
Phytoplankton Community in the Western South China Sea in Winter and Summer



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sample Collection and Analysis
2.3. Data Analysis
3. Results
3.1. Seasonality in the Environmental Variables
3.2. Phytoplankton Species Composition in the Study Region
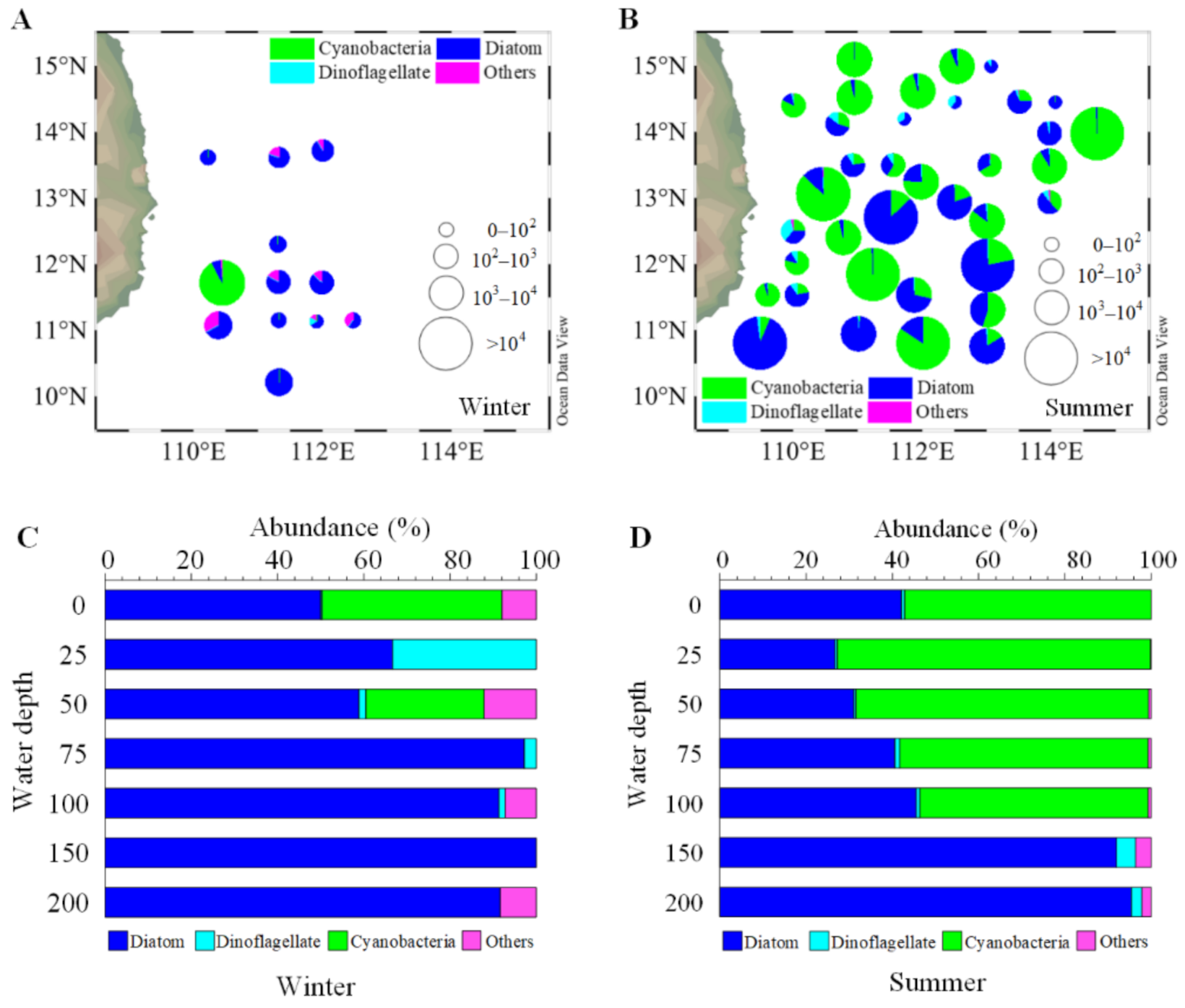
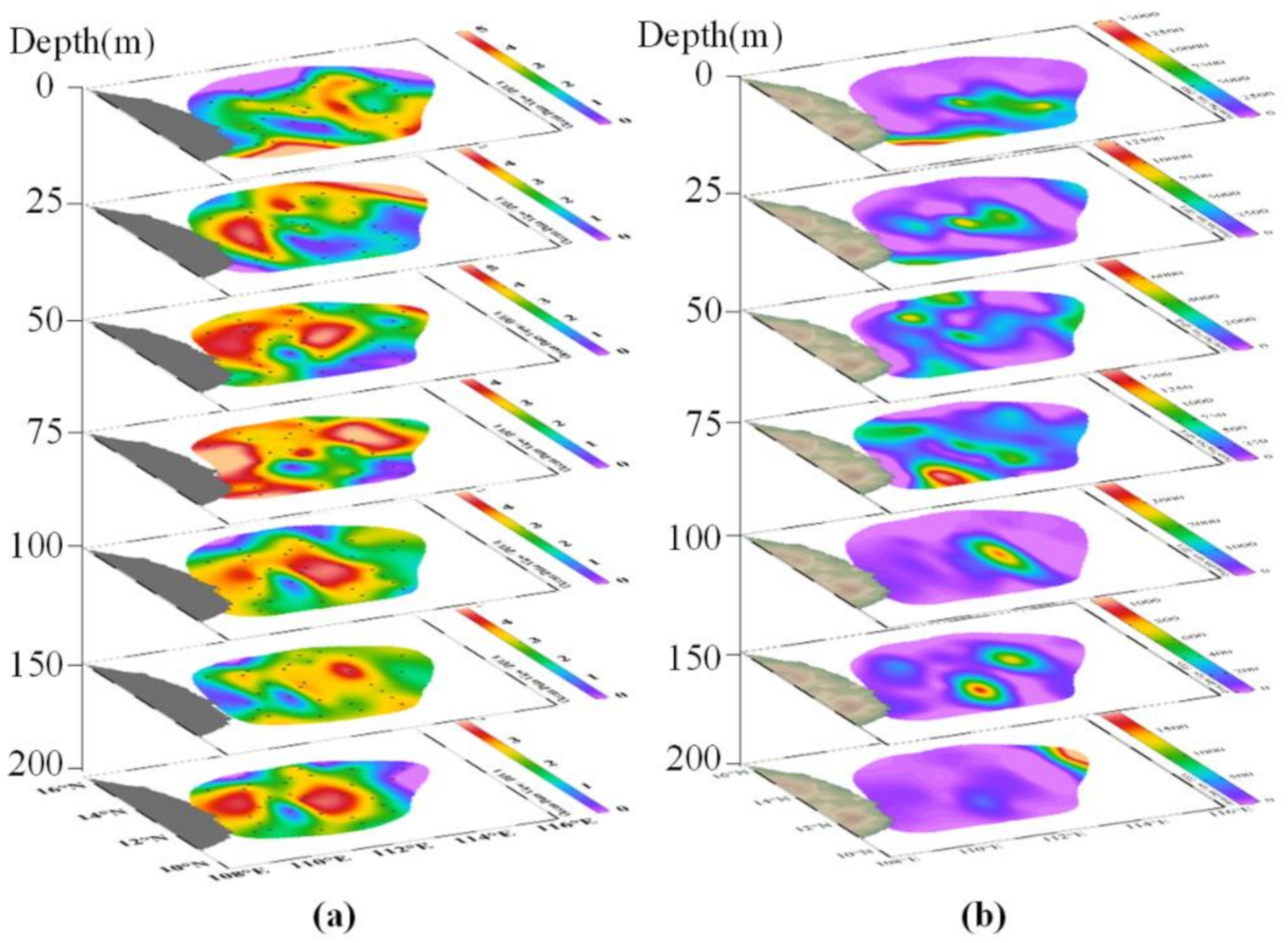
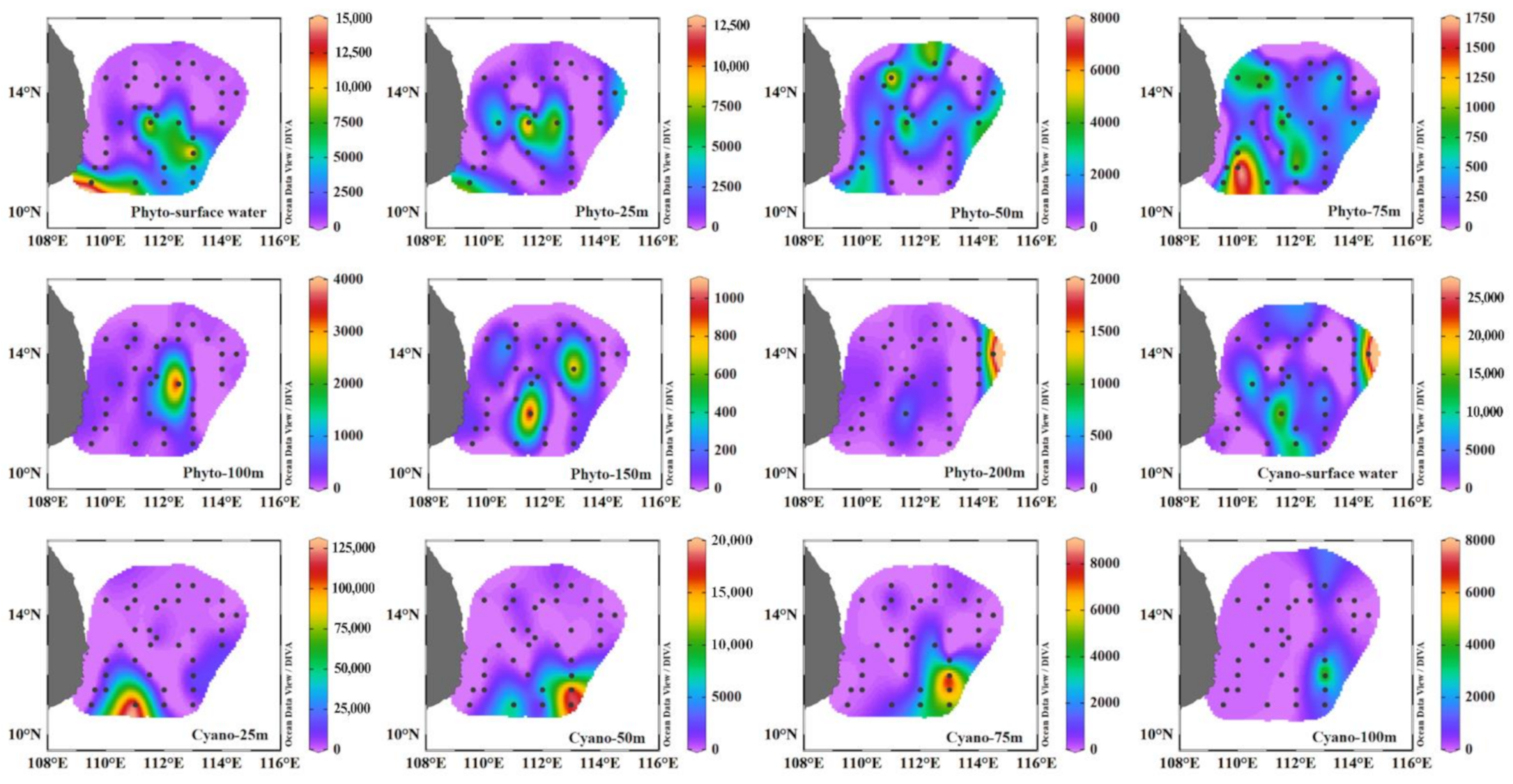
3.3. Seasonal Distribution of Phytoplankton Community
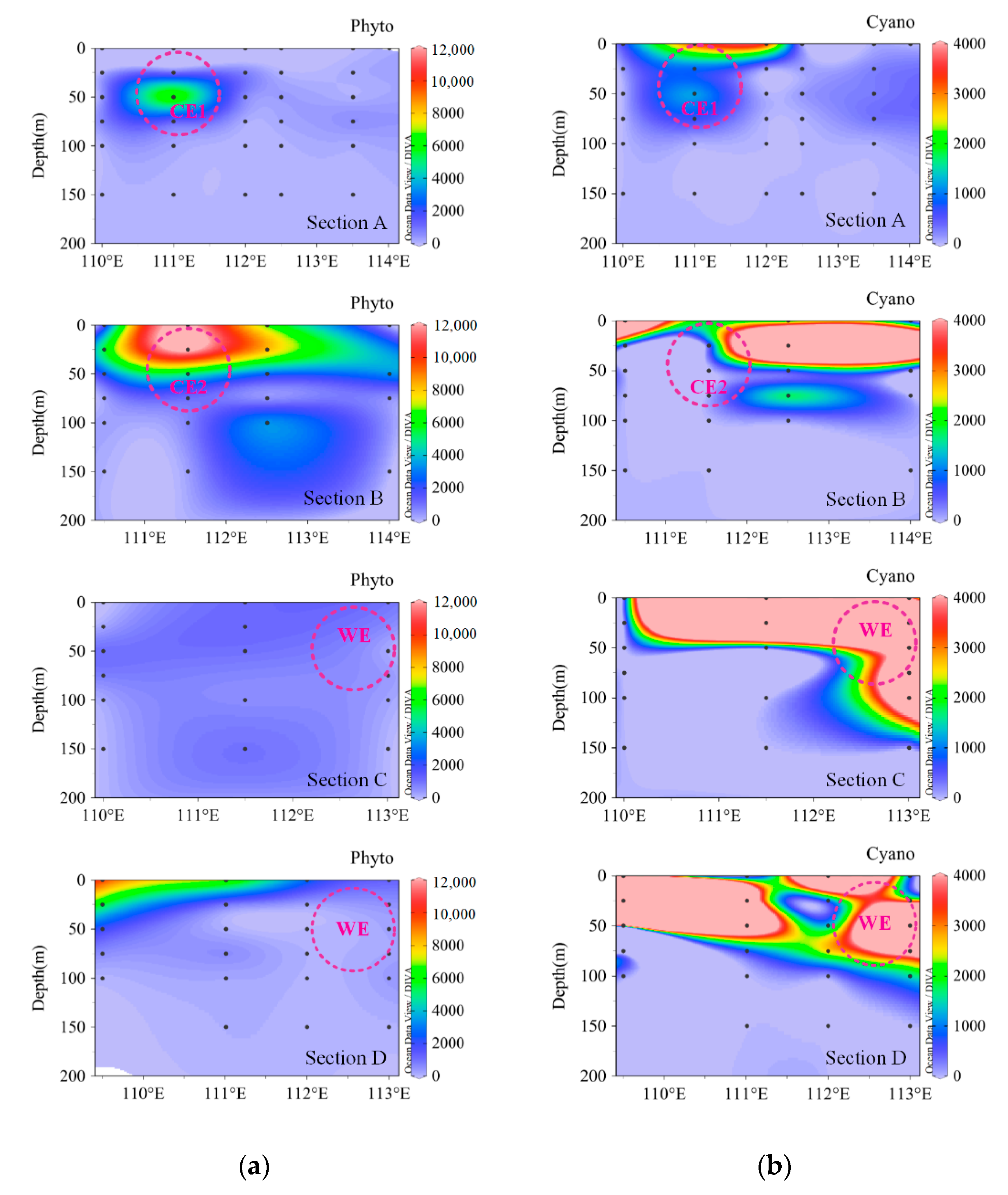
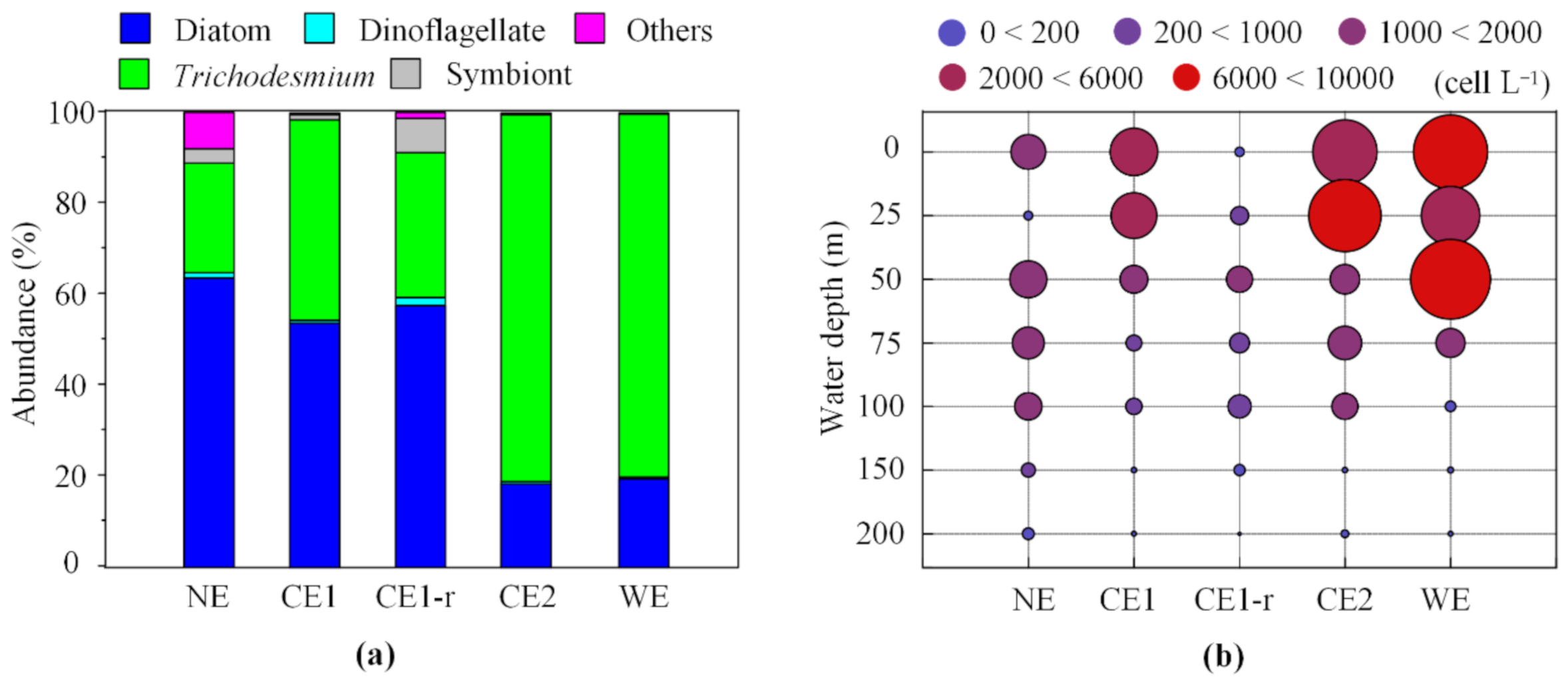
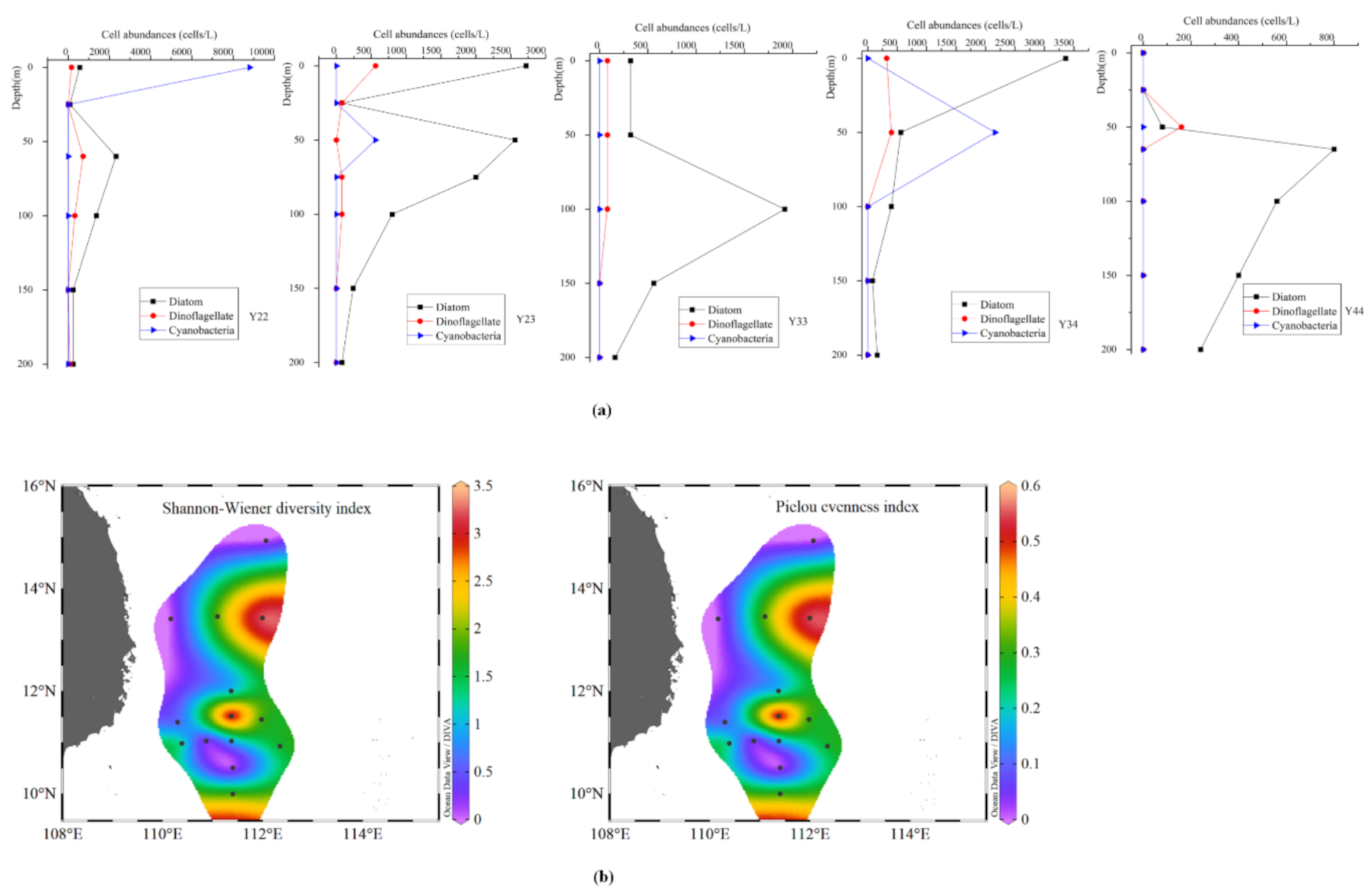
3.4. Vertical Distribution of Phytoplankton Community at Different Eddy Stages
3.5. Diversity of Phytoplankton Community
3.6. Effect of the Environmental Cues on the Phytoplankton Community
4. Discussion
4.1. Influence of Hydrological Processes on the Phytoplankton Community
4.2. Significance of Diazotrophic Cyanobacteria in the Western SCS
4.3. Phytoplankton Thermal Adaptations Inferred from Seasonal Successions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Layer | Temperature (°C) | Salinity | NOx (μmol/L) | PO43− (μmol/L) | SiO32− (μmol/L) |
|---|---|---|---|---|---|---|
| Winter/NE | 0–50 | 27.61 | 33.48 | <0.2 | 0.03 | 2.39 |
| 50–100 | 21.11 | 34.35 | 7.2 | 0.44 | 8.52 | |
| 100–150 | 16.92 | 34.54 | 15.62 | 1.02 | 18.06 | |
| 150–200 | 15.20 | 34.50 | 18.42 | 1.28 | 23.63 | |
| CE1 | 0–50 | 27.55 | 33.77 | 0.50 | 0.05 | 2.70 |
| 50–100 | 22.27 | 34.28 | 5.87 | 0.40 | 7.75 | |
| 100–150 | 17.71 | 34.53 | 14.45 | 0.97 | 15.52 | |
| 150–200 | 13.64 | 34.51 | 15.67 | 1.33 | 24.67 | |
| CE1-r | 0–50 | 27.56 | 33.80 | <0.2 | 0.05 | 2.20 |
| 50–100 | 22.38 | 34.27 | 4.55 | 0.33 | 6.28 | |
| 100–150 | 18.94 | 34.50 | 11.05 | 0.83 | 13.32 | |
| 150–200 | 17.38 | 34.57 | 15.60 | 1.24 | 22.71 | |
| CE2 | 0–50 | 28.36 | 33.37 | <0.2 | 0.03 | 2.28 |
| 50–100 | 22.36 | 34.29 | 6.44 | 0.40 | 7.34 | |
| 100–150 | 17.31 | 34.53 | 14.39 | 0.94 | 14.16 | |
| 150–200 | - | - | 18.80 | 1.27 | 22.19 | |
| WE | 0–50 | 29.18 | 32.99 | <0.2 | 0.05 | 2.25 |
| 50–100 | 24.92 | 33.93 | 4.55 | 0.17 | 3.96 | |
| 100–150 | 17.74 | 34.55 | 14.72 | 0.86 | 13.47 | |
| 150–200 | - | - | 17.50 | 1.11 | 18.16 |
| Group | Winter | Summer |
|---|---|---|
| Diatom Range | 80–3360 | 5–13,482 |
| Average | 670 | 827 |
| Total | 26,800 | 190,045 |
| Proportion | 63.57% | 30.18% |
| Dinoflagellate Range | 80–160 | 2–245 |
| Average | 12 | 16 |
| Total | 480 | 3585 |
| Proportion | 1.14% | 0.57% |
| Cyanobacteria Range | 560–8800 | 16–123,150 |
| Average | 288 | 1891 |
| Total | 11,520 | 434,912 |
| Proportion | 27.32% | 69.07% |
| Others Range | 80–560 | 3–60 |
| Average | 84 | 5 |
| Total | 3360 | 1166 |
| Proportion | 8.00% | 0.19% |
| Phytoplankton Range | 80–9520 | 20–128,820 |
| Average | 1054 | 2738 |
| Total | 42,160 | 629,708 |
| Season | Depth (m) | Diatom | Dinoflagellate | Cyanobacteria | Others | Total | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Average (cell L−1) | Proportion | Average (cell L−1) | Proportion | Average (cell L−1) | Proportion | Average (cell L−1) | Proportion | Average (cell L−1) | ||
| Winter | 0 | 704 | 50.00% | 5.33 | 0.38% | 586.67 | 41.67% | 112 | 7.95% | 1408 |
| 25 | 53.33 | 66.67% | 26.67 | 33.33% | 0 | 0 | 0 | 0 | 80 | |
| 50 | 880 | 51.16% | 0 | 0 | 680 | 39.53% | 160 | 9.30% | 1720 | |
| 75 | 1706.67 | 86.49% | 80 | 4.05% | 0 | 0 | 186.67 | 9.46% | 1973.33 | |
| 100 | 1008 | 91.30% | 16 | 1.45% | 0 | 0 | 80 | 7.25% | 1104 | |
| 150 | 304 | 100% | 0 | 0 | 0 | 0 | 0 | 0 | 304 | |
| 200 | 176 | 91.67% | 0 | 0 | 0 | 0 | 16 | 8.33% | 192 | |
| Summer | 0 | 1978.19 | 38.02% | 39.14 | 0.75% | 3182.31 | 61.16% | 3.04 | 0.06% | 5202.69 |
| 25 | 1372.29 | 17.06% | 32.58 | 0.40% | 6633.21 | 82.45% | 6.50 | 0.08% | 8044.56 | |
| 50 | 1295.67 | 41.45% | 13.52 | 0.43% | 1803.94 | 57.71% | 13.10 | 0.42% | 3126.23 | |
| 75 | 414.11 | 31.61% | 10.49 | 0.80% | 880.32 | 67.20% | 5.42 | 0.41% | 1310.35 | |
| 100 | 264.39 | 43.48% | 4.90 | 0.81% | 335.40 | 55.17% | 3.68 | 0.61% | 608.39 | |
| 150 | 125.54 | 95.84% | 3.17 | 2.42% | 0 | 0 | 2.08 | 1.58% | 130.79 | |
| 200 | 107.60 | 97.82% | 1.23 | 1.12% | 0 | 0 | 1.09 | 0.99% | 109.92 | |
| Group | NE | CE1 | CE1-r | CE2 | WE | |
|---|---|---|---|---|---|---|
| Abundance (cell L−1) | Diatoms | 26,800 | 104,334 | 3654 | 53,827 | 28,231 |
| Dinoflagellate | 480 | 1258 | 116 | 1579 | 627 | |
| Cyanobacteria | 11,520 | 87,816 | 2507 | 236,753 | 116,098 | |
| Trichodesmium | 10,160 | 85,635 | 2025 | 236,341 | 115,810 | |
| R. intracellularis | 1360 | 2181 | 482 | 411 | 288 | |
| Other groups | 3360 | 621 | 82 | 406 | 56 | |
| Total | 42,160 | 194,029 | 6359 | 292,564 | 145,012 | |
| Proportion | Diatoms | 63.57% | 53.77% | 57.46% | 18.40% | 19.47% |
| Dinoflagellate | 1.14% | 0.65% | 1.82% | 0.54% | 0.43% | |
| Cyanobacteria | 27.32% | 45.26% | 39.42% | 80.92% | 80.06% | |
| Trichodesmium | 24.10% | 44.14% | 31.84% | 80.78% | 79.86% | |
| R. intracellularis | 3.23% | 1.12% | 7.58% | 0.14% | 0.20% | |
| Other groups | 7.97% | 0.32% | 1.29% | 0.14% | 0.04% | |
| Abundance ratio | Diatoms/Dinoflagellate | 55.83 | 82.94 | 31.50 | 34.10 | 44.99 |
| Diatoms/Cyanobacteris | 2.33 | 1.19 | 1.46 | 0.23 | 0.24 | |
| Winter | CE1 | CE1-r | CE2 | |
|---|---|---|---|---|
| Summer | a1, b1, c3, d1 | |||
| CE1-r | a2, b3, c3, d3 | |||
| CE2 | a2, b1, c3, d1 | a2, b2, c3, d1 | ||
| WE | a2, b2, c3, d1 | a2, b2, c3, d3 | a1, b2, c3, d1 |




References
- Voss, M.; Bombar, D.; Loick, N.; Dippner, J.W. Riverine influence on nitrogen fixation in the upwelling region off Vietnam, South China Sea. Geophys. Res. Lett. 2006, 33, L07604. [Google Scholar] [CrossRef]
- Cai, W.-J.; Dai, M.; Wang, Y.; Zhai, W.; Huang, T.; Chen, S.; Zhang, F.; Chen, Z.; Wang, Z. The biogeochemistry of inorganic carbon and nutrients in the Pearl River estuary and the adjacent Northern South China Sea. Cont. Shelf Res. 2004, 24, 1301–1319. [Google Scholar] [CrossRef]
- Guo, M.; Chai, F.; Xiu, P.; Li, S.; Rao, S. Impacts of mesoscale eddies in the South China Sea on biogeochemical cycles. Ocean Dyn. 2015, 65, 1335–1352. [Google Scholar] [CrossRef]
- Yang, H.; Liu, Q.; Liu, Z.; Wang, D.; Liu, X. A general circulation model study of the dynamics of the upper ocean circulation of the South China Sea. J. Geophys. Res. 2002, 107. [Google Scholar] [CrossRef]
- Cheng, Z.D. Fluxes of seawater and some dissolved chemicals in the Bashi Strait. Chin. J. Oceanol. Limnol. 1995, 13, 240–246. [Google Scholar]
- Dippner, J.W.; Nguyen, K.V.; Hein, H.; Ohde, T.; Loick, N. Monsoon-induced upwelling off the Vietnamese coast. Ocean Dyn. 2007, 57, 46–62. [Google Scholar] [CrossRef]
- Gong, G.-C.; Liu, K.K.; Liu, C.-T.; Pai, S.-C. The chemical hydrography of the South China Sea west of Luzon and a comparison with the west Philippine Sea. Terr. Atmos. Ocean. Sci. 1992, 3, 587–602. [Google Scholar] [CrossRef]
- Chen, Y.-L.L.; Chen, H.-Y.; Karl, D.M.; Takahashi, M. Nitrogen modulates phytoplankton growth in spring in the South China Sea. Cont. Shelf Res. 2004, 24, 527–541. [Google Scholar] [CrossRef]
- Chen, Y.-L.L.; Chen, H.-Y.; Lin, I.-I.; Lee, M.-A.; Chang, J. Effects of cold eddy on phytoplankton production and assemblages in Luzon strait bordering the South China Sea. J. Oceanogr. 2007, 63, 671–683. [Google Scholar] [CrossRef]
- Liu, K.-K.; Chao, S.-Y.; Shaw, P.-T.; Gong, G.-C.; Chen, C.-C.; Tang, T. Monsoon-forced chlorophyll distribution and primary production in the South China Sea: Observational and a numerical study. Deep Sea Res. Part I Oceanogr. Res. Pap. 2002, 49, 1387–1412. [Google Scholar] [CrossRef]
- Loick-Wilde, N.; Bombar, D.; Doan, H.N.; Nguyen, L.N.; Nguyen-Thi, A.M.; Voss, M.; Dippner, J.W. Microplankton biomass and diversity in the Vietnamese upwelling area during SW monsoon under normal conditions and after an ENSO event. Prog. Oceanogr. 2017, 153, 1–15. [Google Scholar] [CrossRef]
- Sun, J.; Liu, D.Y. Net-phytoplankton community of the Bohai Sea in the autumn of 2000. Acta Oceanol. Sin. 2005, 27, 124–132. [Google Scholar]
- Guo, S.; Feng, Y.; Wang, L.; Dai, M.; Liu, Z.; Bai, Y.; Jun, S. Seasonal variation in the phytoplankton community of a continental-shelf sea: The East China Sea. Mar. Ecol. Prog. Ser. 2014, 516, 103–126. [Google Scholar] [CrossRef] [Green Version]
- Xue, B.; Sun, J.; Li, T.T. Phytoplankton community structure of northern South China Sea in summer of 2014. Acta Oceanol. Sin. 2016, 38, 54–65. [Google Scholar]
- Gan, J.; Li, H.; Curchitser, E.N.; Haidvogel, D.B. Modeling South China Sea circulation: Response to seasonal forcing regimes. J. Geophys. Res. Ocean. 2006, 111. [Google Scholar] [CrossRef]
- Wang, G.; Chen, D.; Su, J. Generation and life cycle of the dipole in the South China Sea summer circulation. J. Geophys. Res. Ocean. 2006, 111. [Google Scholar] [CrossRef] [Green Version]
- Ning, X.; Chai, F.; Xue, H.; Cai, Y.; Liu, C.; Shi, J. Physical-biological oceanographic coupling influencing phytoplankton and primary production in the South China Sea. J. Geophys. Res. 2004, 109. [Google Scholar] [CrossRef] [Green Version]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahadevan, A. The Impact of submesoscale physics on primary productivity of plankton. Ann. Rev. Mar. Sci. 2016, 8, 161–184. [Google Scholar] [CrossRef] [Green Version]
- Zhong, C.; Xiao, W.P.; Huang, B.Q. The response of phytoplankton to mesoscale eddies in western South China Sea. Adv. Mar. Sci. 2013, 31, 213–220. [Google Scholar]
- Tang, D.; Kawamura, H.; Van Dien, T.; Lee, M. Offshore phytoplankton biomass increase and its oceanographic causes in the South China Sea. Mar. Ecol. Prog. Ser. 2004, 268, 31–41. [Google Scholar] [CrossRef]
- Tang, D.L.; Kawamura, H.; Doan-Nhu, H.; Takahashi, W. Remote sensing oceanography of a harmful algal bloom off the coast of southeastern Vietnam. J. Geophys. Res. 2004, 109. [Google Scholar] [CrossRef]
- Wang, J.J.; Tang, D.L. Phytoplankton patchiness during spring intermonsoon in western coast of South China Sea. Deep Sea Res. II Top. Stud. Oceanogr. 2014, 101, 120–128. [Google Scholar] [CrossRef]
- Liang, Y.; Yuan, D.; Li, Q.; Lin, Q. Flow injection analysis of nanomolar level orthophosphate in seawater with solid phase enrichment and colorimetric detection. Mar. Chem. 2007, 103, 122–130. [Google Scholar] [CrossRef]
- Wang, L.; Huang, B.; Chiang, K.-P.; Liu, X.; Chen, B.; Xie, Y.; Hu, J.Y.; Dai, M.H. Physical-biological coupling in the western South China Sea: The response of phytoplankton community to a mesoscale cyclonic eddy. PLoS ONE 2016, 11, e0153735. [Google Scholar] [CrossRef] [PubMed]
- Ke, Z.; Tan, Y.; Huang, L.; Zhang, J.; Lian, S. Relationship between phytoplankton composition and environmental factors in the surface waters of southern South China Sea in early summer of 2009. Acta Oceanol. Sin. 2012, 31, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.; Tang, D.; Luo, X. Phytoplankton size structure in the western South China Sea under the influence of a ‘jet-eddy system’. J. Mar. Syst. 2018, 187, 82–95. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Barber, R.T.; Smetacek, V. Biogeochemical controls and feedbacks on ocean primary production. Science 1998, 281, 200–206. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Gan, J.; Sun, Z.; Zhu, J.; Dai, M. Observed three-dimensional structure of a cold eddy in the southwestern South China Sea. J. Geophys. Res. 2011, 116. [Google Scholar] [CrossRef]
- Fang, W.D.; Fang, G.H.; Shi, P.; Huang, Q.Z.; Xie, Q. Seasonal structures of upper layer circulation in the southern South China Sea from in situ observations. J. Geophys. Res. 2002, 107, 1–12. [Google Scholar] [CrossRef]
- Shaw, P.T.; Chao, S.Y. Surface circulation in the South China Sea. Deep Sea Res. Part I Oceanogr. Res. Pap. 1994, 41, 1663–1683. [Google Scholar] [CrossRef]
- Yuan, Y.C.; Liu, Y.G.; Liao, G.H.; Lou, R.Y.; Su, J.L.; Wang, K.S. Calculation of circulation in the South China Sea during summer of 2000 by the modified inverse method. Acta Oceanol. Sin. 2005, 24, 14–30. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Hu, J.; Liu, Z.; Belkin, I.M.; Sun, Z.; Zhu, J. A peculiar lens-shaped structure observed in the South China Sea. Sci. Rep. 2017, 7, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.P. Nutrient Dynamics Associated with Mesoscale Eddies in the Western South China Sea. Master’s Dissertation, Xiamen University, Xiamen, China, 2009. [Google Scholar]
- Liu, Q.; Kaneko, A.; Su, J.L. Recent progress in studies of the South China Sea circulation. J. Oceanogr. 2008, 64, 753–762. [Google Scholar] [CrossRef]
- Sun, J.; Liu, D.; Qian, S. A quantative research and analysis method for marine phytoplankton: An introduction to utermöhl method and its modification. J. Oceanogr. Huanghai Bohai Seas 2002, 20, 105–112. [Google Scholar]
- Sun, J.; Liu, D.Y.; Feng, S.Z. Preliminary study on marine phytoplankton sampling and analysis strategy for ecosystem dynamic research in coastal waters. Oceanol. Limnol. Sin. 2003, 34, 224–232. [Google Scholar]
- Sun, J.; Liu, D.Y. The preliminary notion on nomenclature of common phytoplankton in China seas waters. Oceanol. Limnol. Sin. 2002, 33, 271–286. [Google Scholar]
- Dai, M.; Wang, L.; Guo, X.; Zhai, W.; Li, Q.; He, B.; Kao, S.-J. Nitrification and inorganic nitrogen distribution in a large perturbed river/estuarine system: The Pearl River estuary, China. Biogeosciences 2008, 5, 1227–1244. [Google Scholar] [CrossRef] [Green Version]
- Pai, S.-C.; Yang, C.-C.; Riley, J.P. Effects of acidity and molybdate concentration on the kinetics of the formation of the phosphoantimonylmolybdenum blue complex. Anal. Chim. Acta 1990, 229, 115–120. [Google Scholar]
- Ma, J.; Yuan, D.; Liang, Y.; Dai, M.H. A modified analytical method for the shipboard determination of nanomolar concentrations of orthophosphate in seawater. J. Oceanogr. 2008, 64, 443–449. [Google Scholar] [CrossRef]
- Schlitzer, R. Ocean Data View. 2018. Available online: https://odv.awi.de (accessed on 2 March 2018).
- Sun, J.; Liu, D.Y. The application of diversity indices in marine phytoplankton studies. Acta Oceanol. Sin. 2004, 26, 62–75. [Google Scholar]
- RCRTeam. R: A Language and Environment for Statistical Computing, Version 3.6.1. Vienna, Austria. 2019. Available online: http://www.R-project.org/ (accessed on 5 July 2019).
- Zhu, G.H.; Ning, X.R.; Cai, Y.M.; Liu, Z.L.; Liu, C.G. Studies on species composition and abundance distribution of phytoplankton in the South China Sea. Haiyang Xuebao 2003, 25, 8–23. (In Chinese) [Google Scholar]
- Ke, Z.X.; Huang, L.M.; Tan, Y.H.; Yin, J.Q. Species composition and abundance of phytoplankton in the northern South China Sea in summer 2007. J. Trop. Oceanogr. 2011, 30, 131–143. [Google Scholar]
- Li, X.; Sun, J.; Tian, W.; Wang, M. Phytoplankton community in the northern area of South China Sea in summer of 2009. Mar. Sci. 2012, 36, 33–39. (In Chinese) [Google Scholar]
- Ma, W.; Sun, J. Characteristics of phytoplankton community in the northern South China Sea in summer and winter. Acta Ecol. Sin. 2014, 34, 621–632. (In Chinese) [Google Scholar]
- Wang, X.; Wei, Y.; Wu, C.; Guo, C.; Sun, J. The profound influence of Kuroshio intrusion on microphytoplankton community in the northeastern South China Sea. Acta Oceanol. Sin. 2020, 39, 79–87. [Google Scholar] [CrossRef]
- Zhong, Q.; Xue, B.; Noman, M.A.; Wei, Y.; Sun, J. Effect of river plume on phytoplankton community structure in Zhujiang River estuary. J. Oceanol. Limnol. 2020, 39, 550–565. [Google Scholar] [CrossRef]
- Chen, G.; Xiu, P.; Chai, F. Physical and biological controls on the summer chlorophyll bloom to the east of Vietnam. J. Oceanogr. 2014, 70, 323–328. [Google Scholar] [CrossRef]
- Martin, A.P.; Pondaven, P. On estimates for the vertical nitrate flux due to eddy pumping. J. Geophys. Res. Ocean. 2003, 108, 3359. [Google Scholar] [CrossRef]
- McGillicuddy, D.J.; Anderson, L.A.; Bates, N.R.; Bibby, T.; Buesseler, K.O.; Carlson, C.; Carlson, C.; Davis, C.S.; Ewart, C.; Falkowski, P.G.; et al. Eddy/wind interactions stimulate extraordinary mid-ocean plankton blooms. Science 2007, 316, 1021–1026. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.P.; Nuncio, M.; Ramaiah, N.; Sardesai, S.; Narvekar, J.; Fernandes, V.; Paul, J.T. Eddy-mediated biological productivity in the Bay of Bengal during fall and spring intermonsoons. Deep Sea Res. Part I Oceanogr. Res. Pap. 2007, 54, 1619–1640. [Google Scholar] [CrossRef] [Green Version]
- Edwards, K.F.; Litchman, E.; Klausmeier, C.A. Functional traits explain phytoplankton community structure and seasonal dynamics in a marine ecosystem. Ecol. Lett. 2013, 16, 56–63. [Google Scholar] [CrossRef]
- Boyd, P.W.; Hutchins, D.A. Understanding the responses of ocean biota to a complex matrix of cumulative anthropogenic change. Mar. Ecol. Prog. Ser. 2012, 470, 125–135. [Google Scholar] [CrossRef] [Green Version]
- Edwards, M.; Richardson, A.J. Impact of climate change on marine pelagic phenology and trophic mismatch. Nature 2004, 430, 881–884. [Google Scholar] [CrossRef]
- Zehr, J.P.; Capone, D.G. Changing perspectives in marine nitrogen fixation. Science 2020, 368. [Google Scholar] [CrossRef]
- Xiao, W.; Wang, L.; Laws, E.; Xie, Y.; Chen, J.; Liu, X.; Chen, B.; Huang, B. Realized niches explain spatial gradients in seasonal abundance of phytoplankton groups in the South China Sea. Prog. Oceanogr. 2018. [Google Scholar] [CrossRef]
- Chen, C.-T.A.; Wang, S.-L.; Wang, B.-J.; Pai, S.-C. Nutrient budgets for the South China Sea basin. Mar. Chem. 2001, 75, 281–300. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, Z.; Sun, J.; Jiao, N. Diversity and distribution of diazotrophic communities in the South China Sea deep basin with mesoscale cyclonic eddy perturbations. FEMS Microbiol. Ecol. 2011, 78, 417–427. [Google Scholar] [CrossRef] [Green Version]
- Moisander, P.H.; Beinart, R.A.; Voss, M.; Zehr, J.P. Diversity and abundance of diazotrophic microorganisms in the South China Sea during intermonsoon. ISME J. 2008, 2, 954–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnet, S.; Berthelot, H.; Turk-Kubo, K.; Cornet-Barthaux, V.; Fawcett, S.; Berman-Frank, I.; Barani, A.; Grégori, G.; Dekaezemacker, J.; Benavides, M.; et al. Diazotroph derived nitrogen supports diatom growth in the south West Pacific: A quantitative study using nanosims. Limnol. Oceanogr. 2016, 61, 1549–1562. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, E.J.; Montoya, J.P.; Burns, J.; Mulholland, M.; Subramaniam, A.; Capone, D.G. Extensive bloom of N2-fixing diatom/cyanobacterial association in the tropical Atlantic Ocean. Mar. Ecol. Prog. Ser. 1999, 185, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Foster, R.A.; Zehr, J.P. Characterization of diatom-cyanobacteria symbioses on the basis of nifH, hetR, and 16S rRNA sequences. Environ. Microbiol. 2006, 8, 1913–1925. [Google Scholar] [CrossRef]
- Foster, R.A.; Subramaniam, A.; Mahaffey, C.; Carpenter, E.J.; Capone, D.G.; Zehr, J.P. Influence of the Amazon River plume on distributions of free-living and symbiotic cyanobacteria in the western tropical North Atlantic Ocean. Limnol. Oceanogr. 2007, 52, 517–532. [Google Scholar] [CrossRef] [Green Version]
- Foster, R.A.; Goebel, N.L.; Zehr, J.P. Isolation of Calothrix Rhizosoleniae (Cyanobacteria) strain SC01 from Chaetoceros (Bacillariophyta) spp. diatoms of the Subtropical North Pacific Ocean. J. Phycol. 2010, 46, 1028–1037. [Google Scholar] [CrossRef]
- Foster, R.A.; Kuypers, M.M.M.; Vagner, T.; Paerl, R.W.; Musat, N.; Zehr, J.P. Nitrogen fixation and transfer in open ocean diatom-cyanobacterial symbioses. ISME J. 2011, 5, 1484–1493. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Trenberth, K.E.; Fasullo, J.; Boyer, T.; Abraham, J.; Zhu, J. Improved estimates of ocean heat content from 1960 to 2015. Sci. Adv. 2017, 3, e1601545. [Google Scholar] [CrossRef] [Green Version]
- Bopp, L.; Monfray, P.; Aumont, O.; Dufresne, J.-L.; Le Treut, H.; Madec, G.; Terray, L.; Orr, J.C. Potential impact of climate change on marine export production. Glob. Biogeochem. Cycles 2001, 15, 81–99. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, S.E.; Mather, R.L.; Wolff, G.A.; Williams, R.G.; Landolfi, A.; Sanders, R.; Woodward, E.M.S. How widespread and important is N2 fixation in the North Atlantic Ocean? Glob. Biogeochem. Cycles 2007, 21. [Google Scholar] [CrossRef] [Green Version]
- Huertas, I.E.; Rouco, M.; Lopez-Rodas, V.; Costas, E. Warming will affect phytoplankton differently: Evidence through a mechanistic approach. Proc. R. Soc. B Biol. Sci. 2011, 278, 3534–3543. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.K.; Kremer, C.T.; Klausmeier, C.A.; Litchman, E. A global pattern of thermal adaptation in marine phytoplankton. Science 2012, 338, 1085–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobday, A.J.; Alexander, L.V.; Perkins, S.E.; Smale, D.A.; Straub, S.C.; Oliver, E.C.J.; Benthuysen, J.A.; Burrows, M.T.; Donat, M.G.; Feng, M.; et al. A hierarchical approach to defining marine heatwaves. Prog. Oceanogr. 2016, 141, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Oliver, E.C.J.; Donat, M.G.; Burrows, M.T.; Moore, P.J.; Smale, D.A.; Alexander, L.V.; Benthuysen, J.A.; Feng, M.; Sen Gupta, A.; Hobday, A.J.; et al. Longer and more frequent marine heatwaves over the past century. Nat. Commun. 2018, 9, 1324. [Google Scholar] [CrossRef] [PubMed]
- Wernberg, T.; Bennett, S.; Babcock, R.C.; De Bettignies, T.; Cure, K.; Depczynski, M.; Dufois, F.; Fromont, J.; Fulton, C.J.; Hovey, R.K.; et al. Climate-driven regime shift of a temperate marine ecosystem. Science 2016, 353, 169–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bond, N.A.; Cronin, M.F.; Freeland, H.; Mantua, N. Causes and impacts of the 2014 warm anomaly in the NE Pacific. Geophys. Res. Lett. 2015, 42, 3414–3420. [Google Scholar] [CrossRef]
- Irwin, A.J.; Finkel, Z.V.; Müller-Karger, F.E.; Ghinaglia, L.T. Phytoplankton adapt to changing ocean environments. Proc. Natl. Acad. Sci. USA 2015, 112, 5762–5766. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Koester, J.A.; Liefer, J.D.; Irwin, A.J.; Finkel, Z.V. Molecular mechanisms of temperature acclimation and adaptation in marine diatoms. ISME J. 2019, 13, 2415–2425. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Sun, J.; Guo, C.; Wei, Y.; Wang, X.; Yang, S.; Wu, C. Effects of typhoon Roke and Haitang on phytoplankton community structure in northeastern South China Sea. Ecosyst. Health Sustain. 2019, 5, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wu, C.; Xu, W.; Wang, X.; Thangaraj, S.; Zhang, G.; Zhang, X.; Zhao, Y.; Sun, J. Surface phytoplankton assemblages and controlling factors in the strait of Malacca and Sunda Shelf. Front. Mar. Sci. 2019, 7, 33. [Google Scholar] [CrossRef]
- Moore, C.M.; Mills, M.M.; Arrigo, K.R.; Berman-Frank, I.; Bopp, L.; Boyd, P.W.; Galbraith, E.D.; Geider, R.J.; Guieu, C.; Jaccard, S.L.; et al. Processes and patterns of oceanic nutrient limitation. Nat. Geosci. 2013, 6, 701–710. [Google Scholar] [CrossRef] [Green Version]






| Winter | Summer | ||||
|---|---|---|---|---|---|
| Species | f | Y | Species | f | Y |
| Thalassionema nitzschioides | 0.23 | 0.0231 | Trichodesmium thiebautii | 0.29 | 0.1842 |
| Nitzschia spp. | 0.33 | 0.0136 | Thalassionema nitzschioides | 0.57 | 0.0171 |
| Trichodesmium thiebautii | 0.05 | 0.0120 | Trichodesmium erythraeum | 0.13 | 0.0071 |
| Thalassiosira rotula | 0.30 | 0.0102 | Chaetoceros dichaeta | 0.28 | 0.0066 |
| Navicula spp. | 0.28 | 0.0089 | Chaetoceros affinis | 0.40 | 0.0054 |
| Chaetoceros spp. | 0.23 | 0.0081 | Thalassionema frauenfeldii | 0.54 | 0.0052 |
| Bacteriastrum spp. | 0.13 | 0.0066 | Chaetoceros lorenzianus | 0.30 | 0.0043 |
| Dictyocha fibula | 0.25 | 0.0057 | Pseudo-nitzschia delicatissima | 0.35 | 0.0041 |
| Thalassiosira subtilis | 0.20 | 0.0049 | Pseudo-nitzschia pungens | 0.28 | 0.0030 |
| Chaetoceros affinis | 0.08 | 0.0019 | Leptocylindrus danicus | 0.30 | 0.0026 |
| Chaetoceros coarctatus | 0.05 | 0.0016 | Hemiaulus hauckii | 0.37 | 0.0025 |
| Chaetoceros lorenzianus | 0.03 | 0.0014 | Nitzschia spp. | 0.65 | 0.0025 |
| Corethron hystrix | 0.05 | 0.0013 | Navicula spp. | 0.68 | 0.0022 |
| Chaetoceros atlanticus | 0.10 | 0.0008 | Bacteriastrum comosum | 0.23 | 0.0020 |
| Chaetoceros laciniosus | 0.05 | 0.0007 | Bacteriastrum hyalinum | 0.20 | 0.0019 |
| Thalassiothrix frauenfeldii | 0.05 | 0.0006 | Chaetoceros messanense | 0.23 | 0.0015 |
| Rizizosoleniu Rhizosolenia hebetata-Richelia | 0.08 | 0.0005 | Chaetoceros curvisetus | 0.14 | 0.0013 |
| Rhizosolenia hebetata | 0.03 | 0.0005 | Chaetoceros tortissimus | 0.16 | 0.0013 |
| Octactis octonaria | 0.08 | 0.0004 | Bacteriastrum elongatum | 0.21 | 0.0012 |
| Leptocylindrus mediterraneus | 0.03 | 0.0004 | Dactyliosolen blavyanus | 0.36 | 0.0010 |
| Sampling Date | Region | Water Depth (m) | Species Number | Abundance (103 cell L−1) | Reference |
|---|---|---|---|---|---|
| 1998–06–07 | 6–23° N,108–120° E | 0–150 m | 88 | 0.84 | [17] |
| 1998.06 | 5–25° N,105–120° E | Surface | 63 | 0.83 | [45] |
| 1998.08 | 18–22° N, 105–117° E | Surface | 58 | 181.00 | [45] |
| 1998.11–12 | 18–22° N, 105–117° E | 0–150 m | 85 | 8.46 | [17] |
| 2006.12 | 10–15° N, 110–112.5° E | 0–200 m | 117 | 2.74 | This study |
| 2007.08–09 | 10–15° N, 110–112.5° E | 0–200 m | 314 | 1.05 | This study |
| 2007.08 | 18–22° N, 110–120° E | 0–200 m | 216 | 11.22 | [46] |
| 2009.08 | 18–22° N, 110–117° E | 0–200 m | 109 | 8.20 | [47] |
| 2009.07–08 | 18–23.5° N, 109–120° E | 0–200 m | 150 | 26.49 | [48] |
| 2014.08 | 18–22° N, 114–116° E | 0–200 m | 229 | 16.32 | [14] |
| 2014.08 | 18–22° N, 114–116° E | Surface | 98 | 0.23 | [14] |
| 2015.07–08 | 21–23.5° N, 111–117° E | 0–200 m | 212 | 45.61 | [49] |
| 2017.07–08 | 14–23° N, 114–124° E | 0–200 m | 287 | 2.14 | [50] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, C.; Sun, J.; Narale, D.D.; Liu, H. Phytoplankton Community in the Western South China Sea in Winter and Summer. Water 2021, 13, 1209. https://doi.org/10.3390/w13091209
Ding C, Sun J, Narale DD, Liu H. Phytoplankton Community in the Western South China Sea in Winter and Summer. Water. 2021; 13(9):1209. https://doi.org/10.3390/w13091209
Chicago/Turabian StyleDing, Changling, Jun Sun, Dhiraj Dhondiram Narale, and Haijiao Liu. 2021. "Phytoplankton Community in the Western South China Sea in Winter and Summer" Water 13, no. 9: 1209. https://doi.org/10.3390/w13091209
APA StyleDing, C., Sun, J., Narale, D. D., & Liu, H. (2021). Phytoplankton Community in the Western South China Sea in Winter and Summer. Water, 13(9), 1209. https://doi.org/10.3390/w13091209