1. Introduction
The ecosystem in the water beneath our feet, i.e., in the groundwater, is organized in some aspect like all other aquatic systems [
1], e.g., gas solubility (dissolved oxygen, carbon dioxide, hydrogen sulfide) and the transport of carbon, nutrients (nitrogen, phosphorus etc.), and contaminants structure microbial (prokaryote, unicellular eukaryote = protozoa and fungi) and faunal (multicellular eukaryotes; metazoa) communities [
2]. Fauna and some microorganisms use dissolved oxygen [
3,
4] and produce dissolved organic carbon [
5,
6]. The sediment is also bioturbated by fauna [
7], thereby creating secondary porosity which is particularly important in porous aquifers. As the porosity and hydraulic pathways increase in complexity, the rates for bioremediation may increase, i.e., the biologically aided breakdown of contaminants [
8]. Thus, biological processes not only depend on the physical and chemical environment, but also influence it [
9]. This contribution is novel in that it summarizes in a condensed form the earlier elaborate developments that have arisen from studying the pore scale.
In contrast to most surface systems, two main factors constrain these processes in porous groundwater: the lack of light [
10] and the porosity [
11]. The lack of light excludes all phototrophic production, and groundwater systems rely on heterotrophy and chemolithoautotrophy. The porosity leads mostly to a reduction in the space available for growing and to decreased exchange [
11]. This exchange acts on all scales. The larger scale sets boundary conditions for the geochemical properties, the source of energy and nutrients, and exchange strength and processes. Groundwater aquifers are managed on the macro-scale (kilometers), and thus most understanding was gained at this scale. However, within these boundaries, the micrometer scale brings about variability, enabling a higher number of reduction and oxidation reactions. In a range of applications, such as the recovery of oil reservoirs or the storage of carbon dioxide, one needs to know more about processes in the pores [
12]. Bioremediation relies on such processes. In recent years, research into the ecological processes occurring at pore scale in groundwater is increasing, e.g., in order to understand bioremediation better, but knowledge is still scarce. One reason for this is that it is difficult to study unicellular organisms where they live and where they are predated upon. Sampling at the pore scale in situ remains a major challenge. Laboratory investigations provide insight but may present a distorted picture because of their artificial settings. The difficulties in studying the micro-scale in situ can be partly circumvented in silico, i.e., computer-aided, ecosystem modelling which considers the micro-scale [
13]. Such in silico models are usually based on conceptual models. The conceptual model introduced here as a method may become the basis for future groundwater ecological models.
2. Methods
The conceptual model was developed based on the recent knowledge introduced above. A conceptual model is a verbal or visual expression of general ideas. Tests are not easily possible, mostly because boundary conditions are usually not well-defined in a thought model, and usually the thought model is not quantitative. One example is the paradigm of groundwater fauna being generalist feeders and more (or less) sensitive than their surface relatives, or the assumption that fauna can only occur in the very surficial waters where exchange is high. This is a paradigm that clashes with observations of fauna in depauperate groundwater in up to 4 km depth [
14]. In contrast, quantitative, computational models define boundary conditions and assumptions not only as verbal or visual concepts, but by numbers and conditions that can be put into computer code. They range from univariate or multivariate statistical models that test hypotheses to spatially and/or temporally resolved models resulting in scenarios that cover a development of growth, contaminant degradation, clogging, etc. Conceptual and quantitative models may exist on all spatial and temporal scales but are more frequent at 0 dimensions (0D). Computational models mostly exist on the 1D scale (largely statistical models) or 2D (some process-based models). Modelling the third dimension adds complexity and is often considered too computationally demanding, considering the envisaged outcome.
Conceptual models may include state variables such as standing stocks, flows such as processes, and forcing functions such as boundary conditions, but conceptual models do not rely on such strict definitions. In contrast, computational models must define state variables, forcing functions and flows mathematically and quantitatively. Usually, a conceptual model is the first step for setting up a concrete computational model.
Here, the focus is on establishing a conceptual model of the groundwater ecosystem, which is the basis for the ongoing development of quantitative groundwater ecological models. First, the reasons for including several scales are explained, then the reasons for including microbial communities in models of groundwater fauna are given. Lastly, the reasons for including two size classes of fauna are explained.
On the macro-scale, i.e., landscape scale, major boundary conditions are set. For example, geological structures drive geochemical processes, and recharge from precipitation supplies the aquifer with energy and nutrients during the soil passage. On the meso-scale, i.e., meter scale, layers of material with different hydraulic conductivities limit or facilitate recharge into groundwater, thus driving exchange intensity and dynamics. On the microscale, i.e., in the pores, flow, diffusion, and the inhomogeneous distribution of organisms create a micro mosaic of redox situations, and thus a mosaic of reaction spaces and available resources that mix only on slow time scales. Modelling only on the macro-scale neglects the variety of processes taking place on the pore scale, for example, through microorganisms.
Macro-scale boundary conditions, such as recharge from the surface, cannot sufficiently explain the pore scale distribution of gases [
15], and thus they cannot predict the living conditions for microorganisms and for fauna feeding on the microorganisms. It is usually hypothesized that local oxygen minima in aquifers may be due to locally occurring, easily degradable organic matter. One major insight recently was that microbial processes on the micro-scale are critical for biogeochemical reactions [
13]. Depending on the resources available, microorganisms perform aerobic or anaerobic reactions. When they perform aerobic reactions, they may deplete dissolved oxygen levels. These depleted levels of dissolved oxygen exclude stygoxene (foreign to groundwater; usually surface water organisms) and stygophile (“friend to groundwater”; they use groundwater, but largely live on the surface, as opposed to the groundwater-bound stygobites) organisms, which become stressed at levels of oxygen that are still fine for stygobites [
16]. This easily degradable organic matter is used by microorganisms under the depletion of dissolved oxygen. Microorganisms occur in discrete colonies which usually only contain around 50 cells in the subsurface. Therefore, such depletion of oxygen and the subsequent biogeochemical reactions take place at the scale of these microcolonies. This, in turn, means that fauna not only has to find the sporadically occurring food sources, but also must cope with micro-scale patches of redox conditions and contaminants. The distribution and patterns of activity of fauna should therefore be conceptually and quantitatively modelled by considering the distribution and activity of their food source: the microorganisms.
Predators, even if only facultatively predatory, usually have a larger body size than their prey. Larger body sizes were found to coincide with relatively higher energy requirements [
17]. Where predators occur, production must be high enough to not only support microbial colonies, but also to support a surplus microbial production that feeds the predators indirectly through small faunal grazers. While fauna body size varies along a continuum, for a conceptual model, two fauna size classes and their respective feeding requirements appear to be appropriate for a first approximation. The smaller size class only grazes on microorganisms, while the larger size class uses the smaller size class fauna, at least facultatively.
The main points to include in any conceptual model of faunal distribution and activity are boundary conditions on different scales, including microbial communities, and the division of fauna into (at least) two size classes, where the larger size class feeds not only on microbial producers but also on the smaller faunal size class.
3. Results
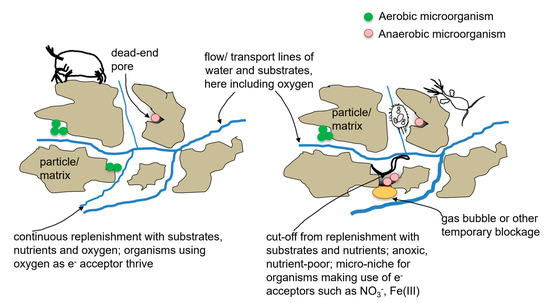
Figure 1 shows a conceptual model of the breakdown of micro-scale carbon-based substances. Such carbon-based substances, including pollutants, are found in the entire network of feeding relationships (
Figure 1). The
x axis represents time, and the
y axis represents space.
Figure 1 indicates how situations that differ in the way that physical laws apply or chemical reactions take place, and how water flows on the level of the micrometer pore scale (
Figure 1c,f), the millimeter pore scale (
Figure 1b,e), and on the level equivalent to tens of millimeters pore scale (
Figure 1a,d), may exist in close vicinity to each other. Each of the various situations provides an environment for a different biological community. Thus, the situations at each of the pore scales leads to the performance of different ecosystem functions.
The organisms depend on their environment, but they also shape their physical and chemical surroundings. In
Figure 1a–c (sequence from the largest to the smallest scale considered here) a well-replenished aquifer situation is shown, characterized by fast processes. Replenishment may result from fast recharge through highly conducive soil sediments and may include the introduction of dissolved oxygen, organic matter, and nutrients, but may also involve the intrusion of contaminants. In
Figure 1d–f (sequence from the largest to the smallest scale considered here) an impoverished situation is represented. The scarcity of resources in such a situation, also on the pore scale, may be due to low recharge from the surface, and geological and water chemical conditions that are low in dissolved oxygen, nutrients, and carbon. A scarcity of resources leads to fewer organisms finding enough resources. Here, only organisms that have a slow metabolism will survive, which is a typical trait for groundwater organisms [
18]. Since there are fewer organisms, fewer and slower transformations take place. In contrast, where the recharge introduces contaminants, the situation may also share similarities with a situation of lower resource recharge because the contaminants may be detrimental to the organisms. Depending on the type of contaminants, a high recharge situation looks like
Figure 1a–c, or leads to an impoverished food web with fewer stygoxene taxa in low numbers, as depicted in
Figure 1d–f.
The bubble in
Figure 1c, showing the micrometer pore scale, forms the fundament for communities of organisms that take on size quickly on the left side, i.e., supports what is depicted in
Figure 1b, which represents the millimeter pore scale, and 1a, covering tens of millimeters pore scale. These two sub plots represent consecutive parts of the food web with growing feeding pressure. The transformations of organic matter, sketched in
Figure 1c, yield the energy for microbial growth. In
Figure 1c, there are more and qualitatively better resources than in
Figure 1f which lead to the growth of more and larger microbial cells. Protozoa (
Figure 1b) graze on this increased microbial growth (
Figure 1c). Depicted in
Figure 1a, larger metazoa put feeding pressure on the small multicellular animals and unicellular organisms which are visible in
Figure 1b. These larger metazoa are usually crayfish.
Figure 1f shows lower concentrations of less easily degradable substrates compared to
Figure 1c. The degradation of the available substrates in an impoverished pore scale situation is presented in
Figure 1f, i.e., less degradable substrates yield less energy than the situation in
Figure 1c. The microbial colonies may grow to a lesser extent in
Figure 1f than in
Figure 1c. This means that fewer animals are able to retrieve sufficient food in
Figure 1d,e than in
Figure 1a,b. An increase in cell or body size is also slower in
Figure 1f, and thus in
Figure 1d,e, compared to
Figure 1c. The situation depicted in
Figure 1f supports only slow growth of a few small organisms in
Figure 1d,e. For exemplary situations represented by such scenarios, see [
11].
4. Conclusions and Outlook
Conceptual models, like the one presented here, are instruments not only for developing quantitative computational models on the pore scale, but also for knowledge exchange between disciplines. Since they are usually—and hopefully—more intuitive than presenting the full computational model, they may increase contact among various associations of interested people, including researchers, politicians and state agency employees. All these interest groups have a different way of talking to each other. They use specific words and terms. Scientists have begun to use such conceptual models and have furthered our understanding by modelling groundwater on the pore scale [
13,
19].
Following the paradigm of Occam’s razor [
20], to keep the model as simple as possible without losing too much information, only two size classes of fauna are considered necessary. This assumption, however, needs to be tested. There is also a lack of data exploring general microbial ecological patterns in groundwater. The available investigations show that microbial communities vary considerably according to the physical and chemical pore scale environment [
2]. It remains to be seen whether it is sufficient to subsume microbial production as one functional type, or whether models need to include the dominant functional types, which may depend on the aquifer itself.
Thus, the development of pore scale representations of the environment results in different wording of the topics we need to answer. This development highlights the areas in which we lack knowledge, such as:
Conceptual models for the impacts on all scales from groundwater fauna grazing;
Numbers for standing stocks of, and relationships among, prey and predators on the micrometer, millimeter, decimeter, meter, and kilometer extent;
Parameters, i.e., standing stocks;
Velocity of flow in the pore scale, velocity of predating, velocity of increase in cell or body size, velocity of substance breakdown, velocity of connection/losing touch, velocity of substance transformation for keeping body processes going, velocity of producing offspring, velocity of dying by unicellular and multicellular animals on the micrometer, millimeter, decimeter, meter, and kilometer extent;
Figures and numbers representing the burrowing activities in the subsurface living space and what they mean for the micrometer extent;
Translating processes on the pore scale, i.e., processes spanning micrometers and millimeters, to processes spanning meters and kilometers.
By closing these knowledge gaps, further insight into the suppositions and assumptions made by researchers from different disciplines may be gained and communicated. This is the basis for models that are better verified and validated.



{kind=link}
{kind=link}
