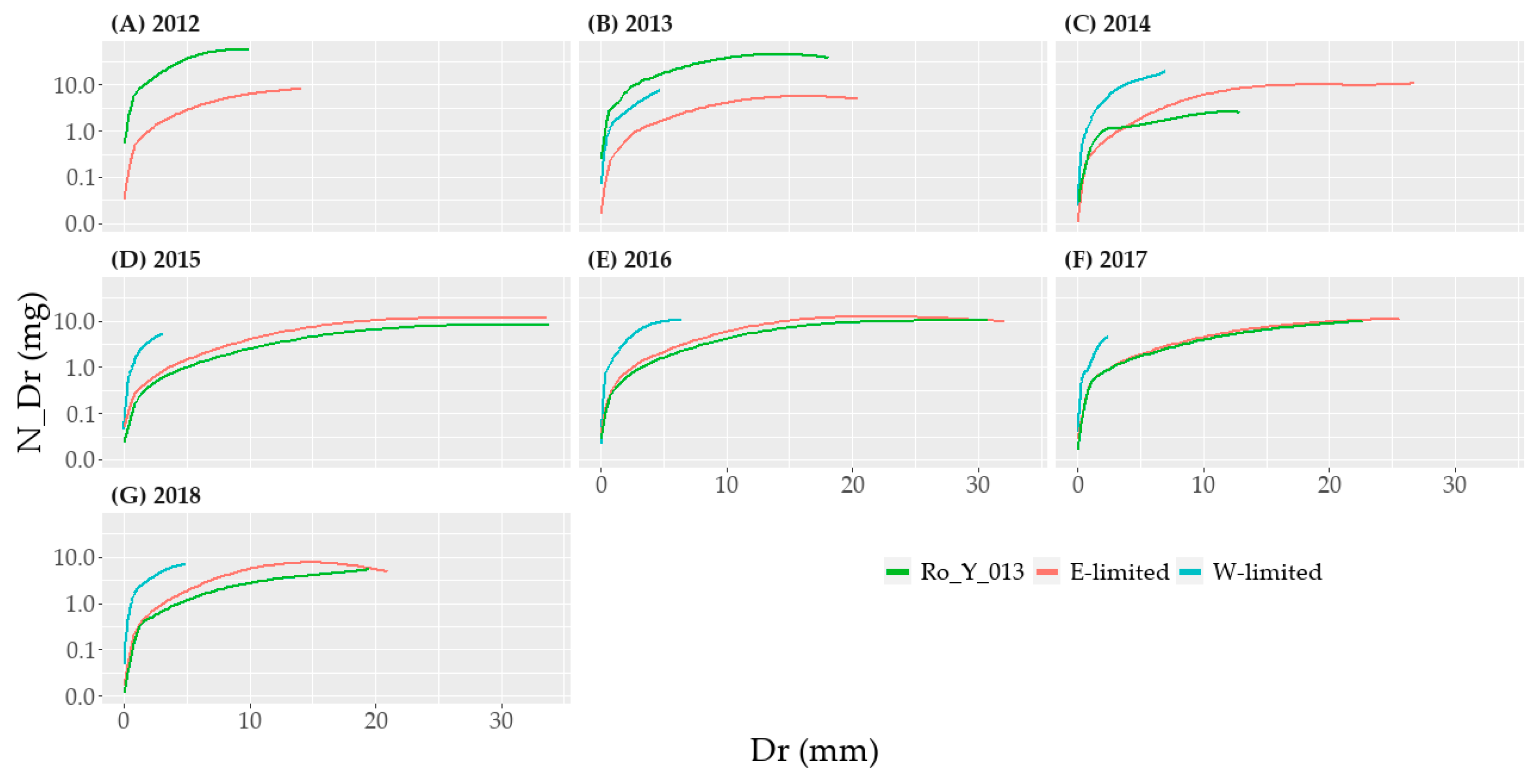
The interpolation of N_Dr according to Dr was done using a Locally Estimated Scatterplot Smoothing (LOESS) with a span of 0.75 using the function geom_smooth from the package ggplot2 of R [
65]. To evaluate the influence of the site factor (W-limited or E-limited) on nitrogen loss, we looked at three elements of the nitrogen balance: N_N
2O-rate, N_Dr, and N_PU per harvest. We also took into account the year factor as control variable as well as a random factor specific for each lysimeter. As the data corresponds to observations from the same sites, the assumption of independence necessary for ordinary least square regression was not met. For this reason, the lmer function from the lmerTest package [
66] was used with the anova function from the car package (Analysis of Variance Table of type III with Satterthwaite’s method) to perform a mixed-effects analysis for repeated-measures.
When analyzing N_PU, the harvest number of the year was also used as a control variable.
When analyzing N_Dr, Dr was also taken into account as control variable. Even once transformed, the N_Dr data did not meet the assumption of normality and homoscedasticity of variance according to the Shapiro and Levene tests. However, the sample size was very important—e.g., 1498 observations for E-limited in 2014. This affected negatively the
p-value calculation of those two tests. We therefore still implemented the parametric mixed-effects analysis, bearing in mind that the results may not be as trustworthy. Indeed, according to the quantile-quantile plot (
Figure A3) the data departed in some subgroups from a normal-like distribution.


 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







