
Copepod Diel Vertical Distribution in the Open Southern Adriatic Sea (NE Mediterranean) under Two Different Environmental Conditions


Abstract
:1. Introduction
2. Materials and Methods
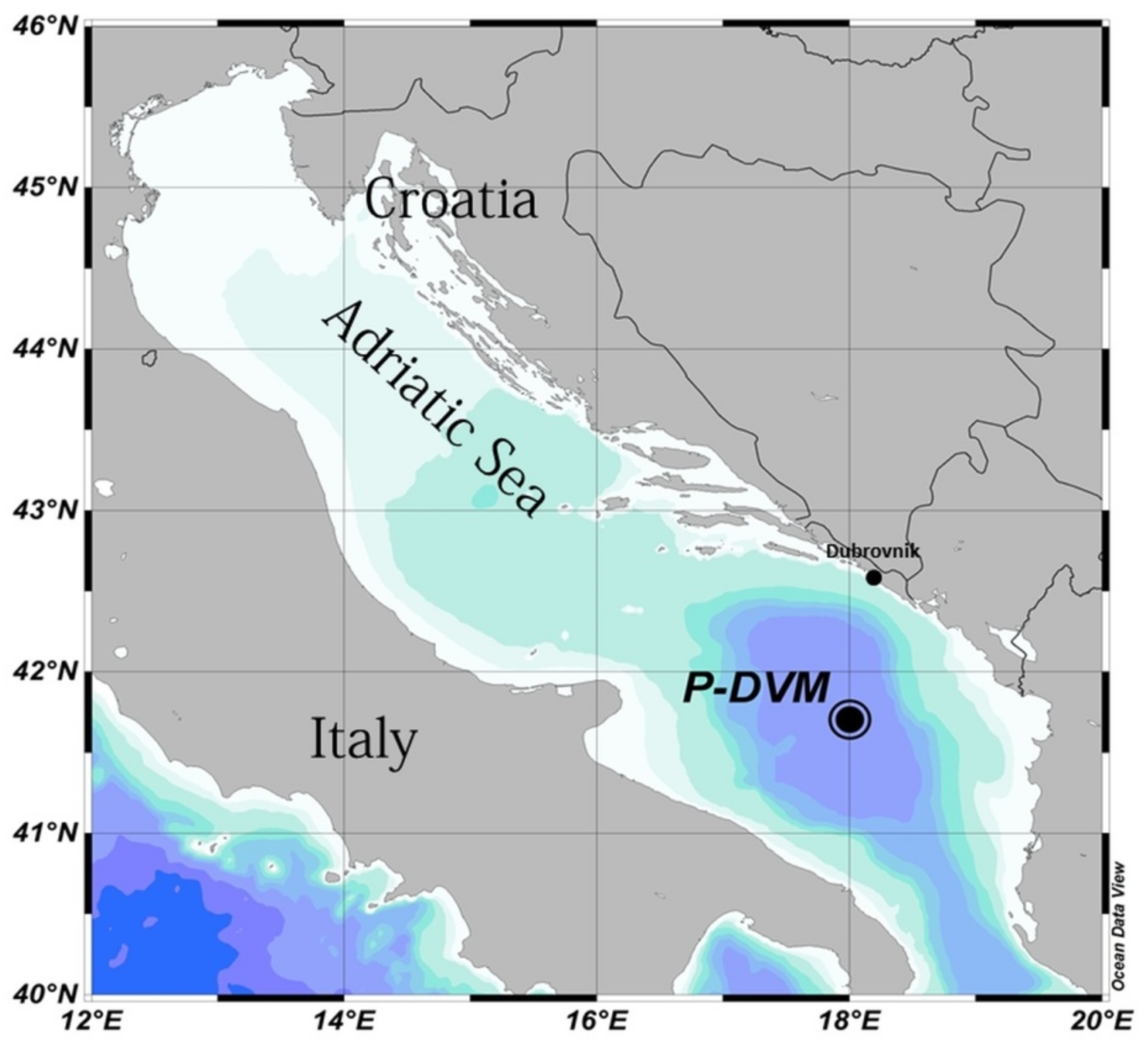
2.1. Study Area
2.2. Sampling and Laboratory Methods
2.3. Data Analysis
3. Results
3.1. Environmental Conditions
3.2. Vertical Distribution of Copepod Abundances and Diversity
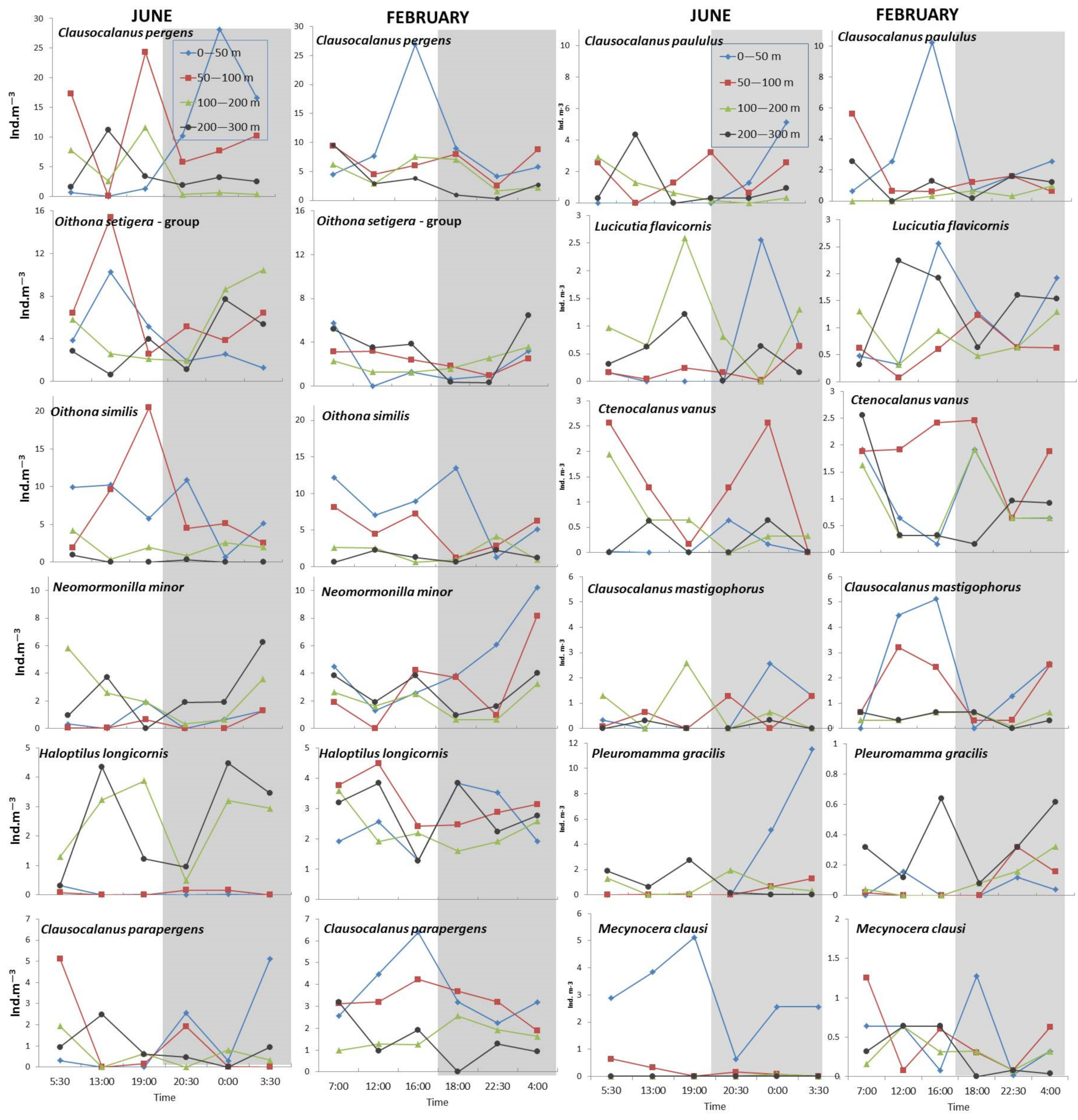
3.3. Dial and Seasonal Vertical Distribution Patterns of Dominant Copepods
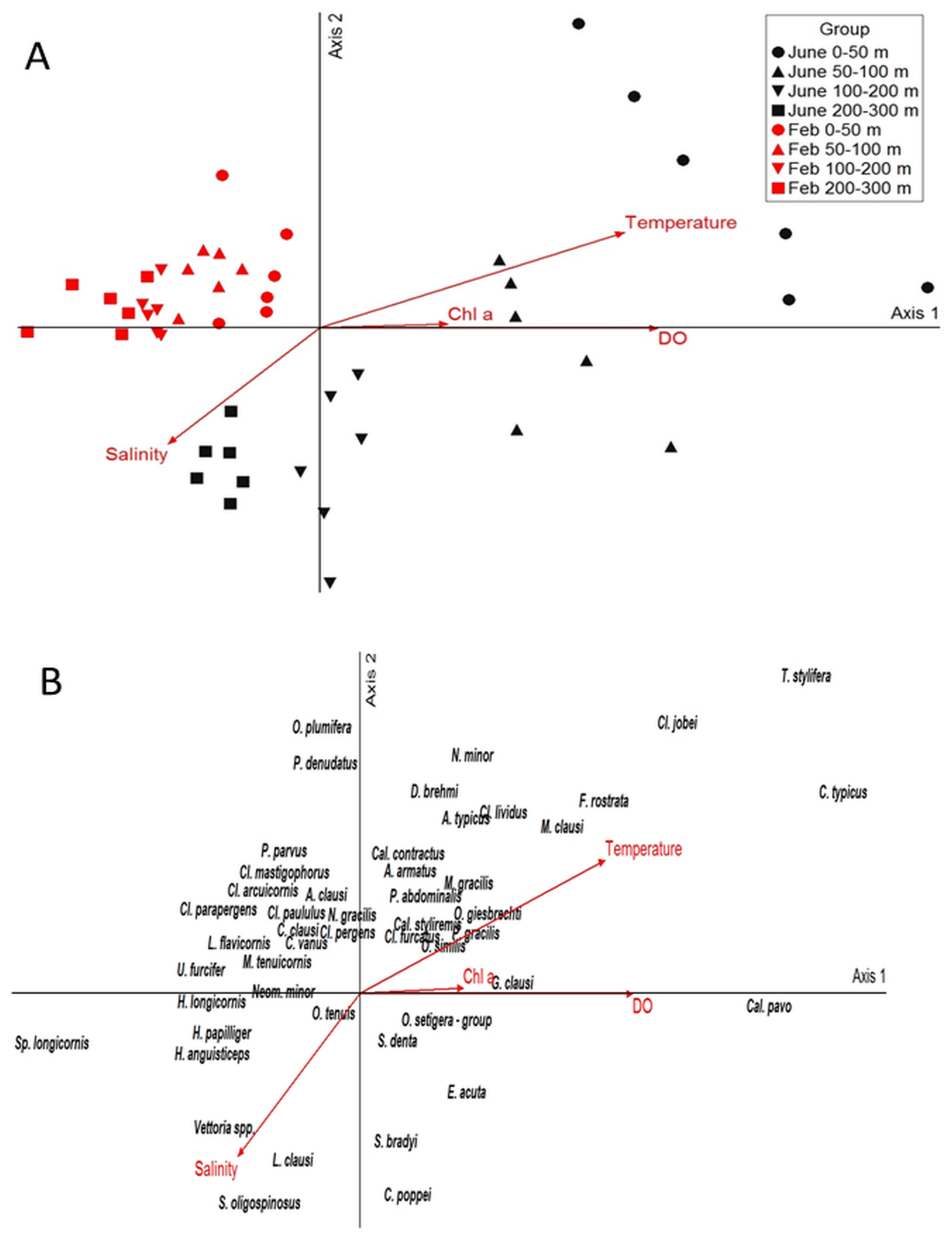
3.4. Environmental Drivers of the Copepod Community
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Licandro, P.; Icardi, P. Basin Scale Distribution of Zooplankton in the Ligurian Sea (North-Western Mediterranean) in Late Autumn. Hydrobiologia 2009, 617, 17–40. [Google Scholar] [CrossRef]
- Hays, G.; Harris, R.; Head, R. The Vertical Nitrogen Flux Caused by Zooplankton Diel Vertical Migration. Mar. Ecol. Prog. Ser. 1997, 160, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Schnetzer, A.; Steinberg, D. Active Transport of Particulate Organic Carbon and Nitrogen by Vertically Migrating Zooplankton in the Sargasso Sea. Mar. Ecol. Prog. Ser. 2002, 234, 71–84. [Google Scholar] [CrossRef]
- Hays, G.C. A Review of the Adaptive Significance and Ecosystem Consequences of Zooplankton Diel Vertical Migrations. In Migrations and Dispersal of Marine Organisms; Jones, M.B., Ingólfsson, A., Ólafsson, E., Helgason, G.V., Gunnarsson, K., Svavarsson, J., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2003; pp. 163–170. [Google Scholar] [CrossRef]
- Dawidowicz, P.; Pijanowska, J. Diel Vertical Migration of Aquatic Crustaceans: Adaptive Role, Underlying Mechanisms, and Ecosystem Consequences. In The Natural History of the Crustacea: Life Histories; Oxford University Press: Oxford, UK, 2018; pp. 231–256. [Google Scholar]
- Cushing, D.H. The vertical migration of planktonic crustacea. Biol. Rev. 1951, 26, 158–192. [Google Scholar] [CrossRef] [PubMed]
- Winfried, L. Ultimate Causes of Diel Vertical Migration of Zooplankton: New Evidence for the Predator-Avoidance Hypothesis. Arch. Hydrobiol. Beih. Ergeb. Limnol. 1993, 39, 79–88. [Google Scholar]
- Dale, T. Diel Patterns in Stage-Specific Vertical Migration of Calanus Finmarchicus in Habitats with Midnight Sun. ICES J. Mar. Sci. 2000, 57, 1800–1818. [Google Scholar] [CrossRef]
- Kaartvedt, S. Habitat Preference during Overwintering and Timing of Seasonal Vertical Migration of Calanus finmarchicus. Ophelia 1996, 44, 145–156. [Google Scholar] [CrossRef]
- De Robertis, A. Size-Dependent Visual Predation Risk and the Timing of Vertical Migration: An Optimization Model. Limnol. Oceanogr. 2002, 47, 925–933. [Google Scholar] [CrossRef]
- Liu, S.-H. Diel Vertical Migration of Zooplankton Following Optimal Food Intake under Predation. J. Plankton Res. 2003, 25, 1069–1077. [Google Scholar] [CrossRef] [Green Version]
- Isla, A.; Scharek, R.; Latasa, M. Zooplankton Diel Vertical Migration and Contribution to Deep Active Carbon Flux in the NW Mediterranean. J. Mar. Syst. 2015, 143, 86–97. [Google Scholar] [CrossRef]
- Hure, J.; Kršinić, F. Planktonic Copepods of the Adriatic Sea. Nat. Croat. 1998, 7, 1–135. [Google Scholar]
- Hure, J. Distribution Annuelle Vertical Du Zooplankton Sur Une Station de l’Adriatique Méridionale. Acta Adriat. 1955, 7, 1–72. [Google Scholar]
- Hure, J. Dnevna Migracija i Sezonska Vertikalna Raspodjela Zooplanktona Dubljeg Mora. Acta Adriat. 1961, 9, 1–59. [Google Scholar]
- Hure, J.; Scotto di Carlo, B. Diurnal Vertical Migration of Some Deep Water Copepods in the Southern Adriatic (East Mediterranean). Pubbl. Staz. Zool. Napoli 1969, 37, 581–598. [Google Scholar]
- Hure, J.; Scotto di Carlo, B. Ripartizione Quantitative e Distribuzione Verticale Dei Copepodi Pelagici Di Profondita Su Una Stazione Nel Mar Tirreno Ed Una Nell’Adriatico Meridionale. Pubbl. Staz. Zool. Napoli 1969, 37, 51–83. [Google Scholar]
- Batistić, M.; Jasprica, N.; Carić, M.; Čalić, M.; Kovačević, V.; Garić, R.; Njire, J.; Mikuš, J.; Bobanović-Ćolić, S. Biological Evidence of a Winter Convection Event in the South Adriatic: A Phytoplankton Maximum in the Aphotic Zone. Cont. Shelf Res. 2012, 44, 57–71. [Google Scholar] [CrossRef]
- Hure, M.; Mihanović, H.; Lučić, D.; Ljubešić, Z.; Kružić, P. Mesozooplankton Spatial Distribution and Community Structure in the South Adriatic Sea during Two Winters (2015, 2016). Mar. Ecol. 2018, 39, e12488. [Google Scholar] [CrossRef]
- Hure, M.; Batistić, M.; Kovačević, V.; Bensi, M.; Garić, R. Copepod Community Structure in Pre- and Post- Winter Conditions in the Southern Adriatic Sea (NE Mediterranean). J. Mar. Sci. Eng. 2020, 8, 567. [Google Scholar] [CrossRef]
- Ursella, L.; Cardin, V.; Batistić, M.; Garić, R.; Gačić, M. Evidence of Zooplankton Vertical Migration from Continuous Southern Adriatic Buoy Current-Meter Records. Progr. Oceanogr. 2018, 167, 78–96. [Google Scholar] [CrossRef]
- Gačić, M.; Civitarese, G.; Miserocchi, S.; Cardin, V.; Crise, A.; Mauri, E. The Open-Ocean Convection in the Southern Adriatic: A Controlling Mechanism of the Spring Phytoplankton Bloom. Cont. Shelf Res. 2002, 22, 1897–1908. [Google Scholar] [CrossRef]
- Batistić, M.; Viličić, D.; Kovačević, V.; Jasprica, N.; Garić, R.; Lavigne, H.; Carić, M. Occurrence of Winter Phytoplankton Bloom in the Open Southern Adriatic: Relationship with Hydroclimatic Events in the Eastern Mediterranean. Cont. Shelf Res. 2019, 174, 12–25. [Google Scholar] [CrossRef]
- Ljubimir, S.; Jasprica, N.; Čalić, M.; Hrustić, E.; Dupčić Radić, I.; Car, A.; Batistić, M. Interannual (2009–2013) Variability of Winter-Spring Phytoplankton in the Open South Adriatic Sea: Effects of Deep Convection and Lateral Advection. Cont. Shelf Res. 2017, 143, 311–321. [Google Scholar] [CrossRef]
- Gačić, M.; Marullo, S.; Santoleri, R.; Bergamasco, A. Analysis of the Seasonal and Interannual Variability of the Sea Surface Temperature Field in the Adriatic Sea from AVHRR Data (1984-1992). J. Geophys. Res. 1997, 102, 22937–22946. [Google Scholar] [CrossRef]
- Faganeli, J.; Gačić, M.; Malej, A.; Smodlaka, N. Pelagic Organic Matter in the Adriatic Sea in Relation to Winter Hydrographic Conditions. J. Plankton Res. 1989, 11, 1129–1141. [Google Scholar] [CrossRef]
- Malanotte-Rizzoli, P.; Robinson, A.R.; Roether, W.; Manca, B.; Bergamasco, A.; Brenner, S.; Civitarese, G.; Georgopoulos, D.; Haley, P.J.; Kioroglou, S.; et al. Experiment in Eastern Mediterranean Probes Origin of Deep Water Masses. Eos Trans. AGU 1996, 77, 305. [Google Scholar] [CrossRef]
- Gačić, M.; Borzelli, G.L.E.; Civitarese, G.; Cardin, V.; Yari, S. Can Internal Processes Sustain Reversals of the Ocean Upper Circulation? The Ionian Sea Example: Internal processes and upper circulation. Geophys. Res. Lett. 2010, 37, L09608. [Google Scholar] [CrossRef]
- Vilibić, I.; Matijević, S.; Šepić, J.; Kušpilić, G. Changes in the Adriatic Oceanographic Properties Induced by the Eastern Mediterranean Transient. Biogeosciences 2012, 9, 2085–2097. [Google Scholar] [CrossRef] [Green Version]
- Batistić, M.; Garić, R.; Molinero, J. Interannual Variations in Adriatic Sea Zooplankton Mirror Shifts in Circulation Regimes in the Ionian Sea. Clim. Res. 2014, 61, 231–240. [Google Scholar] [CrossRef]
- Lučić, D.; Ljubešić, Z.; Babić, I.; Bosak, S.; Cetinić, I.; Vilibić, I.; Mihanović, H.; Hure, M.; Njire, J.; Lučić, P.; et al. Unusual Winter Zooplankton Bloom in the Open Southern Adriatic Sea. Turk. J. Zool. 2017, 471, 1024–1035. [Google Scholar] [CrossRef]
- Čalić, M.; Ljubimir, S.; Bosak, S.; Car, A. First Records of Two Planktonic Indo-Pacific Diatoms: Chaetoceros Bacteriastroides and C. pseudosymmetricus in the Adriatic Sea. Oceanologia 2018, 60, 101–105. [Google Scholar] [CrossRef]
- Njire, J.; Batistić, M.; Kovačević, V.; Garić, R.; Bensi, M. Tintinnid Ciliate Communities in Pre- and Post-Winter Conditions in the Southern Adriatic Sea (NE Mediterranean). Water 2019, 11, 2329. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E.; Wiener, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1963. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. Primer V5: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2001. [Google Scholar]
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- McCune, B.; Mefford, M.J. PC-ORD Multivariate Analysis of Ecological Data; MjM Software: Gleneden Beach, OR, USA, 2006. [Google Scholar]
- Weikert, H.; Trinkaus, S. Vertical Mesozooplankton Abundance and Distribution in the Deep Eastern Mediterranean Sea SE of Crete. J. Plankton Res. 1990, 12, 601–628. [Google Scholar] [CrossRef]
- Siokou-Frangou, I. Zooplankton Annual Cycle in a Mediterranean Coastal Area. J. Plankton Res. 1996, 18, 203–223. [Google Scholar] [CrossRef] [Green Version]
- Mazzocchi, M.G. Spring Mesozooplankton Communities in the Epipelagic Ionian Sea in Relation to the Eastern Mediterranean Transient. J. Geophys. Res. 2003, 108, 8114. [Google Scholar] [CrossRef] [Green Version]
- Bottger-Schnack, R. Vertical Structure of Small Metazoan Plankton, Especially Non-Calanoid Copepods. I. Deep Arabian Sea. J. Plankton Res. 1996, 18, 1073–1101. [Google Scholar] [CrossRef]
- Calbet, A. Annual Zooplankton Succession in Coastal NW Mediterranean Waters: The Importance of the Smaller Size Fractions. J. Plankton Res. 2001, 23, 319–331. [Google Scholar] [CrossRef]
- Pansera, M.; Granata, A.; Guglielmo, L.; Minutoli, R.; Zagami, G.; Brugnano, C. How Does Mesh-Size Selection Reshape the Description of Zooplankton Community Structure in Coastal Lakes? Estuar. Coast. Shelf Sci. 2014, 151, 221–235. [Google Scholar] [CrossRef]
- Scotto di Carlo, B.; Ianora, A.; Fresi, E.; Hure, J. Vertical Zonation Patterns for Mediterranean Copepods from the Surface to 3000 m at a Fixed Station in the Tyrrhenian Sea. J. Plankton Res. 1984, 6, 1031–1056. [Google Scholar] [CrossRef]
- Mazzocchi, M.G.; Christou, E.D.; Fragopoulu, N.; Siokou-Frangou, I. Mesozooplankton Distribution from Sicily to Cyprus (Eastern Mediterranean): I. General Aspects. Oceanol. Acta 1997, 20, 521–535. [Google Scholar]
- Siokou-Frangou, I.; Christou, E.D.; Fragopoulu, N.; Mazzocchi, M.G. Mesozooplankton Distribution from Sicily to Cyprus (Eastern Mediterranean): II. Copepod Assemblages. Oceanol. Acta 1997, 20, 537–548. [Google Scholar]
- Brugnano, C.; Granata, A.; Guglielmo, L.; Zagami, G. Spring Diel Vertical Distribution of Copepod Abundances and Diversity in the Open Central Tyrrhenian Sea (Western Mediterranean). J. Mar. Syst. 2012, 105–108, 207–220. [Google Scholar] [CrossRef]
- Zagami, G.; Granata, A.; Brugnano, C.; Minutoli, R.; Bonanzinga, V.; Guglielmo, L. Spring Copepod Vertical Zonation Pattern and Diel Migration in the Open Ligurian Sea (North-Western Mediterranean). Progr. Oceanogr. 2020, 183, 102297. [Google Scholar] [CrossRef]
- Kiorboe, T.; Nielsen, T.G. Regulation of Zooplankton Biomass and Production in a Temperate, Coastal Ecosystem. 1. Copepods. Limnol. Oceanogr. 1994, 39, 493–507. [Google Scholar] [CrossRef]
- Turner, J.T. The Importance of Small Planktonic Copepods and Their Roles in Pelagic Marine Food. Zool. Stud. 2004, 43, 255–266. [Google Scholar]
- Nakamura, Y.; Turner, J.T. Predation and Respiration by the Small Cyclopoid Copepod Oithona similisr: How Important Is Feeding on Ciliates and Heterotrophic Flagellates? J. Plankton Res. 1997, 19, 1275–1288. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.A.; Muxagata, E. The Seasonal Abundance and Production of Oithona Nana (Copepoda:Cyclopoida) in Southampton Water. J. Plankton Res. 2006, 28, 1055–1065. [Google Scholar] [CrossRef] [Green Version]
- Miloslavić, M.; Lučić, D.; Žarić, M.; Gangai, B.; Onofri, I. The Importance of Vertical Habitat Gradients on Zooplankton Distribution in an Enclosed Marine Environment (South Adriatic Sea). Mar. Biol. Res. 2015, 11, 462–474. [Google Scholar] [CrossRef]
- Irigoien, X.; Conway, D.; Harris, R. Flexible Diel Vertical Migration Behaviour of Zooplankton in the Irish Sea. Mar. Ecol. Prog. Ser. 2004, 267, 85–97. [Google Scholar] [CrossRef] [Green Version]
- Hure, J.; Ianora, A.; Scotto di Carlo, B. Spatial and Temporal Distribution of Copepod Communities in the Adriatic Sea. J. Plankton Res. 1980, 2, 295–316. [Google Scholar] [CrossRef]
- Andersen, V. Zooplankton Community During the Transition from Spring Bloom to Oligotrophy in the Open NW Mediterranean and Effects of Wind Events. 1. Abundance and Specific Composition. J. Plankton Res. 2001, 23, 227–242. [Google Scholar] [CrossRef] [Green Version]
- Siokou, I.; Zervoudaki, S.; Christou, E.D. Mesozooplankton Community Distribution down to 1000 m along a Gradient of Oligotrophy in the Eastern Mediterranean Sea (Aegean Sea). J. Plankton Res. 2013, 35, 1313–1330. [Google Scholar] [CrossRef] [Green Version]
- Koppelmann, R.; Weikert, H. Spatial and Temporal Distribution Patterns of Deep-Sea Mesozooplankton in the Eastern Mediterranean? Indications of a Climatically Induced Shift? Mar. Ecol. 2007, 28, 259–275. [Google Scholar] [CrossRef]
- Kehayias, G.; Fragopoulu, N.; Lykakis, J. Vertical Community Structure and Ontogenetic Distribution of Chaetognaths in Upper Pelagic Waters of the Eastern Mediterranean. Mar. Biol. 1994, 119, 647–653. [Google Scholar] [CrossRef]
- Kouwenberg, J.H.M. Copepod Distribution in Relation to Seasonal Hydrographics and Spatial Structure in the North-Western Mediterranean (Golfe Du Lion). Estuar. Coast. Shelf Sci. 1994, 38, 69–90. [Google Scholar] [CrossRef] [Green Version]
- Peralba, À.; Mazzocchi, M.G. Vertical and Seasonal Distribution of Eight Clausocalanus Species (Copepoda: Calanoida) in Oligotrophic Waters. ICES J. Mar. Sci. 2004, 61, 645–653. [Google Scholar] [CrossRef]
- Kleppel, G.S.; Frazel, D.; Pieper, R.E. Natural Diets of Zooplankton off Southern California. Mar. Ecol. Prog. 1988, 49, 231–241. [Google Scholar] [CrossRef]
- Mazzocchi, M.G.; Paffenhöfer, G.-A. First Observations on the Biology of Clausocalanus Furcatus (Copepoda, Calanoida). J. Plankton Res. 1998, 20, 331–342. [Google Scholar] [CrossRef] [Green Version]
- Cummings, J.A. Habitat Dimensions of Calanoid Copepods in the Western Gulf of Mexico. J. Mar. Res. 1984, 42, 163–188. [Google Scholar] [CrossRef]
- Frederiksen, M.; Edwards, M.; Richardson, A.J.; Halliday, N.C.; Wanless, S. From Plankton to Top Predators: Bottom-up Control of a Marine Food Web across Four Trophic Levels. J. Anim. Ecol. 2006, 75, 1259–1268. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| June | February | |||||
|---|---|---|---|---|---|---|
| Abundance | WMD | Abundance | WMD | t | p | |
| Clausocalanus pergens | 7.0 ± 7.9 | 116 | 6.0 ± 5.3 | 118 | 0.17 | 0.86 |
| Oithona setigera—group | 4.9 ± 3.6 | 140 | 2.4 ± 1.7 | 166 | −0.17 | 0.86 |
| Oithona similis | 4.2 ± 4.6 | 76 | 4.1 ± 4.1 | 94 | −1.03 | 0.31 |
| Neonmormonilla minor | 1.5 ± 1.8 | 185 | 3.1 ± 2.4 | 133 | 0.26 | 0.79 |
| Haloptilus longicornis | 1.3 ± 1.6 | 198 | 2.7 ± 0.9 | 151 | 0.93 | 0.36 |
| Clausocalanus parapergens | 1.0 ± 1.5 | 136 | 2.5 ± 1.4 | 117 | 0.64 | 0.52 |
| Clausocalanus paululus | 1.2 ± 1.5 | 142 | 1.6 ± 2.2 | 114 | 0.90 | 0.36 |
| Lucicutia flavicornis | 0.6 ± 0.7 | 153 | 1 ± 0.7 | 162 | 0.53 | 0.60 |
| Ctenocalanus vanus | 0.6 ± 0.8 | 127 | 1.2 ± 0.8 | 136 | −0.11 | 0.90 |
| Clausocalanus mastigophorus | 0.5 ± 0.8 | 114 | 1.2 ± 1.4 | 94 | −0.61 | 0.54 |
| Pleuromamma gracilis | 1.2 ± 2.5 | 120 | 0.2 ± 1.2 | 204 | 0.25 | 0.80 |
| Mecynocera clausi | 0.8 ± 1.4 | 29 | 0.4 ± 0.4 | 131 | −0.24 | 0.02 |
| Clausocalanus cop. spp. | 5.9 ± 0.8 | 106 | 6.4 ± 6.2 | 111 | −0.13 | 0.89 |
| Pleuromamma cop. spp. | 0.7 ± 1.6 | 107 | 0.4 ± 0.3 | 167 | 0.83 | 0.41 |
| Enviromnental Variables/Axes | Axis 1 | Axis 2 |
|---|---|---|
| Temperature | 0.816 | 0.514 |
| Salinity | −0.570 | −0.541 |
| DO | 0.878 | 0.043 |
| Chl a | 0.534 | 0.135 |
| Group of Sampling Stations Tested for the Following Hypotheses | T | A | p-Value |
|---|---|---|---|
| June (24) versus February (24) | −16.486 | 0.2047 | 0.0000 |
| Depth strata (0–50 m (12), 50–100 m (12), 100–200 m(12), 200–300 m (12)) | −0.399 | 0.0909 | 0.0016 |
| Day (24) versus night (24) | −2.329 | 0.0295 | 0.0324 |
| Null Hypotheseses Tested for no Abundance Differences among Station Groups | Copepod Taxa | Indicator Value | Group | p-Value |
|---|---|---|---|---|
| June (24) versus February (24) | Oncaeidea | 65.8 | June | 0.0030 |
| Oithona setigera-group | 59.4 | June | 0.0030 | |
| Euchaeta acuta | 51.2 | June | 0.0010 | |
| Lucicutia clausi | 40.7 | June | 0.0240 | |
| Calocalanus pavo | 37.5 | June | 0.0020 | |
| Chiridium poppei | 28.6 | June | 0.0334 | |
| Spinocalanus oligospinosus | 25.0 | June | 0.0222 | |
| Temora stylifera | 24.4 | June | 0.0464 | |
| Paracalanus parvus | 84.4 | February | 0.0002 | |
| Urocorycaeus furcifer | 74.3 | February | 0.0004 | |
| Haloptilus longicornis | 68.2 | February | 0.0002 | |
| Clausocalanus parapergens | 67.5 | February | 0.0002 | |
| Ctenocalanus vanus | 66.1 | February | 0.0008 | |
| Lucicutia flavicornis | 63.2 | February | 0.0056 | |
| Neomormonilla minor | 61.4 | February | 0.0054 | |
| Clausocalanus mastigophorus | 57.3 | February | 0.0090 | |
| Corycaeus clausi | 47.3 | February | 0.0170 | |
| Onychocorycaeus giesbrechty | 46.3 | February | 0.0050 | |
| Heterorhabdus papilliger | 45.9 | February | 0.0204 | |
| Paracalanus denudatus | 30.3 | February | 0.0492 | |
| Depth strata (0–50 m (12), 50–100 m (12), 100–200 m (12), 200–300 m (12)) | Centropages typicus | 71.4 | 0–50 m | 0.0002 |
| Mecynocera clausi | 61.6 | 0–50 m | 0.0002 | |
| Farranula rostrata | 61.4 | 0–50 m | 0.0002 | |
| Temora stylifera | 48.8 | 0–50 m | 0.0010 | |
| Onychocorycaeus giesbrechty | 48.3 | 0–50 m | 0.0018 | |
| Oithona similis | 37.7 | 0–50 m | 0.0018 | |
| Acartia (Acartiura) clausi | 35.3 | 0–50 m | 0.0356 | |
| Paracalanus denudatus | 30.8 | 0–50 m | 0.0476 | |
| Calocalanus styliremis | 43.6 | 50–100 m | 0.0028 | |
| Ctenocalanus vanus | 37.8 | 50–100 m | 0.0134 | |
| Heterorhabdus papilliger | 38.2 | 100–200 m | 0.0222 | |
| Haloptilus anguiceps | 38.5 | 200–300 m | 0.0392 | |
| Lucicutia clausi | 38.0 | 200–300 m | 0.0170 | |
| Spinocalanus oligospinosus | 29.8 | 200–300 m | 0.0250 | |
| Day (24) versus night (24) | Pleuromamma gracilis | 55.1 | Night | 0.0488 |
| Scolecithricella dentata | 53.2 | Night | 0.0210 | |
| Pleuromamma abdominalis | 48.3 | Night | 0.0086 | |
| Euchaeta acuta | 40.6 | Night | 0.0394 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hure, M.; Batistić, M.; Garić, R. Copepod Diel Vertical Distribution in the Open Southern Adriatic Sea (NE Mediterranean) under Two Different Environmental Conditions. Water 2022, 14, 1901. https://doi.org/10.3390/w14121901
Hure M, Batistić M, Garić R. Copepod Diel Vertical Distribution in the Open Southern Adriatic Sea (NE Mediterranean) under Two Different Environmental Conditions. Water. 2022; 14(12):1901. https://doi.org/10.3390/w14121901
Chicago/Turabian StyleHure, Marijana, Mirna Batistić, and Rade Garić. 2022. "Copepod Diel Vertical Distribution in the Open Southern Adriatic Sea (NE Mediterranean) under Two Different Environmental Conditions" Water 14, no. 12: 1901. https://doi.org/10.3390/w14121901
APA StyleHure, M., Batistić, M., & Garić, R. (2022). Copepod Diel Vertical Distribution in the Open Southern Adriatic Sea (NE Mediterranean) under Two Different Environmental Conditions. Water, 14(12), 1901. https://doi.org/10.3390/w14121901